{kind=link}
{kind=link}


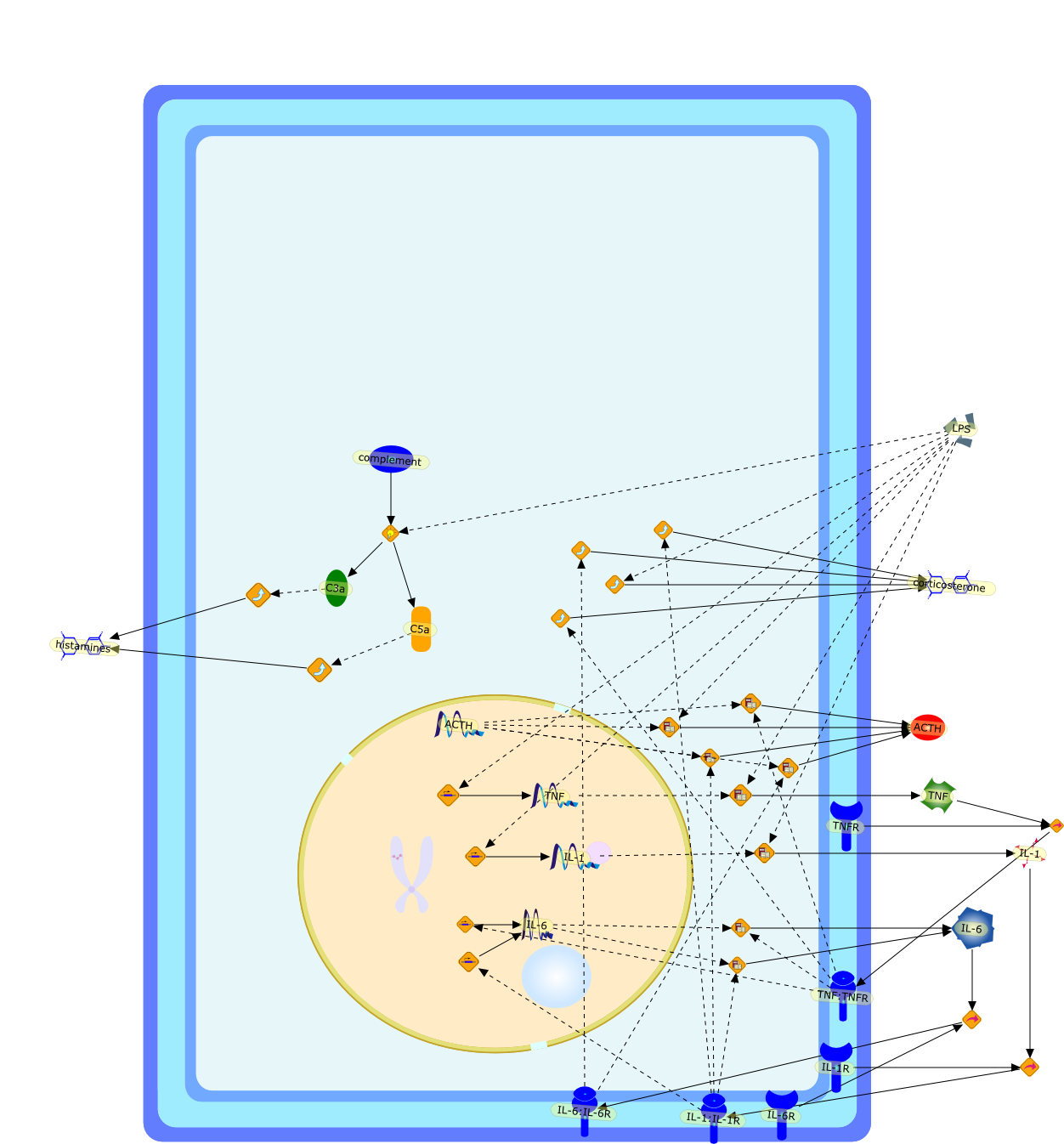
ACTH,C3a,C5a,IL-1,IL-1:IL-1R,IL-1R,IL-6,IL-6:IL-6R,IL-6R,LPS,TNF,TNF:TNFR,TNFR,complement,corticosterone,histamines
With the recent discovery and cloning of the lipopolysaccharide-binding protein(LBP), the "adapter-molecule" for LPS-binding to the cell surface receptor CD14was found. The ligand-receptor pair LPS/LBP-CD14 seems to be one importantelement in LPS-mediated activation of monocytic cells and possibly granulocytesand B cells. Here, some of the known functions of the proteins involved, LBP andCD14, are reviewed in the context of other endotoxin recognition studies, andthe outlook for ongoing and future investigations is described.
{kind=link}
{kind=link}
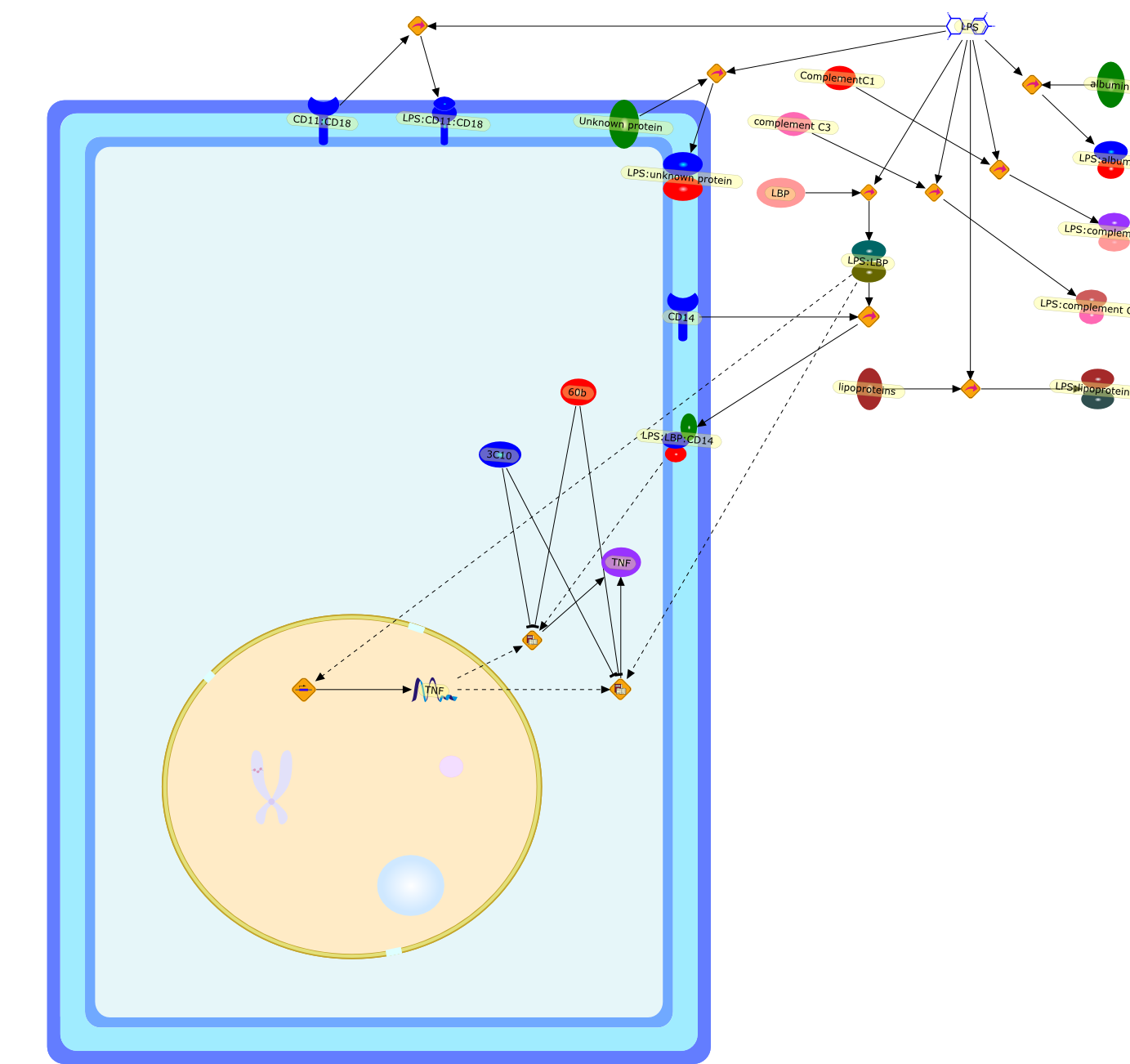
3C10,60b,CD11:CD18,CD14,ComplementC1,LBP,LPS,LPS:CD11:CD18,LPS:LBP,LPS:LBP:CD14,LPS:albumin,LPS:complement C3,LPS:complementC1,LPS:lipoproteins,LPS:unknown protein,TNF,Unknown protein,albumin,complement C3,lipoproteins
Activation of T cells is initiated by the recognition of antigen on antigenpresenting cells to exert the effector functions in immune and inflammatoryresponses. Two types of helper T cell (Th) clones (Th1 and Th2) are defined onthe basis of different patterns of cytokine (lymphokine) secretion. Theydetermine the outcome of an antigenic response toward humoral or cell-mediatedimmunity. Although lymphokine genes are coordinately regulated upon antigenstimulation, they are regulated by the mechanisms common to all as well as thosewhich are unique to each gene. For most lymphokine genes, a combination ofphorbol esters (phorbol 12-myristate 13 acetate, PMA) and calcium ionophores(A23187) is required for their maximal induction. Yet phorbol ester alone orcalcium ionophore alone produce several lymphokines. The production of thegranulocyte-macrophage colony stimulating factor (GM-CSF) is completelydependent on the two signals. We have previously found a cis-acting regionspanning the GM-CSF promoter region (positions -95 to +27) that confersinducibility to reporter genes in transient transfection assays. Furtheranalysis identified three elements required for efficient induction, referred toas GM2, GC-box and conserved lymphokine element (CLE0). GM2 defines a bindingsite for protein(s) whose binding is inducible by PMA. One protein, NF-GM2 issimilar to the transcription factor NF-kB. GC-box is a binding site forconstitutively bound proteins. CLEO defines a binding site for protein(s) whoseoptimum binding is stimulated by PMA and A23187. Viral trans-activators such asTax (human T cell leukemia virus-1, HTLV-1) and E2 (bovine papilloma virus, BPV)proteins are other agents which activate lymphokine gene expression by bypassingT cell receptor (TCR) mediated signaling. The trans-activation domain of E2 andTax is interchangeable although they have no obvious sequence homology betweenthem. The viral trans-activators appear to target specific DNA binding proteinsuch as NF-kB and Sp1 to cis-acting DNA site and promote lymphokine geneexpression without TCR-mediated stimulation.
