| Original Literature | Model OverView |
|---|---|
|
Publication
Title
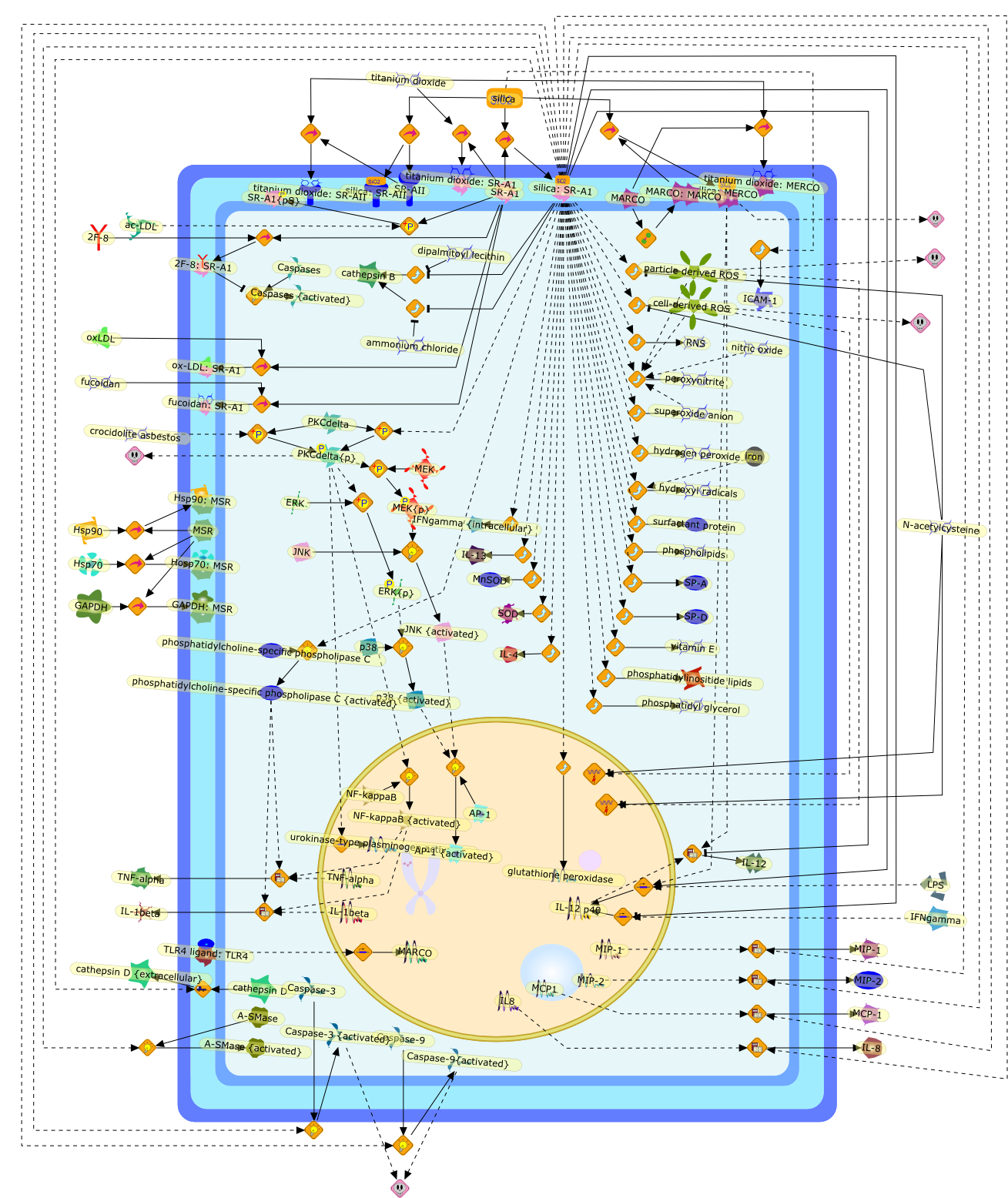
Silica binding and toxicity in alveolar macrophages.
Affiliation
Department of Biomedical and Pharmaceutical Sciences, Center for EnvironmentalHealth Sciences, University of Montana, 32 Campus Drive, SB 154, Missoula, MT59812, USA.
Abstract
Inhalation of the crystalline form of silica is associated with a variety ofpathologies, from acute lung inflammation to silicosis, in addition toautoimmune disorders and cancer. Basic science investigators looking at themechanisms involved with the earliest initiators of disease are focused on howthe alveolar macrophage interacts with the inhaled silica particle and theconsequences of silica-induced toxicity on the cellular level. Based onexperimental results, several rationales have been developed for exactly howcrystalline silica particles are toxic to the macrophage cell that isfunctionally responsible for clearance of the foreign particle. For example,silica is capable of producing reactive oxygen species (ROS) either directly (onthe particle surface) or indirectly (produced by the cell as a response tosilica), triggering cell-signaling pathways initiating cytokine release andapoptosis. With murine macrophages, reactive nitrogen species are produced inthe initial respiratory burst in addition to ROS. An alternative explanation forsilica toxicity includes lysosomal permeability, by which silica disrupts thenormal internalization process leading to cytokine release and cell death. Stillother research has focused on the cell surface receptors (collectively known asscavenger receptors) involved in silica binding and internalization. Thesilica-induced cytokine release and apoptosis are described as the function ofreceptor-mediated signaling rather than free radical damage. Current researchideas on silica toxicity and binding in the alveolar macrophage are reviewed anddiscussed.
PMID
18226603
|
 |