| Original Literature | Model OverView |
|---|---|
|
Publication
Title
Translational mini-review series on Toll-like receptors: recent advances inunderstanding the role of Toll-like receptors in anti-viral immunity.
Affiliation
School of Biochemistry and Immunology, Trinity College Dublin, Ireland.agbowie@tcd.ie
Abstract
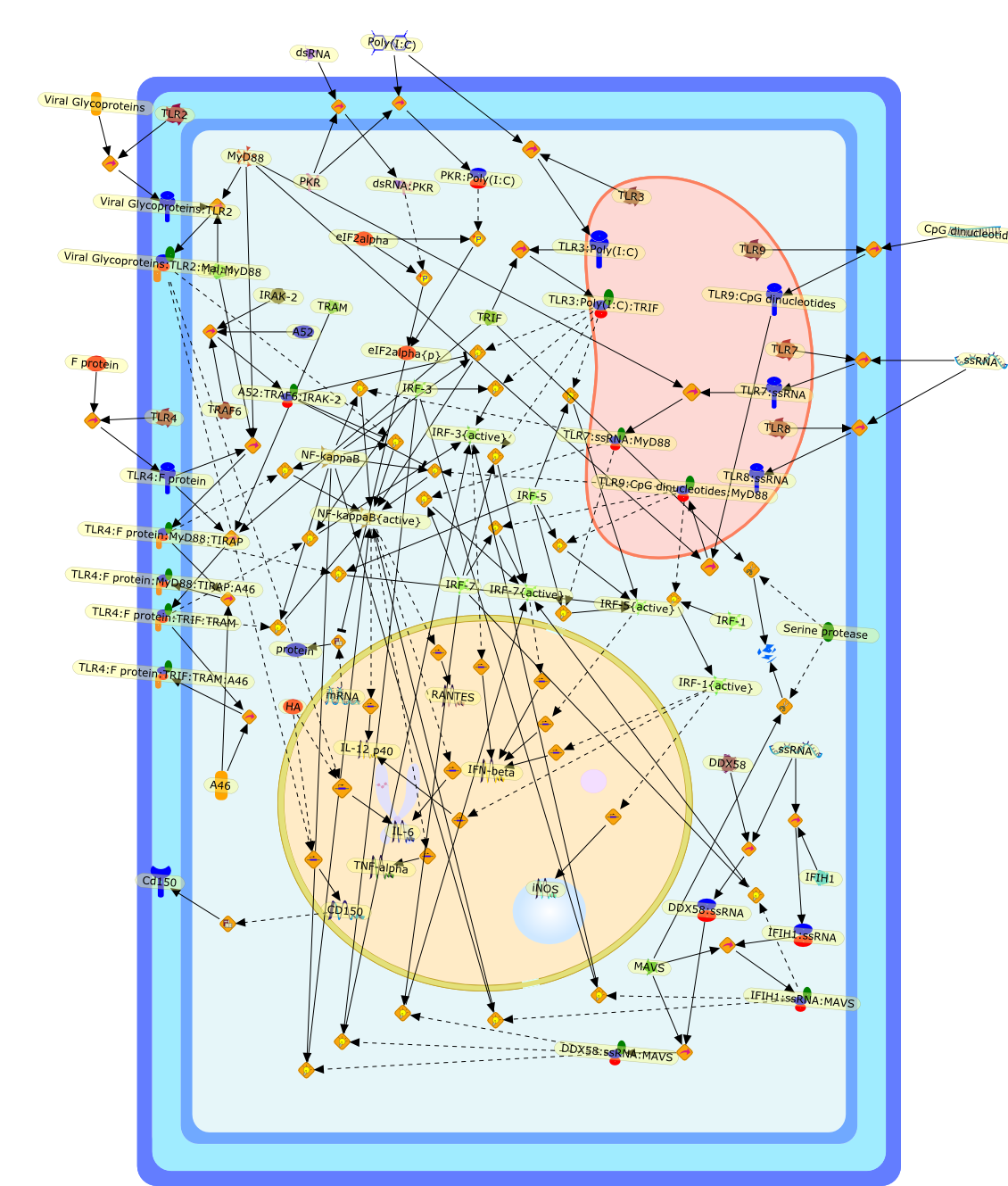
(TLRs) respond to pathogens to initiate the innate immune response and directadaptive immunity, and evidence to date suggests that they have a role in thedetection of viruses. Many viral macromolecules have been shown to activateanti-viral signalling pathways via TLRs, leading to the induction of cytokinesand interferons, while viruses also have means of not only evading detection byTLRs, but also of subverting these receptors for their own purposes. This reviewdiscusses the role of TLRs in the context of other known viral detectionsystems, and examines some of the often surprising results from studies usingmice deficient in TLRs and their adaptors, in an attempt to unravel theparticular contribution of TLRs to anti-viral immunity.
PMID
17223961
|
 |