| Original Literature | Model OverView |
|---|---|
|
Publication
Title
Genetic regulation of macrophage priming/activation: the Lsh gene story.
Affiliation
Department of Medical Parasitology, London School of Hygiene and TropicalMedicine, U.K.
Abstract
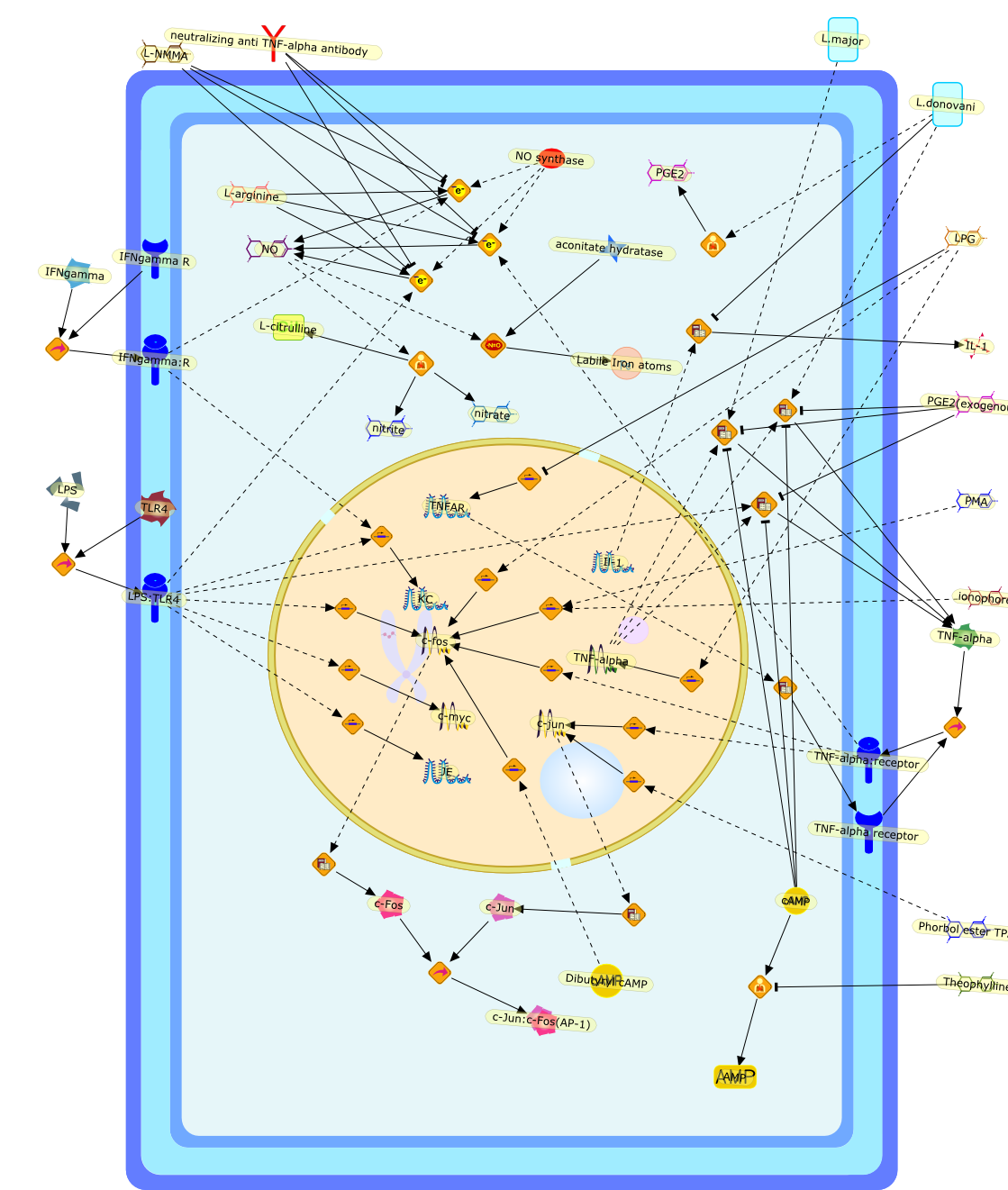
This paper describes functional and genetic studies on the macrophage resistancegene Lsh/Ity/Bcg first described almost two decades ago. Working in vitro withresident peritoneal, liver (Kupffer cells) and bone marrow derived macrophagesfrom congenic B10 (LshS) and B10.L-LshR mice it has been possible to demonstratethat the final effector mechanism for the gene in regulating antileishmanialactivity involves production of reactive nitrogen rather than reactive oxygenintermediates. This in turn is dependent upon priming/activation of macrophagesfor enhanced TNF-alpha release which acts back on the macrophage in an autocrinemanner to increase nitric oxide production. The precise point at which Lsh actsto control macrophage priming/activation has not been identified, but studies ofearly response gene expression show differences in KC mRNA levels at 2 h afterLPS stimulation, and in c-fos mRNA as early as 20 min after stimulation with PMAplus ionophore, in peritoneal macrophages from congenic LshS and LshR mice. Dataavailable suggest that both negative and positive signals may be involved inmacrophage priming/activation, with LshS macrophages down-regulating theircapacity for continued response to the autocrine loop. Work in progress willexamine the role of TPA and cAMP response element-binding proteins in regulatinggene expression in Lsh congenic mice. A major new initiative has also commencedto clone the Lsh gene by reverse genetics using yeast artificial chromosomes towalk towards Lsh from the closet proximal and distal markers on mouse chromosome1.(ABSTRACT TRUNCATED AT 250 WORDS)
PMID
1757110
|
 |