| Original Literature | Model OverView |
|---|---|
|
Publication
Title
Functions of anaphylatoxin C5a in rat liver: direct and indirect actions onnonparenchymal and parenchymal cells.
Affiliation
Institut fur Biochemie und Molekulare Zellbiologie, Georg-August-UniversitatGottingen, Humboldtallee 23, D-37073 Gottingen, Germany. hschief@gwdg.de
Abstract
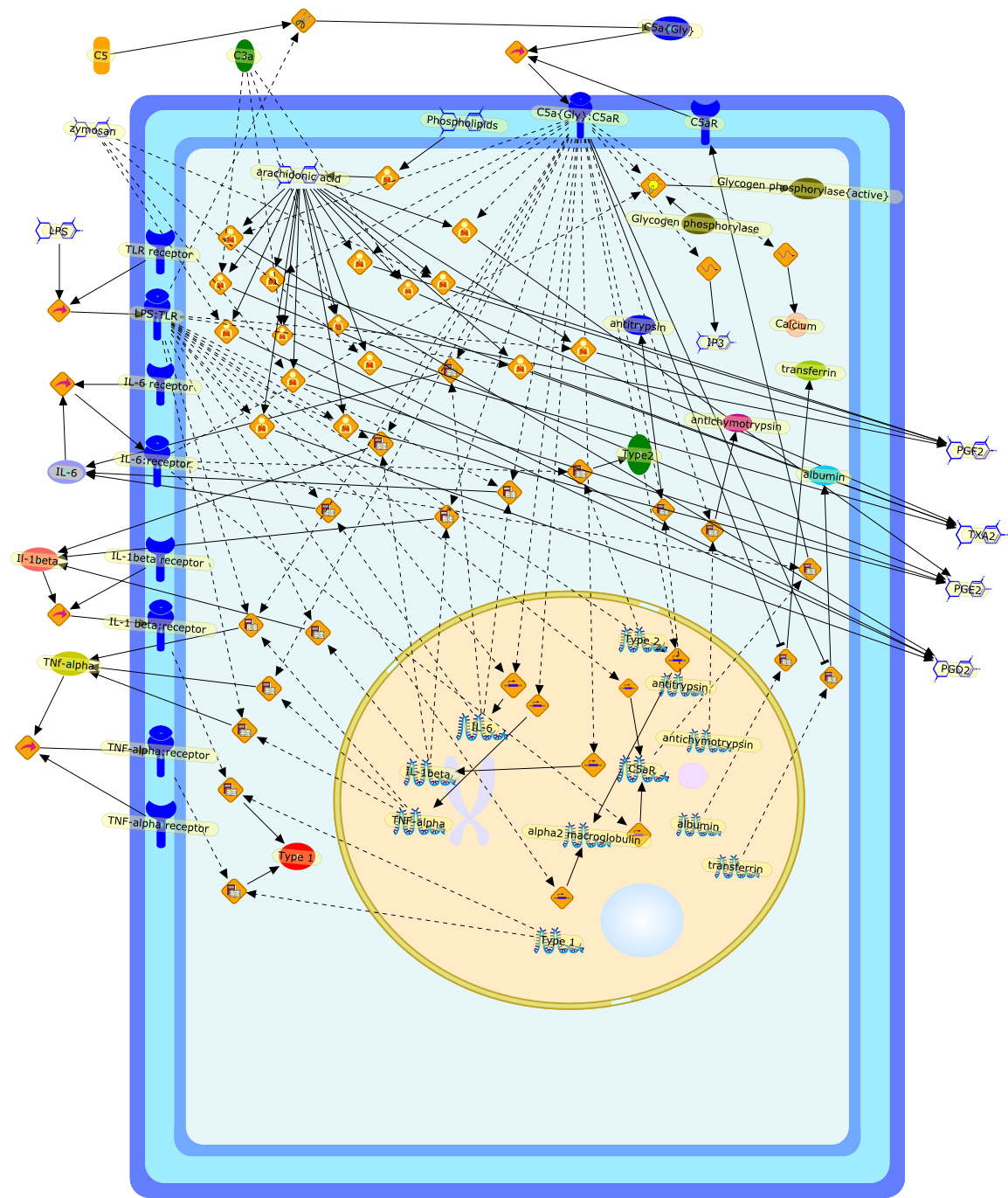
Growing evidence obtained in recent years indicates that anaphylatoxin C5areceptors (C5aR) are not restricted to myeloid cells but are also expressed onnonmyeloid cells in different tissues such as brain, lung, skin and liver. Incontrast to its well-defined systemic functions, the actions of anaphylatoxinsin these organs are poorly characterized. The liver can be a primary targetorgan for the C5a anaphylatoxin since the liver is directly connected to thegut, via the mesenteric veins and portal vein which is a main source ofcomplement activating lipopolysaccharides (LPS). In the normal rat liver, theC5aR is only expressed by nonparenchymal cells, i.e. strongly by Kupffer cells(KC) and hepatic stellate cells (HSC) and weakly by sinusoidal endothelial cells(SEC), but not expressed by the parenchymal hepatocytes (HC). Accordingly,direct effects of C5a were only found in the C5aR-expressing KC and HSC: C5ainduced the release of prostanoids from KC and HSC and enhanced theLPS-dependent release of interleukin-6 from KC. These soluble mediatorsindirectly influenced effector functions of the C5aR-free HC. C5a enhanced theglycogen phosphorylase activity and thus the glucose output from HC indirectlyvia prostanoids released from KC and HSC. Glucose can serve as an energysubstrate as well as an electron donor for the synthesis of reactive oxygenintermediates by KC. Moreover, C5a also enhanced transcription of the gene forthe type-2 acute phase protein alpha 2-macroglobulin in HC indirectly byincreasing LPS-dependent IL-6 release from KC. Under pathological conditions,C5aR was found to be upregulated in various organs including the liver.Simulation of inflammatory conditions by treatment of rats with IL-6, a maininflammatory mediator in the liver, caused a de novo expression of functionalC5aR in HC. In livers of IL-6-treated rats, C5a initiated glucose output from HCand perhaps other HC-specific defense reactions directly without theintervention of soluble mediators from nonparenchymal cells.
PMID
11367531
|
 |