{kind=link}
{kind=link}
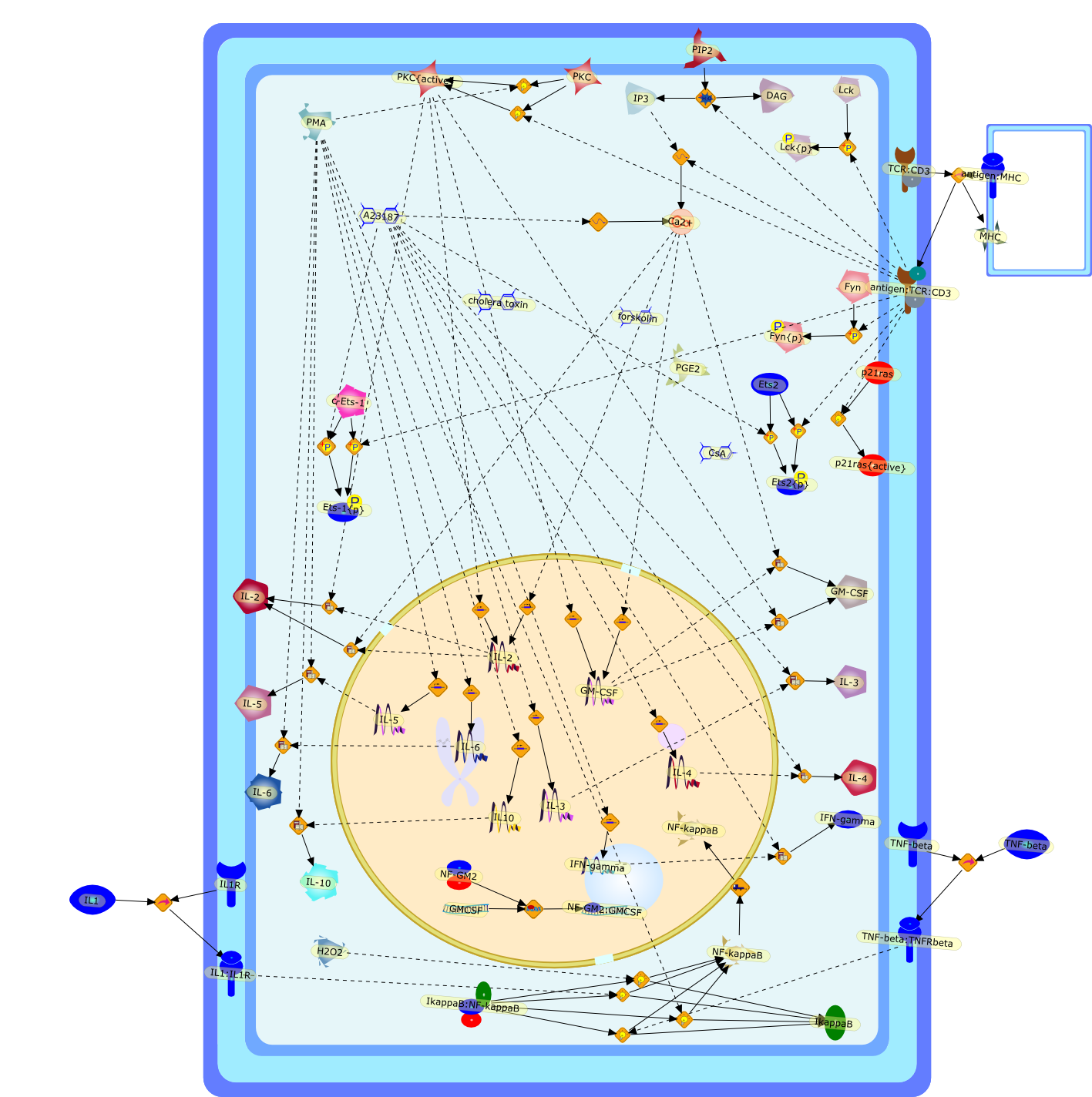
A23187,Ca2+,CsA,DAG,Ets-1{p},Ets2,Ets2{p},Fyn,Fyn{p},GM-CSF,GMCSF,H2O2,IFN-gamma,IL-10,IL-2,IL-3,IL-4,IL-5,IL-6,IL1,IL10,IL1:IL1R,IL1R,IP3,IkappaB,IkappaB:NF-kappaB,Lck,Lck{p},MHC,NF-GM2,NF-GM2:GMCSF,NF-kappaB,PGE2,PIP2,PKC,PKC{active},PMA,TCR:CD3,TNF-beta,TNF-beta:TNFRbeta,antigen:MHC,antigen:TCR:CD3,c-Ets-1,cholera toxin,forskolin,p21ras,p21ras{active}
This paper describes functional and genetic studies on the macrophage resistancegene Lsh/Ity/Bcg first described almost two decades ago. Working in vitro withresident peritoneal, liver (Kupffer cells) and bone marrow derived macrophagesfrom congenic B10 (LshS) and B10.L-LshR mice it has been possible to demonstratethat the final effector mechanism for the gene in regulating antileishmanialactivity involves production of reactive nitrogen rather than reactive oxygenintermediates. This in turn is dependent upon priming/activation of macrophagesfor enhanced TNF-alpha release which acts back on the macrophage in an autocrinemanner to increase nitric oxide production. The precise point at which Lsh actsto control macrophage priming/activation has not been identified, but studies ofearly response gene expression show differences in KC mRNA levels at 2 h afterLPS stimulation, and in c-fos mRNA as early as 20 min after stimulation with PMAplus ionophore, in peritoneal macrophages from congenic LshS and LshR mice. Dataavailable suggest that both negative and positive signals may be involved inmacrophage priming/activation, with LshS macrophages down-regulating theircapacity for continued response to the autocrine loop. Work in progress willexamine the role of TPA and cAMP response element-binding proteins in regulatinggene expression in Lsh congenic mice. A major new initiative has also commencedto clone the Lsh gene by reverse genetics using yeast artificial chromosomes towalk towards Lsh from the closet proximal and distal markers on mouse chromosome1.(ABSTRACT TRUNCATED AT 250 WORDS)
{kind=link}
{kind=link}
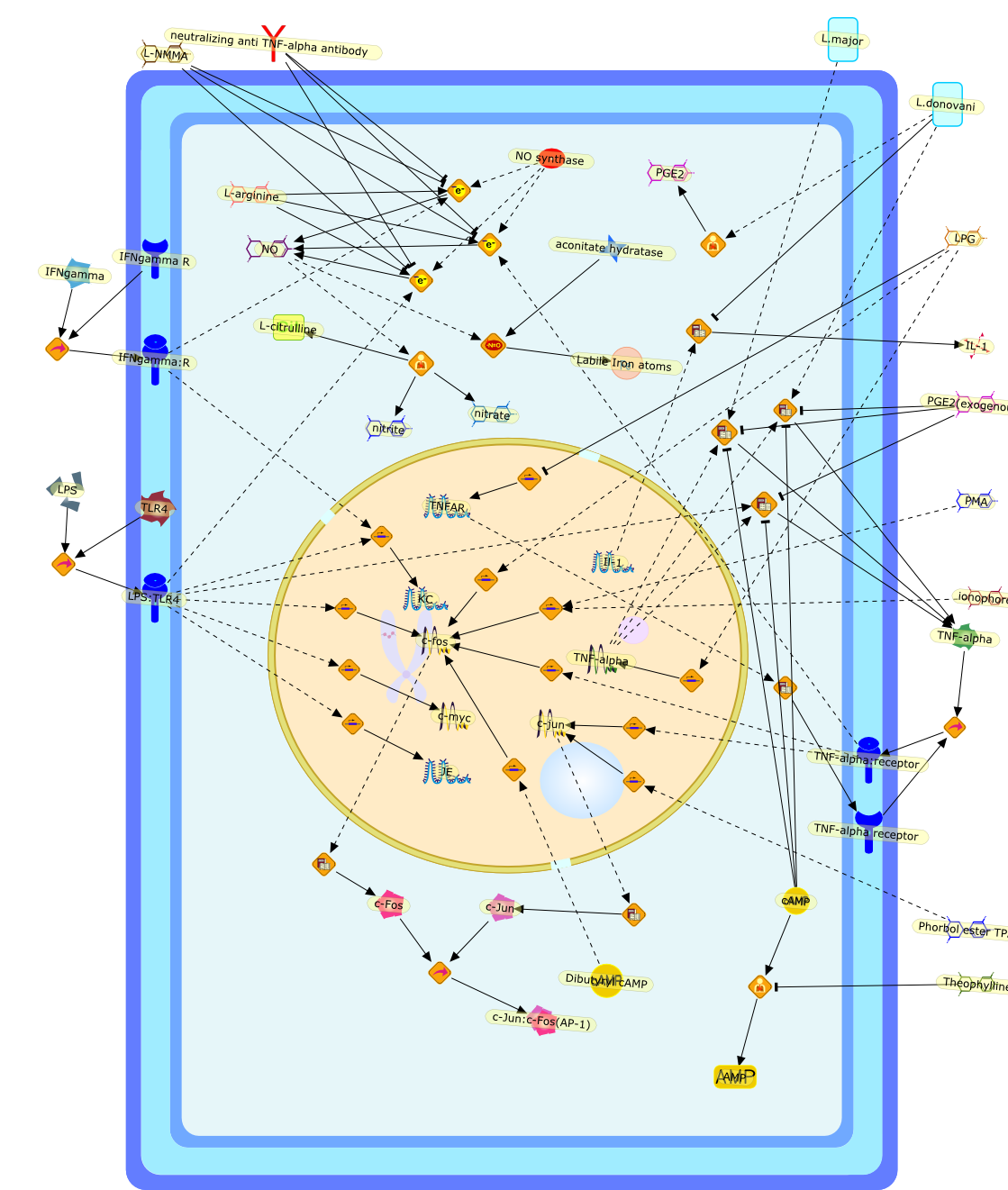
AMP,Dibutyryl cAMP,IFNgamma,IFNgamma R,IFNgamma:R,IL-1,Il-1,JE,KC,L-NMMA,L-arginine,L-citrulline,L.donovani,L.major,LPG,LPS,LPS:TLR4,Labile Iron atoms,NO,NO synthase,PGE2,PGE2(exogenous),PMA,Phorbol ester TPA,TLR4,TNF-alpha,TNF-alpha receptor,TNF-alpha:receptor,TNFAR,Theophylline,aconitate hydratase,c-Fos,c-Jun,c-Jun:c-Fos(AP-1),c-fos,c-jun,c-myc,cAMP,ionophore,neutralizing anti TNF-alpha antibody,nitrate,nitrite
The lipid A domain of lipopolysaccharide (LPS) is a unique, glucosamine-basedphospholipid that makes up the outer monolayer of the outer membrane of mostgram-negative bacteria. Because of its profound pharmacological effects onanimal cells, especially those of the immune system, lipid A is also known asendotoxin. Despite decades of earlier work, the precise chemistry of endotoxinsand the biochemical pathways for their enzymatic synthesis have been elucidatedonly within the past 5 years. In this review, we summarize the essentials ofendotoxin biochemistry and also present recent experiments aimed at identifyingsurface receptors, signal-transducing elements, transcriptional factors, and keyintracellular targets involved in the response of animal cells to endotoxins.
