| Original Literature | Model OverView |
|---|---|
|
Publication
Title
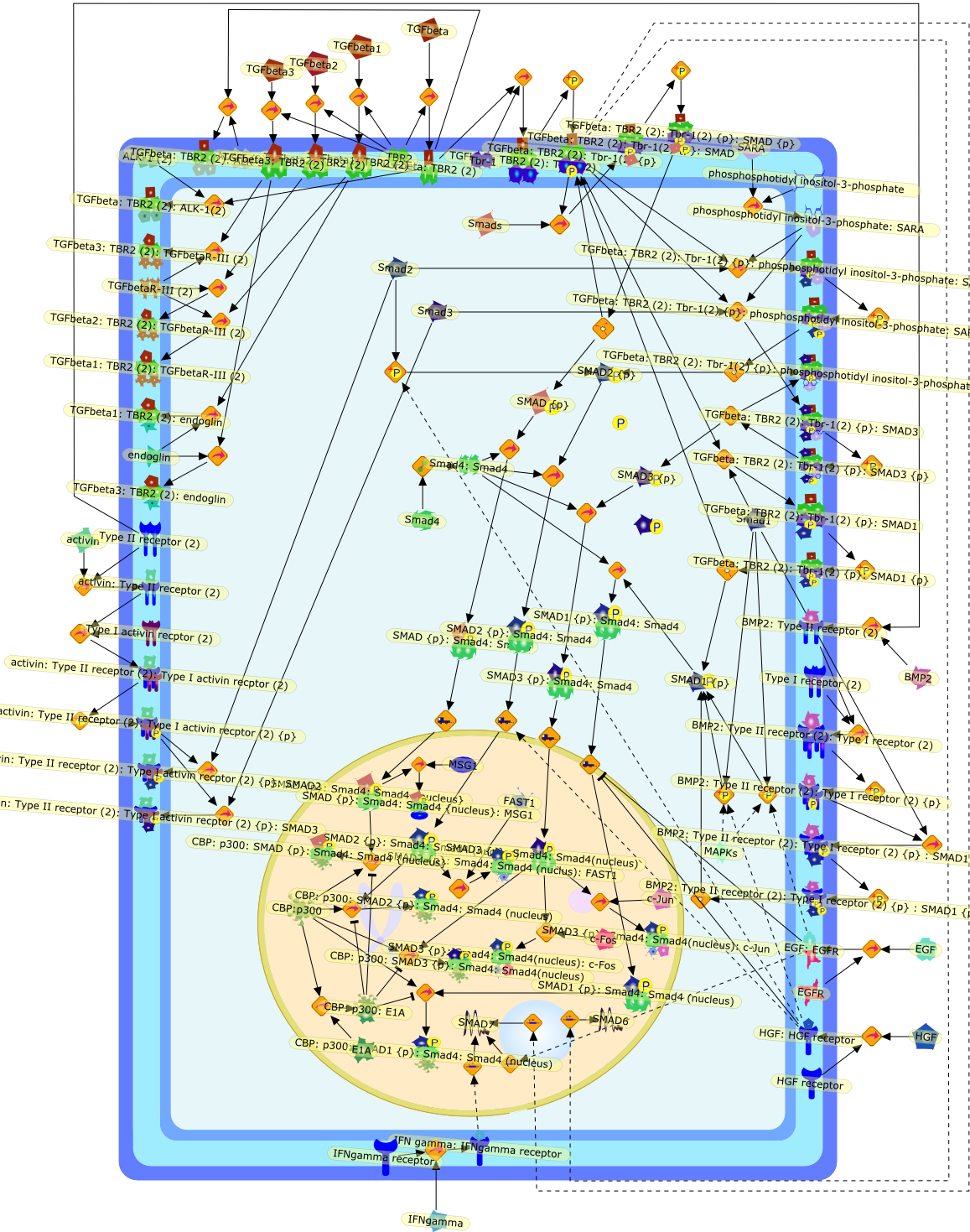
TGF-beta signaling from receptors to the nucleus.
Affiliation
Laboratory of Cell Regulation and Carcinogenesis, National Cancer InstituteBuilding 41, Room C629, 41 Library Drive, MSC 5055, Bethesda, MD 20892-5055,USA.
Abstract
In the past three years, a novel signal transduction pathway downstream of thetransforming growth factor-beta (TGF-beta) superfamily receptor serine-threoninekinases has been shown to be mediated by a family of latent transcriptionfactors called 'Smads'. These proteins mediate a short-circuited pathway inwhich a set of receptor-activated Smads are phosphorylated directly by thereceptor kinase and then translocate to the nucleus complexed to the commonmediator, Smad4, to participate in transcriptional complexes. Smads 2 and 3mediate signals predominantly from the TGF-beta receptors. Of these, specificroles have been ascribed to Smad3 in control of chemotaxis of neutrophils andmacrophages and the inhibition of Smad3 activity by the oncogene Evi-1 suggeststhat it may play a role in leukemogenesis. Other data, such as the induction bythe inflammatory cytokine interferon-gamma of an inhibitory Smad, Smad7, whichblocks the actions of Smad3, suggest that identification of the specific genetargets of Smad proteins in immune cells will provide new insight into themechanisms of TGF-beta action on these cells.
PMID
10611754
|
 |