| Original Literature | Model OverView |
|---|---|
|
Publication
Title
How Toll-like receptors signal: what we know and what we don't know.
Affiliation
School of Biochemistry and Immunology, Trinity College Dublin, Ireland.laoneill@tcd.ie
Abstract
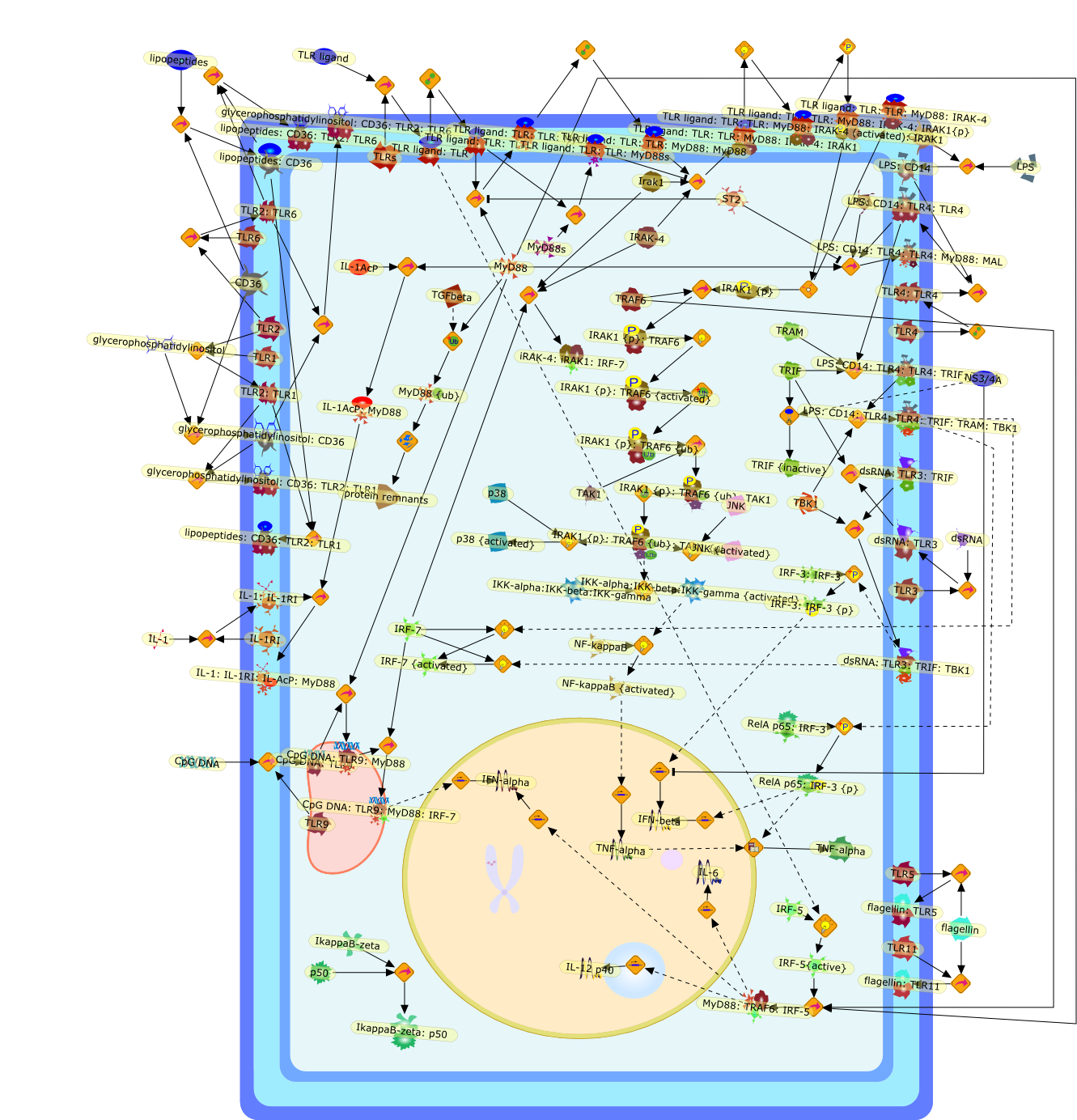
Signal transduction pathways activated by Toll-like receptors (TLRs) havecontinued to be a major focus of research for investigators interested in theinitiation of innate immune responses and the induction of pro-inflammatorycytokines and type I interferons during infection. Biochemical details of themajor signalling pathways have now been obtained, and the specific signallingpathways activated by different TLRs are being elucidated. New insights into theactivation of IRF family members, notably IRF3, IRF5 and IRF7, have beenobtained, and interesting spatiotemporal aspects of signalling by MyD88 leadingto IRF7 activation revealed. Adapters in TLR signalling are targets forinhibition, both by endogenous regulators and by virally derived proteins.Selective targeting of pathways by anti-inflammatory glucocorticoids alsoindicates the potential for modulating TLR signalling therapeutically.
PMID
16343886
|
 |