{kind=link}
{kind=link}
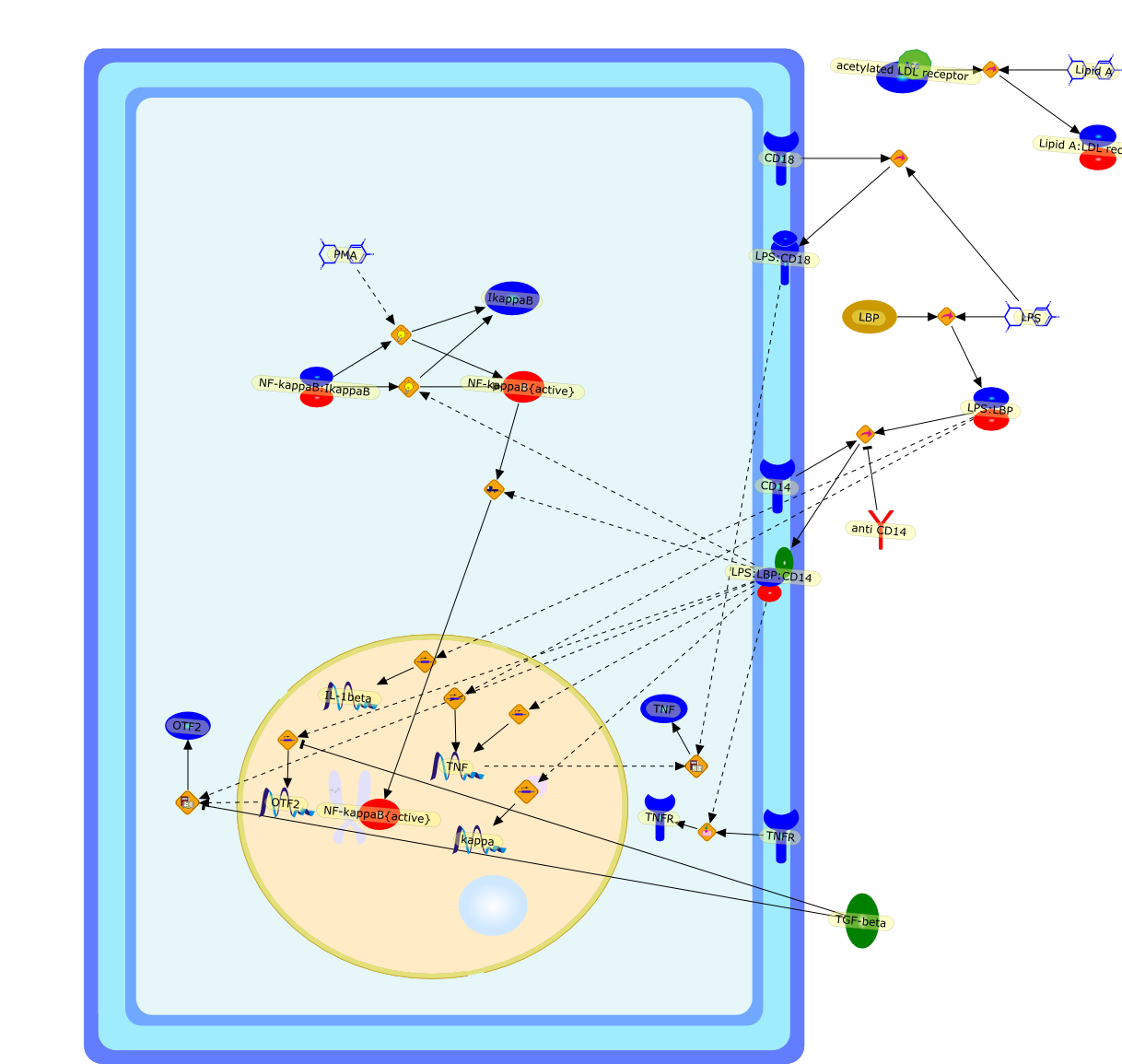
CD14,CD18,IL-1beta,IkappaB,LBP,LPS,LPS:CD18,LPS:LBP,LPS:LBP:CD14,Lipid A,Lipid A:LDL receptor,NF-kappaB:IkappaB,NF-kappaB{active},OTF2,PMA,TGF-beta,TNF,TNFR,acetylated LDL receptor,anti CD14,kappa
Lipopolysaccharide (LPS) or endotoxin elicits a broad, non-specific cascade ofevents in vivo, resulting in secretion of a variety of potent mediators andcytokines produced primarily by activated macrophages and monocytes. Theoverproduction of these effector molecules, such as interleukin-1 and tumornecrosis factor-alpha, contributes to the pathophysiology of endotoxic shock.Cellular recognition of LPS involves several different molecules, includingcluster of differentiation antigen CD14. A thorough understanding of theinteraction of LPS with cells of the immune system is necessary before effectivepreventative or therapeutic measures can be designed to limit the host responseto endotoxin. This review discusses the role of CD14 and other LPS-recognitionmolecules in LPS-mediated macrophage activation.
{kind=link}
{kind=link}
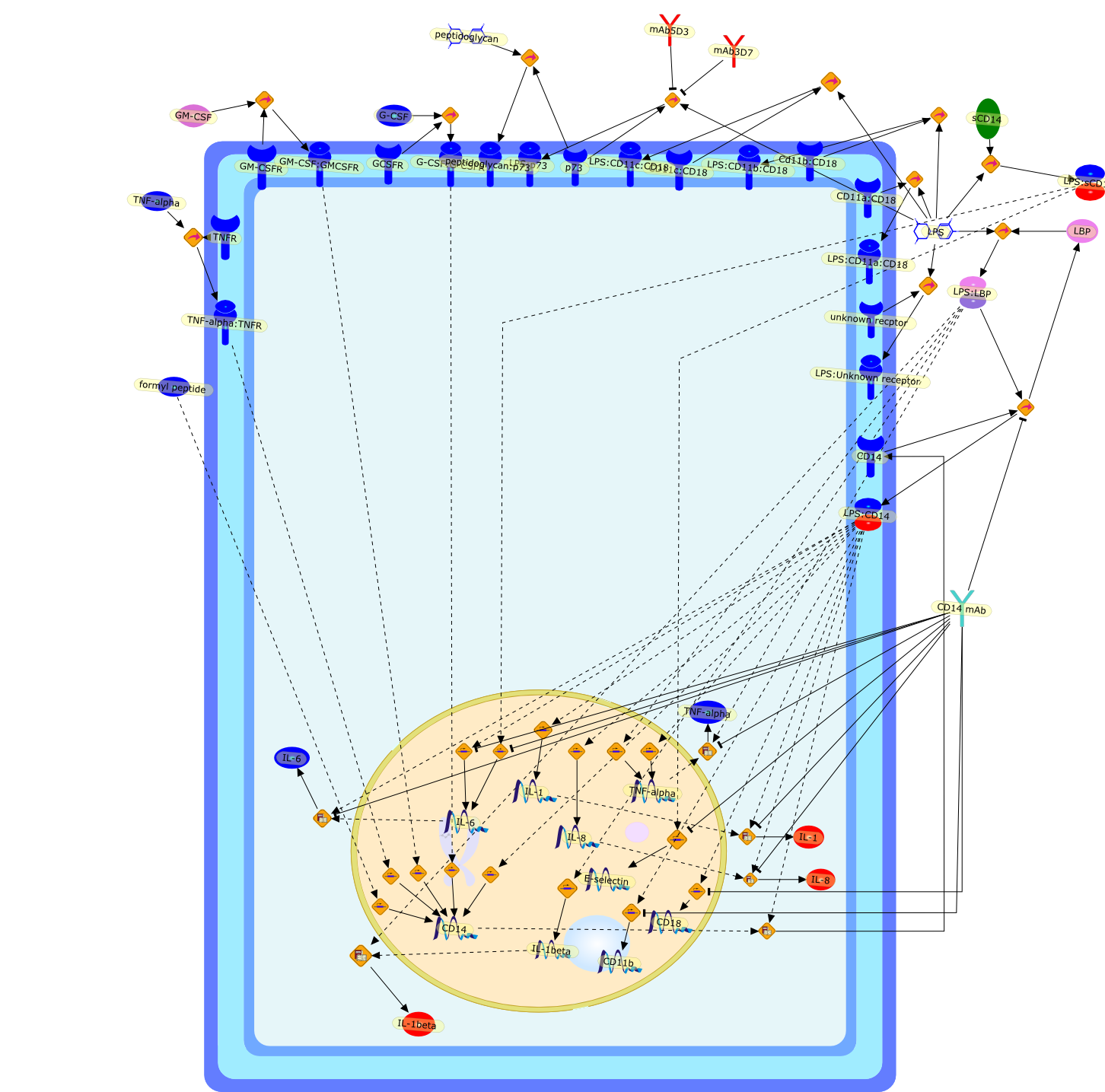
CD11a:CD18,CD11b,CD11c:CD18,CD14,CD14 mAb,CD18,Cd11b:CD18,E-selectin,G-CSF,G-CSF:G-CSFR,GCSFR,GM-CSF,GM-CSF:GMCSFR,GM-CSFR,IL-1,IL-1beta,IL-6,IL-8,LBP,LPS,LPS:CD11a:CD18,LPS:CD11b:CD18,LPS:CD11c:CD18,LPS:CD14,LPS:LBP,LPS:Unknown receptor,LPS:p73,LPS:sCD14,TNF-alpha,TNF-alpha:TNFR,TNFR,formyl peptide,mAb3D7,mAb5D3,p73,peptidoglycan,peptidoglycan:p73,sCD14,unknown recptor
{kind=link}
{kind=link}
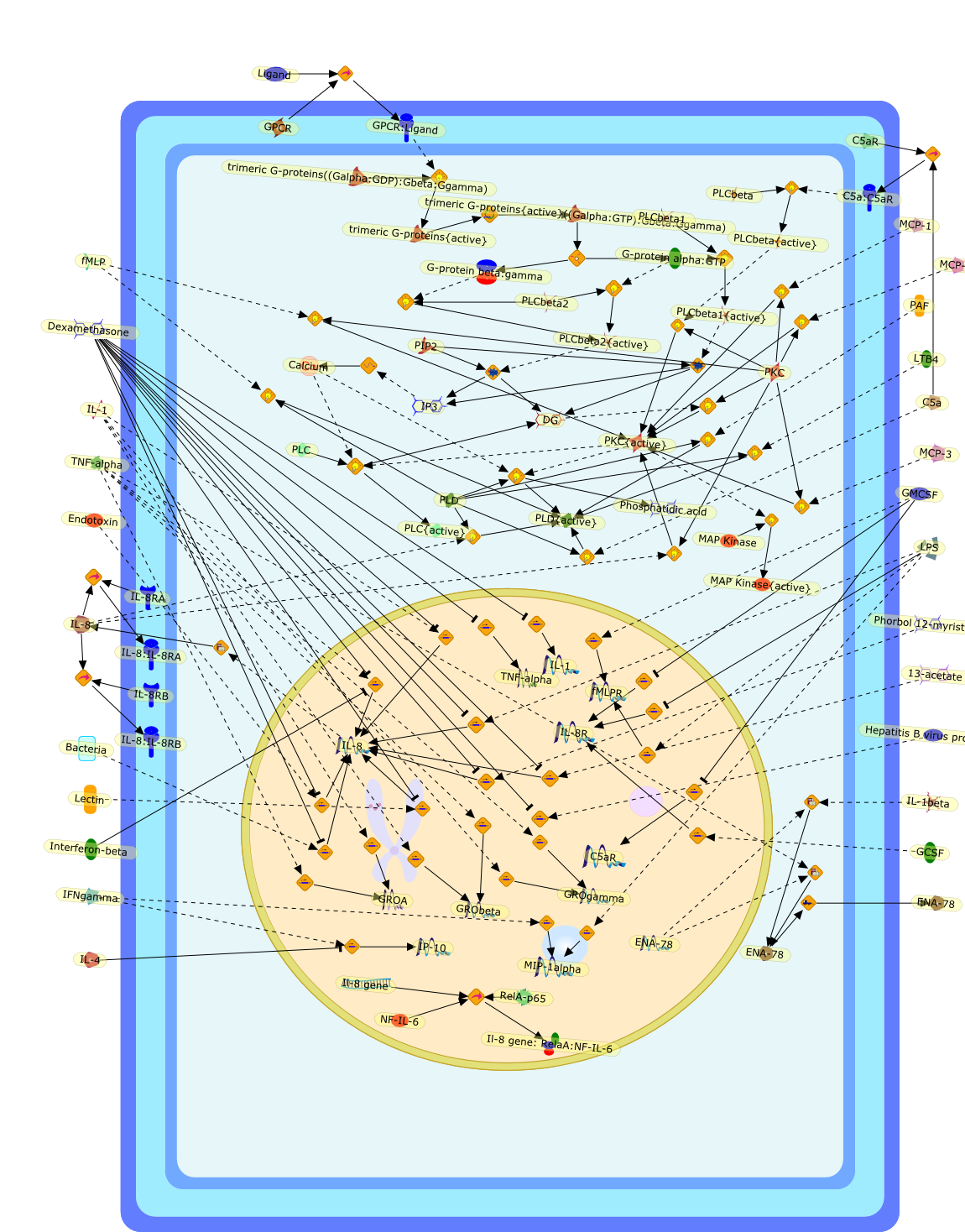
13-acetate,Bacteria,C5a,C5a:C5aR,C5aR,Calcium,DG,Dexamethasone,ENA-78,Endotoxin,G-protein alpha:GTP,G-protein beta:gamma,GCSF,GMCSF,GPCR,GPCR:Ligand,GROA,GRObeta,GROgamma,Hepatitis B virus protein X,IFNgamma,IL-1,IL-1beta,IL-4,IL-8,IL-8:IL-8RA,IL-8:IL-8RB,IL-8R,IL-8RA,IL-8RB,IP-10,IP3,Il-8 gene,Il-8 gene: RelaA:NF-IL-6,Interferon-beta,LPS,LTB4,Lectin,Ligand,MAP Kinase,MAP Kinase{active},MCP-1,MCP-2,MCP-3,MIP-1alpha,NF-IL-6,PAF,PIP2,PKC,PKC{active},PLC,PLCbeta,PLCbeta1,PLCbeta1{active},PLCbeta2,PLCbeta2{active},PLCbeta{active},PLC{active},PLD,PLD{active},Phorbol 12-myristate,Phosphatidic acid,RelA-p65,TNF-alpha,fMLP,fMLPR,trimeric G-proteins((Galpha:GDP):Gbeta:Ggamma),trimeric G-proteins{active},trimeric G-proteins{active}((Galpha:GTP):Gbeta:Ggamma)
