| Original Literature | Model OverView |
|---|---|
|
Publication
Title
Innate immune recognition of, and regulation by, DNA.
Affiliation
Exploratory Research for Advanced Technology (ERATO), Japan Science andTechnology Agency (JST) and Department of Host Defense, Research Institute forMicrobial Diseases, Osaka University, 3-1 Yamada-oka, Suita, Osaka 565-0871,Japan.
Abstract
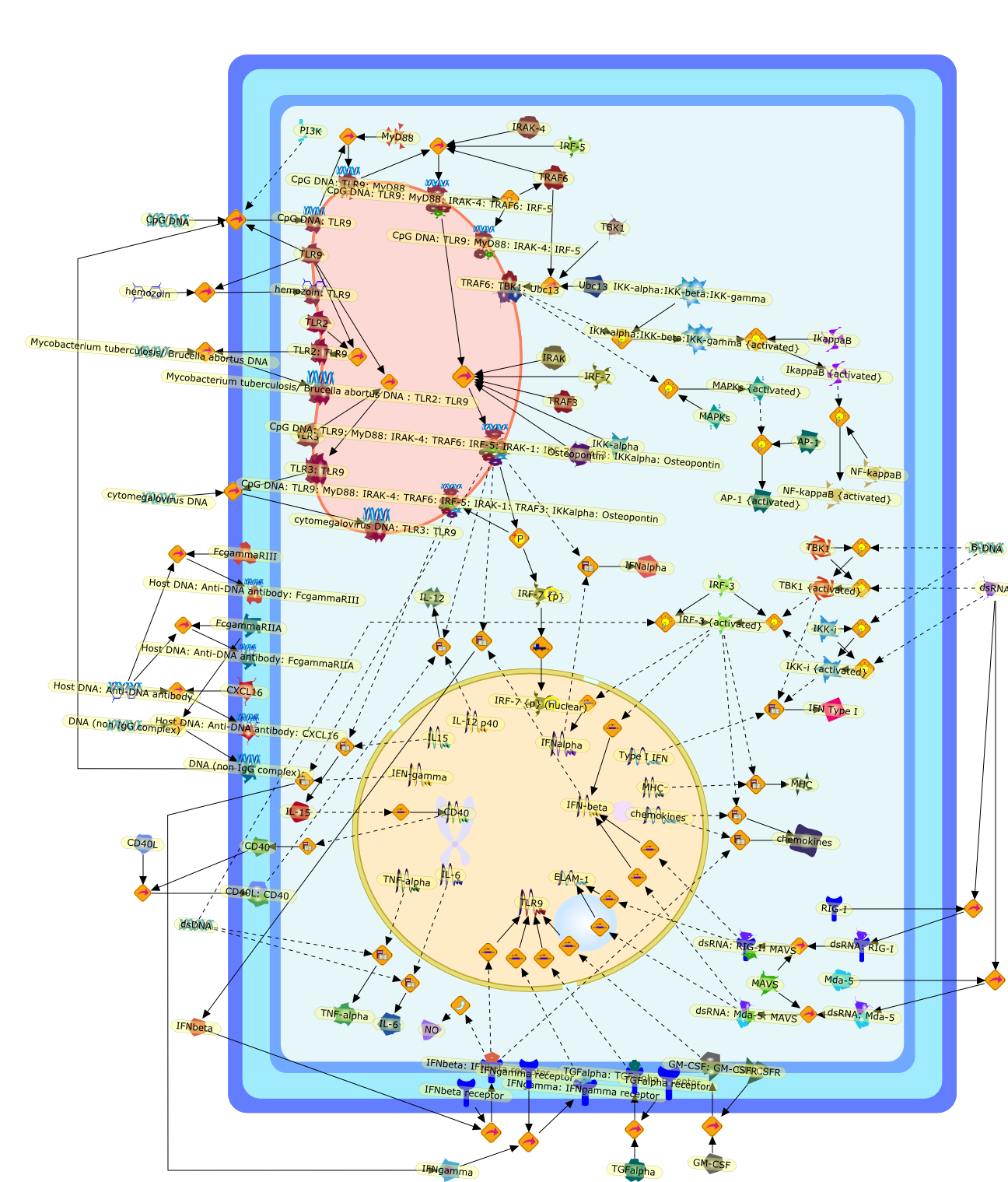
DNA in microbes or host cells is normally sequestered from the immune system,and therefore inert, but becomes an active immunostimulatory molecule duringinfection or tissue damage. Recent evidence suggests that Toll-like receptor(TLR)9, currently the only known immune sensor for DNA, recognizes more diverseelements in its ligand than initially thought, and must cooperate withadditional host factors to provoke an optimal innate immune response in thephysiological environment. Moreover, the innate immune system possesses aTLR9-independent, as-yet-undefined intracellular recognition machinery ofdouble-stranded DNA that induces type I interferons through distinct signalingpathways. TLR9-dependent and TLR9-independent immune recognition of DNA mightplay crucial roles in DNA-associated protective immunity and in pathologicalautoimmunity.
PMID
16979939
|
 |