| Original Literature | Model OverView |
|---|---|
|
Publication
Title
Glucocorticoids and the innate immune system: crosstalk with the toll-likereceptor signaling network.
Affiliation
Hospital for Special Surgery, Department of Microbiology & Immunology, WeillMedical College of Cornell University, 535 E70th Street, Research Building Room425, New York, NY 10021,USA.
Abstract
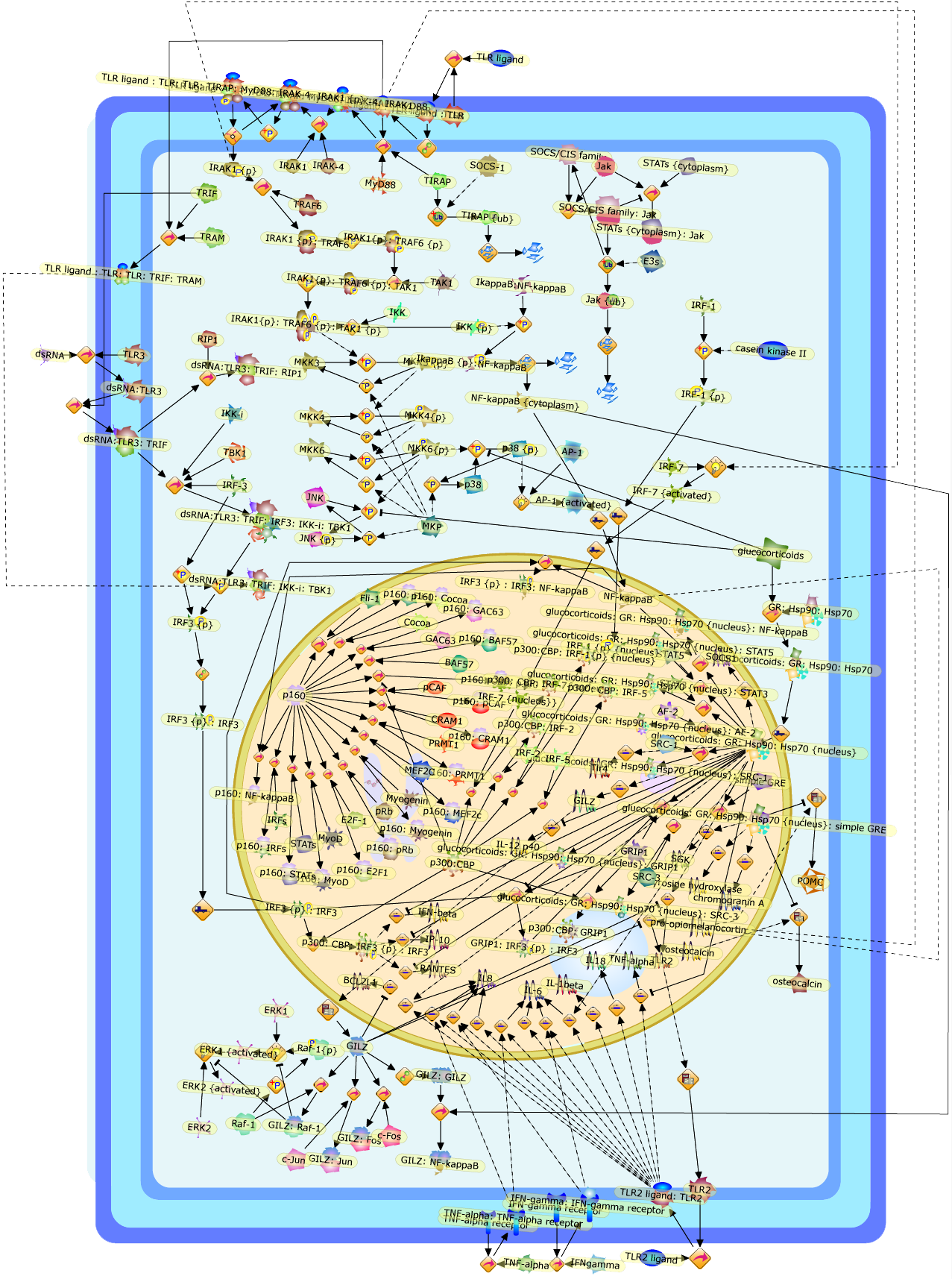
Toll-like receptors (TLRs) are responsible for the recognition of a variety ofmicrobial pathogens and the initial induction of immune and inflammatoryresponses. These responses are normally restricted by the adrenally producedglucocorticoid hormones which provide a feedback mechanism to curb unabatedinflammation. Glucocorticoids act through a ligand-dependent transcriptionfactor-the glucocorticoid receptor (GR), which engages in a complex network ofprotein:protein and protein:DNA interactions ultimately activating or repressingtarget gene transcription. Not surprisingly, multiple mechanisms account for theglucocorticoid interference with TLR signaling including enhanced expression ofthe natural inhibitors of TLR pathways, direct repression of TLR-activatedtranscriptional regulators and cross-utilization of cofactors essential for bothGR and TLR signaling. Here we discuss recent and unexpected examples ofcrosstalk between the two transcriptional networks and the emerging role of GRin the regulation of innate immunity.
PMID
17576036
|
 |