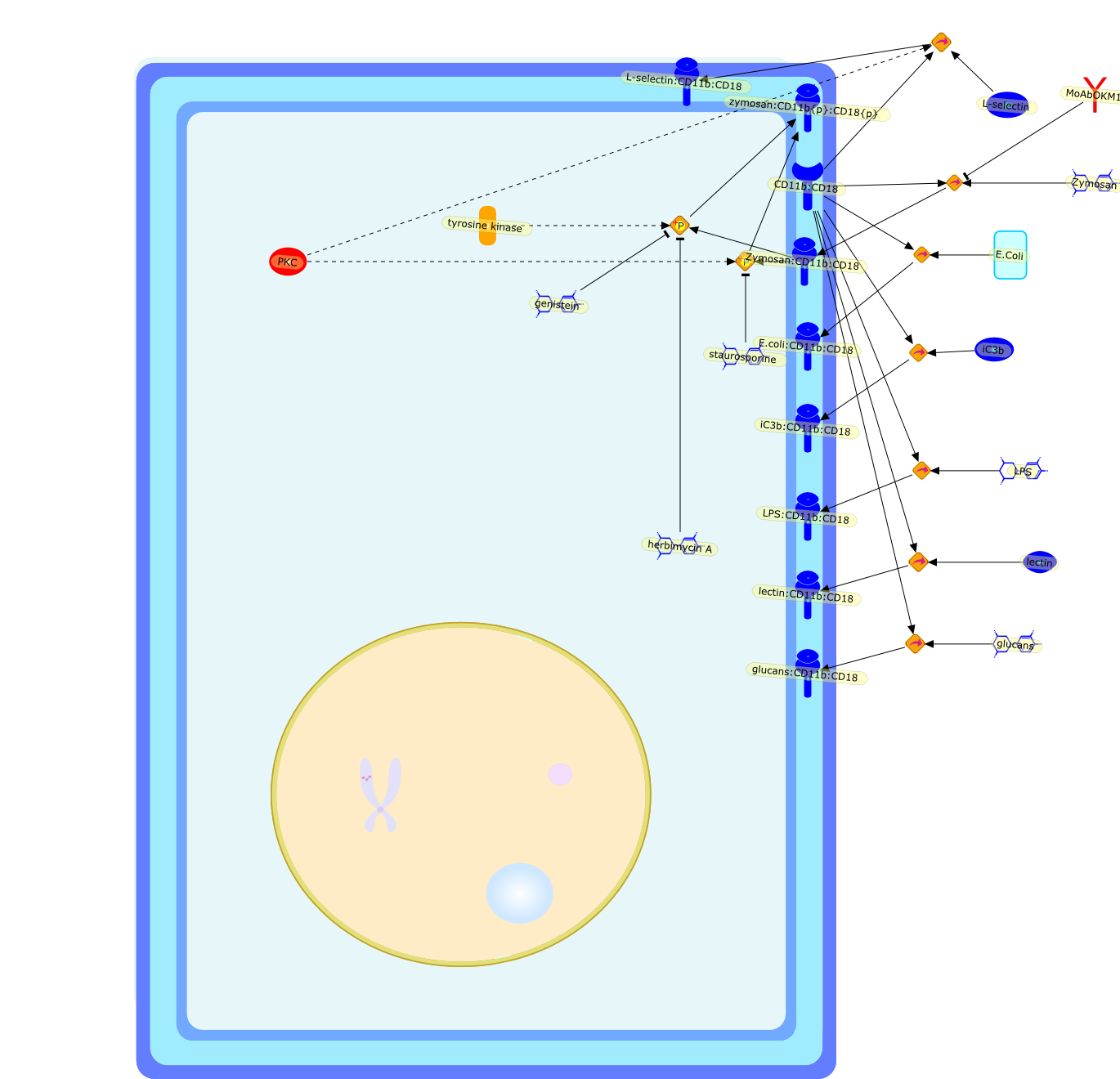
The C3 receptor CR3 is expressed on phagocytic cells, minor subsets of B and Tcells, and natural killer (NK) cells. It has important functions both as anadhesion molecule and a membrane receptor mediating recognition of diverseligands such as intercellular adhesion molecule-1 (ICAM-1) and fixed iC3b. Thereceptor is capable of undergoing an activation event that regulates both itsspecificity for various ligands and its ability to mediate phagocytosis orextracellular cytotoxicity. Certain bacteria express carbohydrates orlipopolysaccharides (LPS) that can bind to and activate CR3, allowing thereceptor to assume its activated state. Soluble beta-glucan derived from theyeast Saccharomyces cerevisiae is a particularly potent stimulator of CR3, andproduces an activated state of the receptor that permits neutrophil phagocytosisof iC3b-coated erythrocytes or NK, cell cytotoxicity of iC3b-coated tumourcells, that are normally resistant to NK cells.
{kind=link}
{kind=link}
CD11b:CD18,E.Coli,E.coli:CD11b:CD18,L-selectin,L-selectin:CD11b:CD18,LPS,LPS:CD11b:CD18,MoAbOKM1,PKC,Zymosan,Zymosan:CD11b:CD18,genistein,glucans,glucans:CD11b:CD18,herbimycin A,iC3b,iC3b:CD11b:CD18,lectin,lectin:CD11b:CD18,staurosporine,tyrosine kinase,zymosan:CD11b{p}:CD18{p}
Through its action on macrophages, bacterial lipopolysaccharide (LPS) orendotoxin can trigger responses that are protective or injurious to the host.This review examines the effects of LPS on macrophages by following events fromthe cell surface to the nucleus. The involvement of protein tyrosine kinases,mitogen-activated protein kinases, protein kinase C, G proteins, protein kinaseA, ceramide-activated protein kinase, and microtubules in this process arereviewed. At the nuclear level, rel, C/EBP, Ets, Egr, fos, and jun familymembers have been implicated in activation of LPS-inducible gene expression.
{kind=link}
{kind=link}
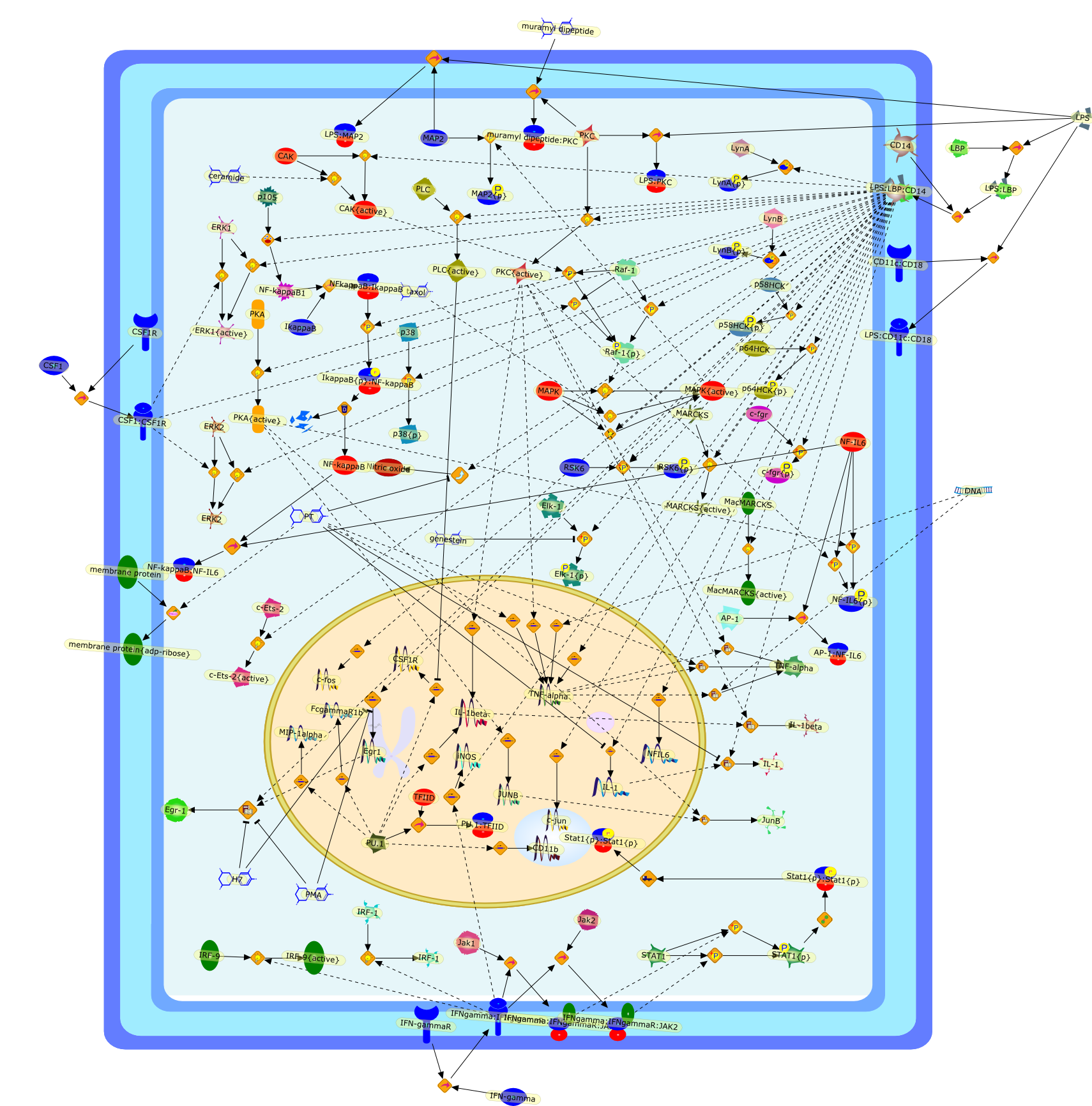
AP-1,AP-1:NF-IL6,CAK,CAK{active},CD11b,CD11c:CD18,CD14,CSF1,CSF1:CSF1R,CSF1R,DNA,ERK1,ERK1{active},ERK2,Egr-1,Egr1,Elk-1,Elk-1{p},FcgammaR1b,H7,IFN-gamma,IFN-gammaR,IFNgamma:IFNgammaR,IFNgamma:IFNgammaR:JAK1,IFNgamma:IFNgammaR:JAK2,IL-1,IL-1beta,IRF-1,IRF-9,IRF-9{active},IkappaB,IkappaB{p}:NF-kappaB,JUNB,Jak1,Jak2,JunB,LBP,LPS,LPS:CD11c:CD18,LPS:LBP,LPS:LBP:CD14,LPS:MAP2,LPS:PKC,LynA,LynA{p},LynB,LynB{p},MAP2,MAP2{p},MAPK,MAPK{active},MARCKS,MARCKS{active},MIP-1alpha,MacMARCKS,MacMARCKS{active},NF-IL6,NF-IL6{p},NF-kappaB,NF-kappaB1,NF-kappaB:NF-IL6,NFIL6,NFkappaB:IkappaB,Nitric oxide,PKA,PKA{active},PKC,PKC{active},PLC,PLC{active},PMA,PT,PU.1,PU.1:TFIID,RSK6,RSK6{p},Raf-1,Raf-1{p},STAT1,STAT1{p},Stat1{p}:Stat1{p},TFIID,TNF-alpha,c-Ets-2,c-Ets-2{active},c-fgr,c-fgr{p},c-fos,c-jun,ceramide,genestein,iNOS,membrane protein,membrane protein{adp-ribose},muramyl dipeptide,muramyl dipeptide:PKC,p105,p38,p38{p},p58HCK,p58HCK{p},p64HCK,p64HCK{p},protein remnants,taxol
Control of cell proliferation involves a finely interwoven network of positiveand negative cell cycle regulators. Signal transduction pathways linking c-fms(CSF-1R) to cellular proliferation and differentiation are being explored. Partof the strategy is to use a series of G1 inhibitors to help pinpoint relevanttargets. Several inhibitors-8Br-cAMP, interferon gamma (IFN gamma), INFalpha/beta, lipopolysaccharide (LPS), tumor necrosis factor-alpha (TNF alpha),and dimethylamiloride-suppress CSF-1-stimulated proliferation in murine bonemarrow-derived macrophages (BMM) even when added in the mid- to late-G1 phase ofthe cell cycle. The down-modulating effects of the inhibitors on the expressionof the following cell cycle regulators have been examined: c-myc, cyclin D1 andD2, cdk4, Rb phosphorylation, E2F binding activity, ribonucleotide reductasesubunits, and PCNA. Some differences in the negative control of such regulatorswere found, for example, in the manner in which IFN gamma and cAMP down-regulatec-myc expression. Using blocking antibodies and BMM from type I IFN receptorknockout mice, it appears that one of these inhibitors, IFN alpha/beta, acts asan endogenous inhibitor in CSF-1-treated BMM and is also responsible, at leastin part, for the inhibition of cell cycle progression by LPS and TNF alpha.Another strategy has been to attempt to relate early biochemical changes inducedby CSF-1 to later changes in the G1 phase, partly by studying cycling versusnoncycling macrophages and partly by using cells expressing c-fms with tyrosinemutations in the intracytoplasmic region. CSF-1-mediated effects on thefollowing signal transduction molecules in these systems will be described:PI3-kinase, myelin basic protein kinases, Erks, and STAT transcription factors.
{kind=link}
{kind=link}
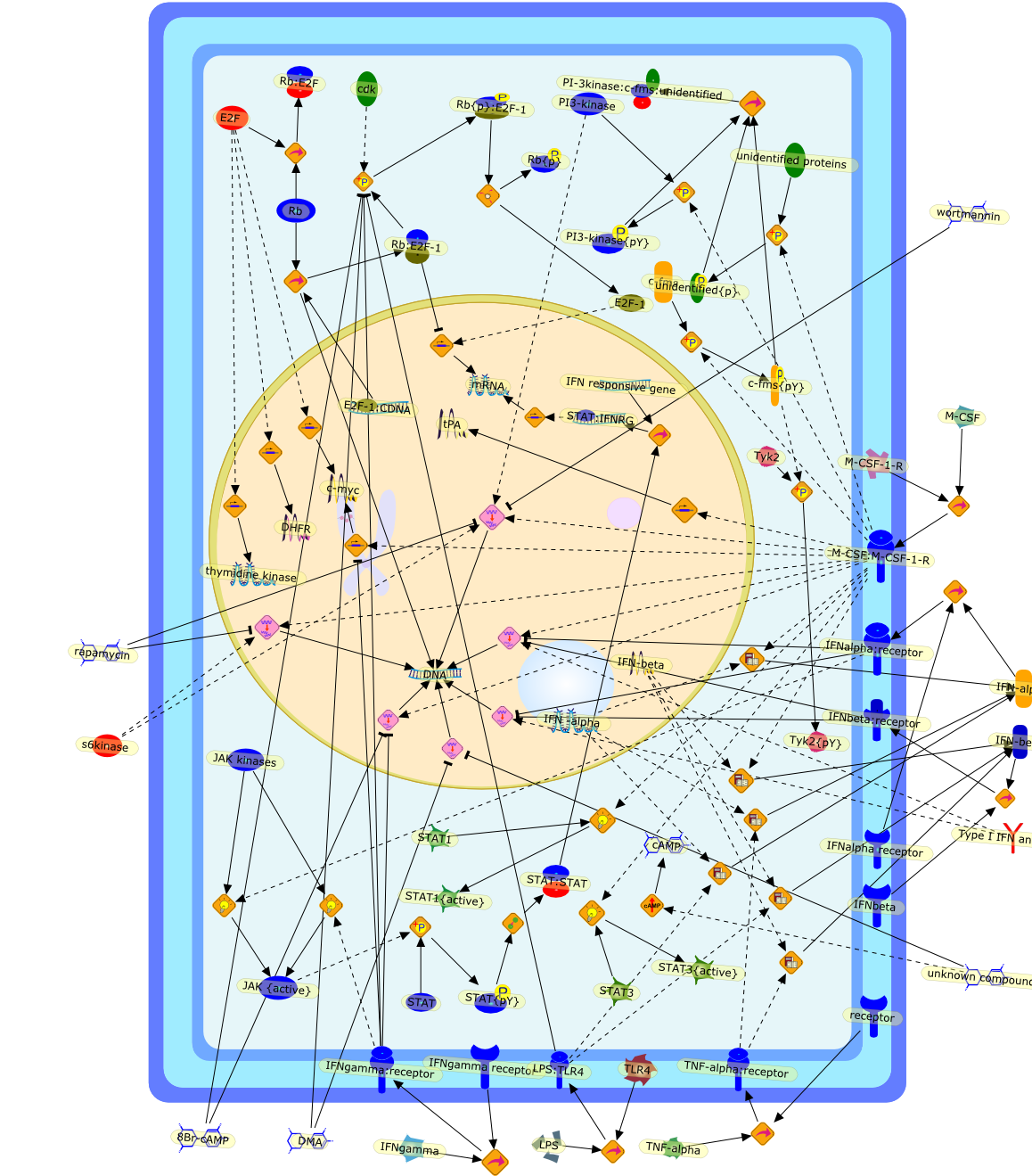
IFN -alpha,8Br-cAMP,DHFR,DMA,DNA,E2F ,E2F-1,E2F-1:CDNA,IFN responsive gene,IFN-alpha,IFN-beta,IFNalpha receptor,IFNalpha:receptor,IFNbeta,IFNbeta:receptor,IFNgamma,IFNgamma receptor,IFNgamma:receptor,JAK kinases,JAK {active},LPS,LPS:TLR4,M-CSF,M-CSF-1-R,M-CSF:M-CSF-1-R,PI-3kinase:c-fms:unidentified,PI3-kinase,PI3-kinase{pY},Rb,Rb:E2F,Rb:E2F-1,Rb{p},Rb{p}:E2F-1,STAT,STAT1,STAT1{active},STAT3,STAT3{active},STAT:IFNRG,STAT:STAT,STAT{pY},TLR4,TNF-alpha,TNF-alpha:receptor,Tyk2,Tyk2{pY},Type I IFN antibody,c-fms,c-fms{pY},c-myc,cAMP,cdk,mRNA,rapamycin,receptor,s6kinase,tPA,thymidine kinase,unidentified proteins,unidentified{p},unknown compounds,wortmannin
{kind=link}
{kind=link}
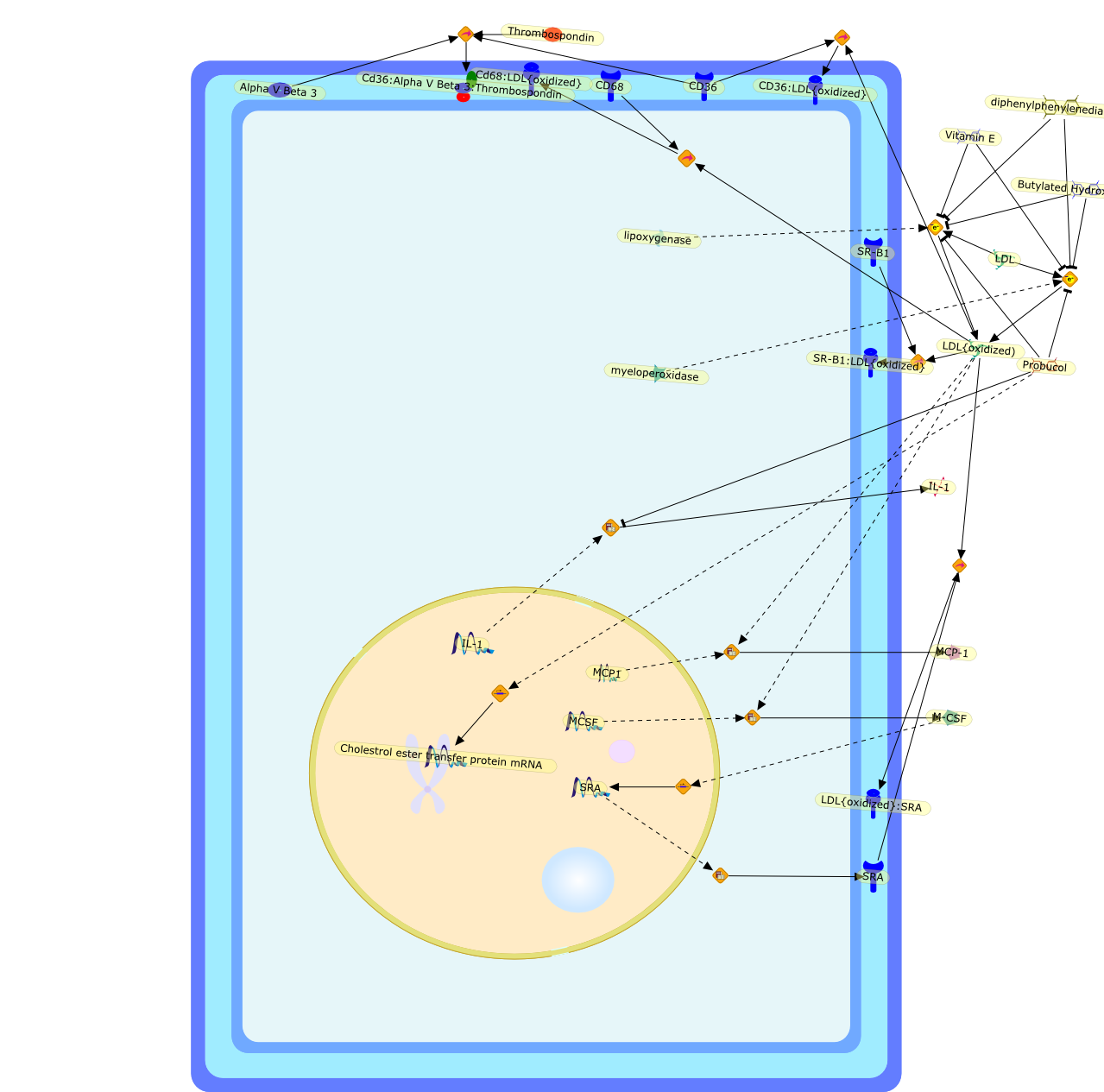
Alpha V Beta 3,Butylated Hydroxytoulene,CD36,CD36:LDL{oxidized},CD68,Cd36:Alpha V Beta 3:Thrombospondin,Cd68:LDL{oxidized},Cholestrol ester transfer protein mRNA,IL-1,LDL,LDL{oxidized),LDL{oxidized}:SRA,M-CSF,MCP-1,MCP1,MCSF,Probucol,SR-B1,SR-B1:LDL{oxidized},SRA,Thrombospondin,Vitamin E,diphenylphenylenediamine,lipoxygenase,myeloperoxidase
During the pathogenesis of atherosclerosis, inflammatory cells such as themonocyte-derived macrophage accumulate in the vessel wall where they releasecytokines. Initially, cytokines may assist in CE removal of lipoprotein-derivedcholesterol/CE hydrolysis to clear intracellular lipid. When plasma levels ofLDL become elevated, the vessel wall becomes lipid-engorged over time because itis unable to traffick the large amounts of endocytosed LDL-CE from the cell. Inaddition, lipoprotein entrapment by the extracellular matrix can lead to theprogressive oxidation of LDL because of the action of lipoxygenases, reactiveoxygen species, peroxynitrite, and/or myeloperoxidase. A range of oxidized LDLspecies is thus generated, ultimately resulting in their delivery to vascularcells through several families of scavenger receptors (Fig 1). These molecularTrojan horses and cellular saboteurs once formed or deposited in the cell cancontribute to, and participate in, formation of macrophage- and smoothmuscle-derived foam cells. A lipid-enriched fatty streak along the vessel wallcan ensue. In addition to foam cell development, products of LDL peroxidationmay activate endothelial cells, increase smooth muscle mitogenesis, or induceapoptosis because of the effects of oxysterols and products of lipidperoxidation (Fig 1). Because antioxidant defenses may be limited in themicroenvironment of the cell or within LDL, the oxidation process continues toprogress. Enzymes associated with HDL such as PAF acetylhydrolase andparaoxonase can participate in the elimination of biologically active lipids,but diminished cellular antioxidant activity coupled with low levels of HDL mayallow acceleration of the clinical course of vascular disease. There is stillmuch to be learned about how modified LDL initiate cellular signals that lead toinflammation, mitosis, or cholesterol accumulation. The present challengesinclude elucidation of the key signaling events that regulatelipoprotein-derived cholesterol trafficking in the vessel wall, which can impacton the pathogenesis of vascular disease.
{kind=link}
{kind=link}
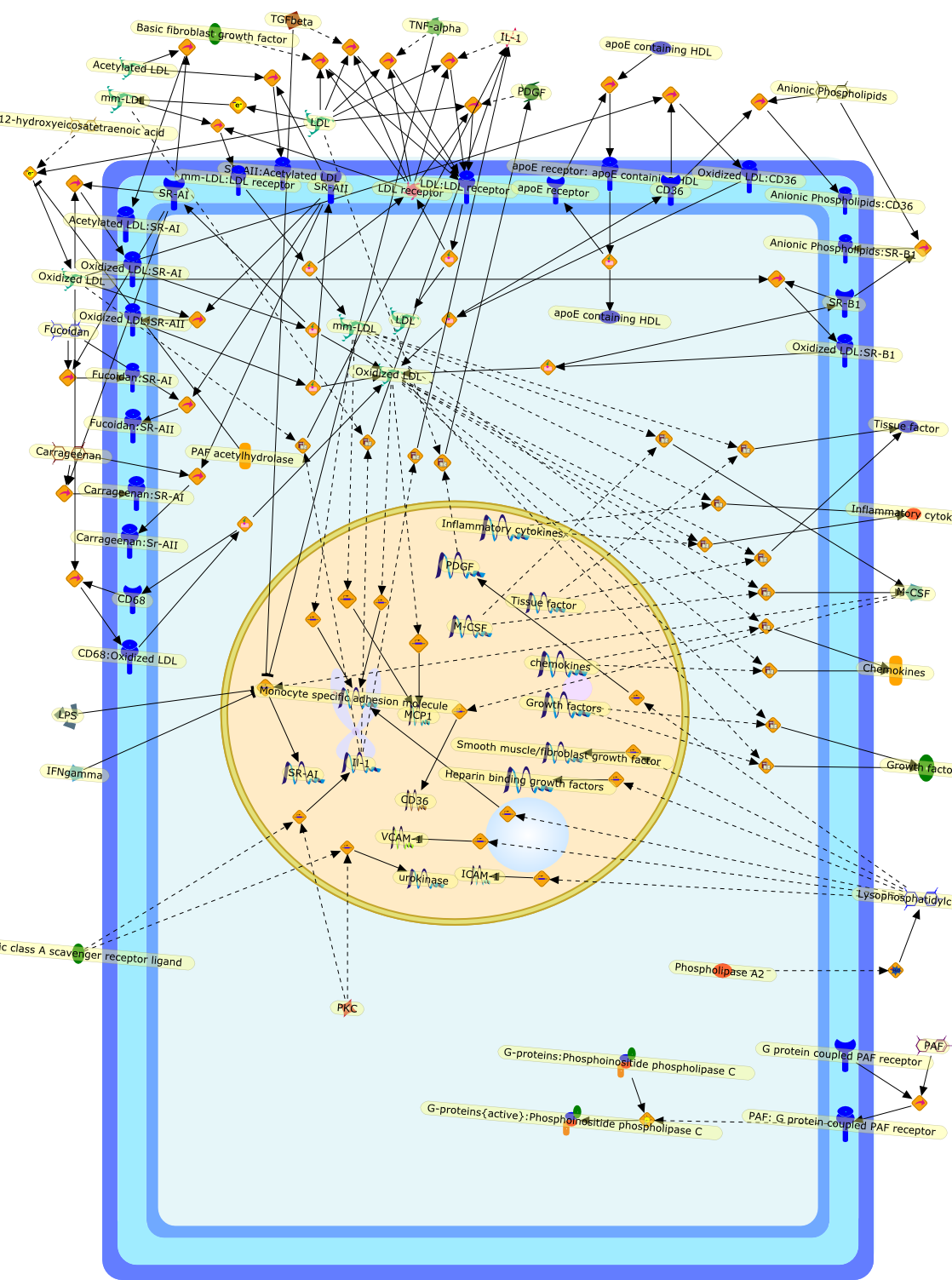
12-hydroxyeicosatetraenoic acid,Acetylated LDL,Acetylated LDL:SR-AI,Anionic Phospholipids,Anionic Phospholipids:CD36,Anionic Phospholipids:SR-B1,Basic fibroblast growth factor,CD36,CD68,CD68:Oxidized LDL,Carrageenan,Carrageenan:SR-AI,Carrageenan:Sr-AII,Chemokines,Fucoidan,Fucoidan:SR-AI,Fucoidan:SR-AII,G protein coupled PAF receptor,G-proteins:Phosphoinositide phospholipase C,G-proteins{active}:Phosphoinositide phospholipase C,Growth factors,Heparin binding growth factors,ICAM-1,IFNgamma,IL-1,Il-1,Inflammatory cytokines,LDL,LDL receptor,LDL:LDL receptor,LPS,Lysophosphatidylcholine,M-CSF,MCP1,Monocyte specific adhesion molecule,Oxidized LDL,Oxidized LDL:CD36,Oxidized LDL:SR-AI,Oxidized LDL:SR-AII,Oxidized LDL:SR-B1,PAF,PAF acetylhydrolase,PAF: G protein coupled PAF receptor,PDGF,PKC,Phospholipase A2,SR-AI,SR-AII,SR-AII:Acetylated LDL,SR-B1,Smooth muscle/fibroblast growth factor,Specific class A scavenger receptor ligand,TGFbeta,TNF-alpha,Tissue factor,VCAM-1,apoE containing HDL,apoE receptor,apoE receptor: apoE containing HDL,chemokines,mm-LDL,mm-LDL:LDL receptor,urokinase
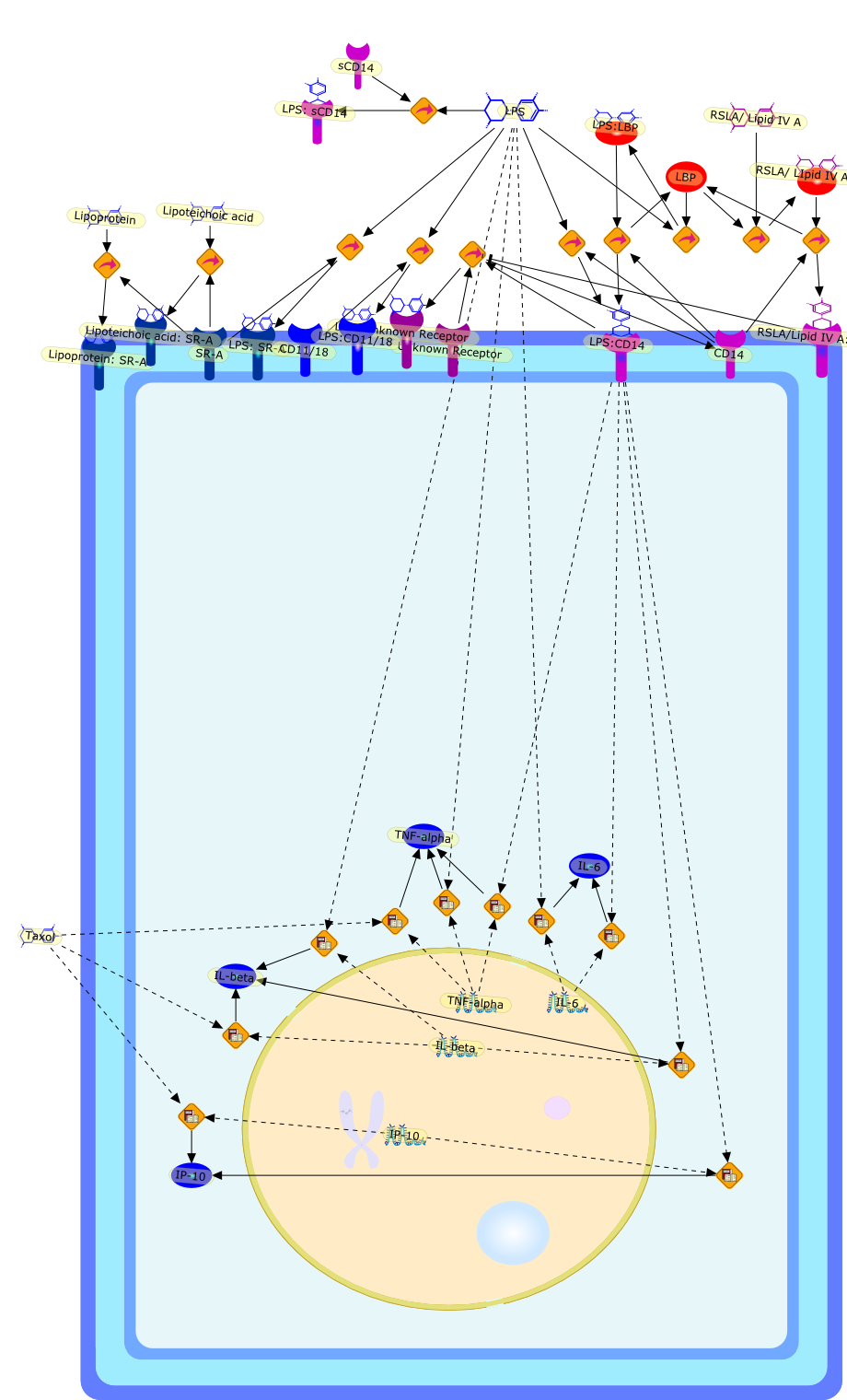
Macrophage activation by gram-negative lipopolysaccharide (LPS) has beenextensively studied in an attempt to define the mechanisms that underlie innateimmunity against bacterial pathogens. Dysregulation of these same mechanismscontributes to the pathophysiological consequences of bacterial sepsis. Thebiological actions of LPS are mediated, at least in part, by both LPS-bindingproteins and LPS receptors. Several LPS receptors (CD14, the macrophagescavenger receptor, and the beta2 integrins), as well as the serum LPS-bindingprotein LBP, have been cloned and studied in detail. In addition, insightsgained through the use of LPS antagonists have led to a better understanding ofa molecule believed to function in conjunction with LPS receptors to transducesignals from the membrane to the cytosol. More recently, the use of knockoutmice has greatly expanded our knowledge of the biology of LPS receptors andbinding proteins. This review will summarize various phenotypes of mice thatlack genes encoding CD14, the scavenger receptor, and LBP. These knockout micehave revealed several unexpected features of LPS action in vivo. Together, theseanimal models may provide a means to develop and evaluate novel therapeuticapproaches to the control of endotoxin shock.
{kind=link}
{kind=link}
CD11/18,CD14,IL-6,IL-beta,IP-10,LBP,LPS,LPS: SR-A,LPS: Unknown Receptor,LPS: sCD14,LPS:CD11/18,LPS:CD14,LPS:LBP,Lipoprotein,Lipoprotein: SR-A,Lipoteichoic acid,Lipoteichoic acid: SR-A,RSLA/ LIpid IV A:LBP,RSLA/ Lipid IV A,RSLA/Lipid IV A: CD14,SR-A,TNF-alpha,Taxol,Unknown Receptor,sCD14
{kind=link}
{kind=link}
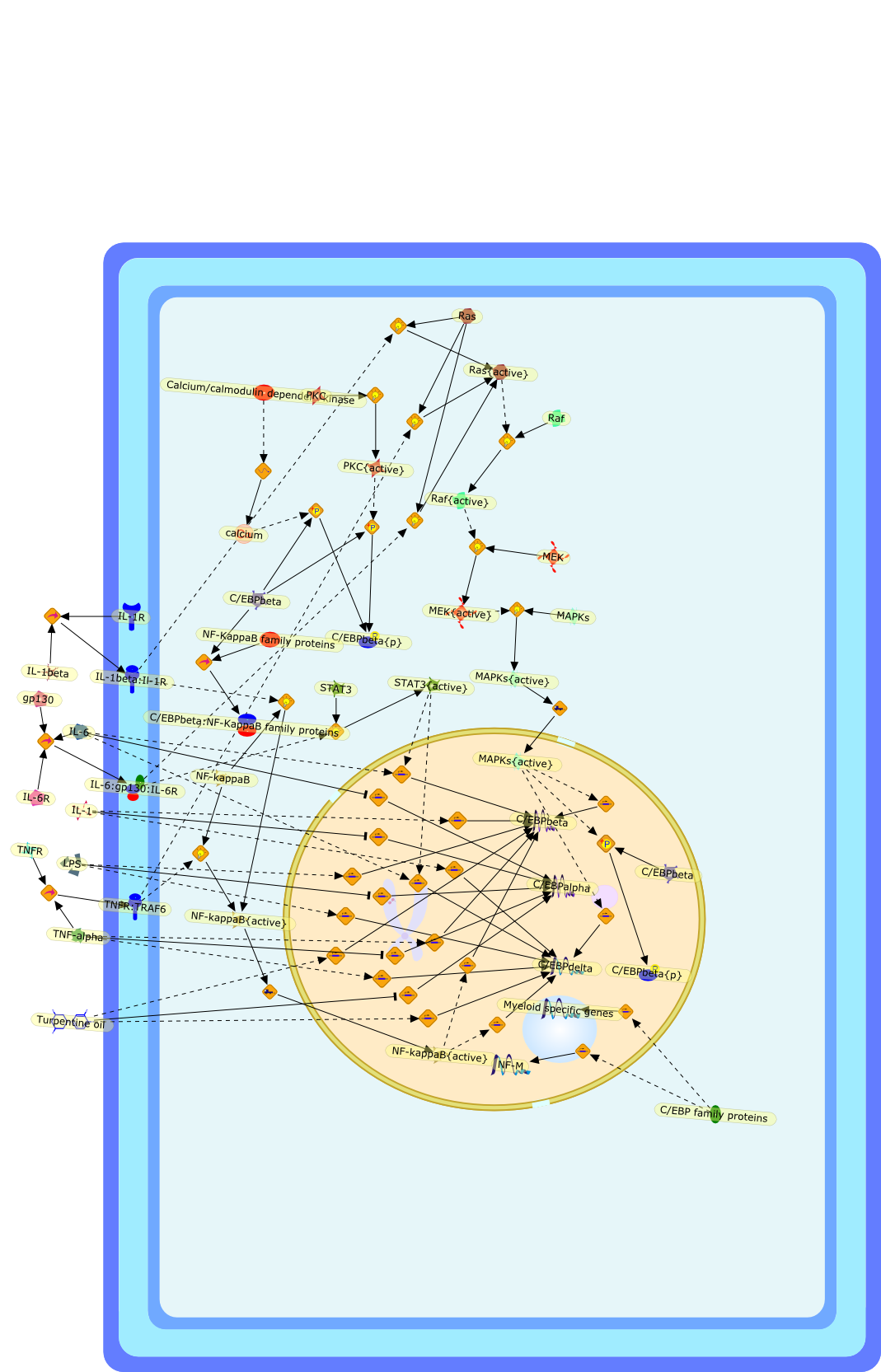
C/EBP family proteins,C/EBPalpha,C/EBPbeta,C/EBPbeta:NF-KappaB family proteins,C/EBPbeta{p},C/EBPdelta,Calcium/calmodulin dependent kinase,IL-1,IL-1R,IL-1beta,IL-1beta:Il-1R,IL-6,IL-6:gp130:IL-6R,IL-6R,LPS,MAPKs,MAPKs{active},MEK,MEK{active},Myeloid specific genes,NF-KappaB family proteins,NF-M,NF-kappaB,NF-kappaB{active},PKC,PKC{active},Raf,Raf{active},Ras,Ras{active},STAT3,STAT3{active},TNF-alpha,TNFR,TNFR:TRAF6,Turpentine oil,calcium,gp130
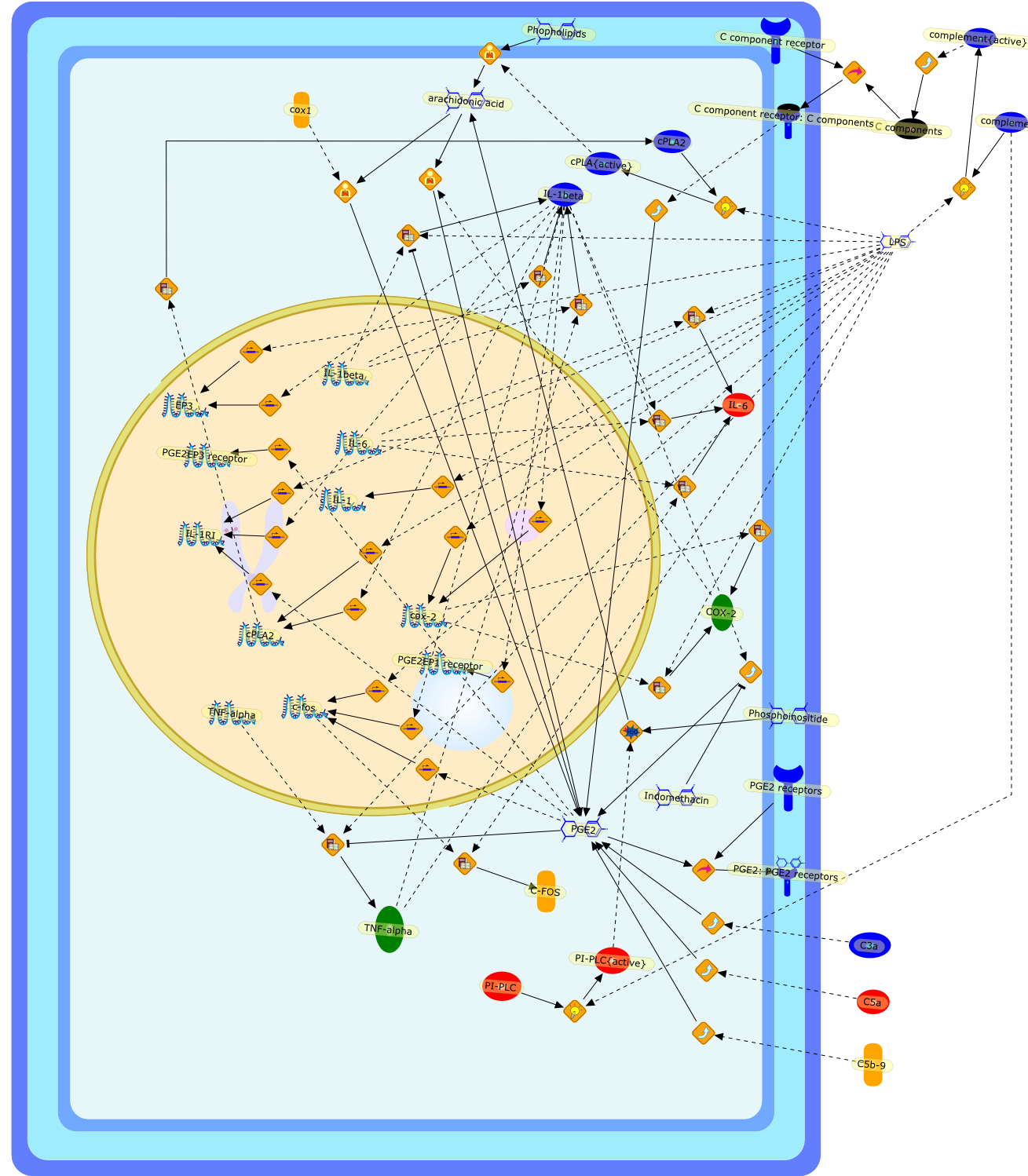
We and others recently showed that fever induced by intravenously orintraperitoneally injected lipopolysaccharide (LPS) may involve brain signalingvia hepatic vagal afferents. This suggests that LPS fever may be initiated bymediators released mainly by cells in the liver, presumably macrophages (Kupffercells, Kc). To verify this possibility, we disabled the Kc of conscious guineapigs with gadolinium chloride and monitored their core temperature andassociated preoptic prostaglandin E2 (PGE2) responses to i.v. LPS. Gadoliniumchloride pretreatment significantly attenuated both the febrile and PGE2 rises,thus supporting the hypothesis. Additionally, fluorescein-labeled LPS wasdetected in Kc 15 minutes after its i.v. administration. Paradoxically, however,the label was also present in gadolinium chloride-pretreated guinea pigs. Thus,either Kc are not the primary source of pyrogenic mediators or LPS does notprovide the stimulus for their production. Because the i.v. injection of LPSelicits virtually immediately the production of complement fragments, and Kcexpress their receptors and produce various mediators on their activation, wehypocomplemented guinea pigs with cobra venom factor. The core temperature risesproduced by i.v. LPS were reduced by complement depletions > 60%. LPS i.v. perse decreased complement, that is, complement was consumed by 12% within 10minutes. Thus, the onset of LPS fever may involve complement system and Kcactivation, but their precise roles await clarification.
{kind=link}
{kind=link}
C component receptor,C component receptor: C components,C components,C-FOS,C3a,C5a,C5b-9,COX-2,EP3,IL-1,IL-1RI,IL-1beta,IL-6,Indomethacin,LPS,PGE2,PGE2 receptors,PGE2: PGE2 receptors,PGE2EP1 receptor,PGE2EP3 receptor,PI-PLC,PI-PLC{active},Phopholipids,Phosphoinositide,TNF-alpha,arachidonic acid,c-fos,cPLA2,cPLA{active},complement,complement{active},cox-2,cox1
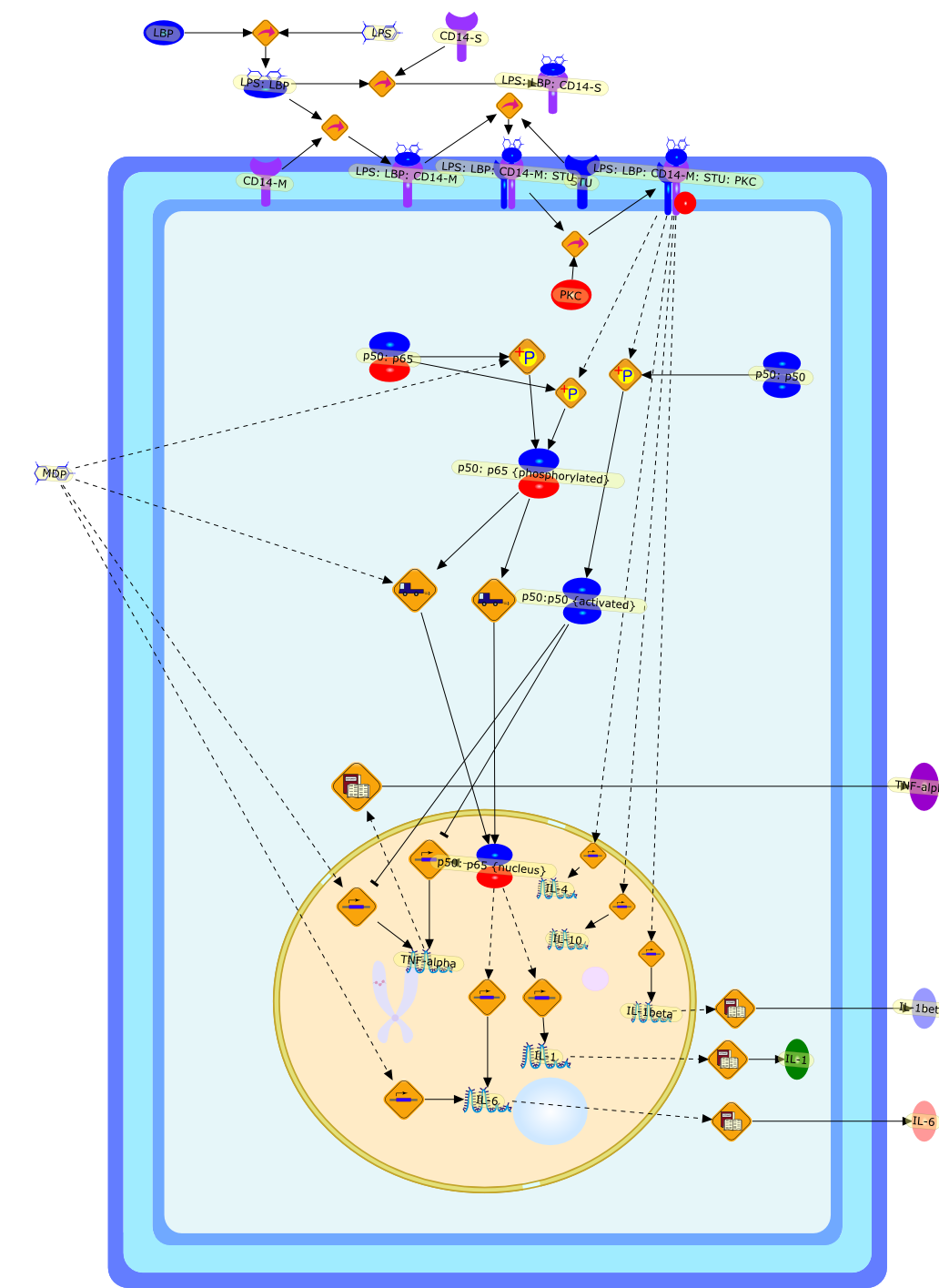
In humans or experimental animals, the repeated confrontation withlipopolysaccharides (LPS) from gram-negative bacteria, but not with muramyldipeptide (MDP) from gram-positive bacteria, leads to attenuation of almost allpathophysiologic effects mediated by proinflammatory cytokines. Our experimentsin guinea pigs and rats demonstrate that attenuation of the febrile responseduring the development of LPS tolerance is associated with a reduced productionof cytokines rather than a decrease in responsiveness to cytokines.Cross-tolerance experiments demonstrate that different stimuli influencingLPS-induced tumor necrosis factor (TNF) release and nitric oxide (NO) synthesiscan modify the development of tolerance. On the other hand, the lack ofcross-tolerance between LPS and MDP indicates that MDP can activate the cytokinecascade and induce the febrile response in animals tolerant to LPS. This mayindicate distinct receptors and signal pathways for LPS and MDP, leading toactivation of the cytokine cascade. LPS tolerance has also been demonstrated inex vivo and in vitro studies. In cultures of monocytes, diminished synthesis ofTNF and NO reported after LPS restimulation could be prevented and reversed byinterferon and granulocyte-macrophage colony-stimulating factor. These findingsadd an additional hypothesis in tolerance development.
{kind=link}
{kind=link}
CD14-M,CD14-S,IL-1,IL-10,IL-1beta,IL-4,IL-6,LBP,LPS,LPS: LBP,LPS: LBP: CD14-M,LPS: LBP: CD14-M: STU,LPS: LBP: CD14-M: STU: PKC,LPS: LBP: CD14-S,MDP,PKC,STU,TNF-alpha,p50: p50,p50: p65,p50: p65 {nucleus},p50: p65 {phosphorylated},p50:p50 {activated}
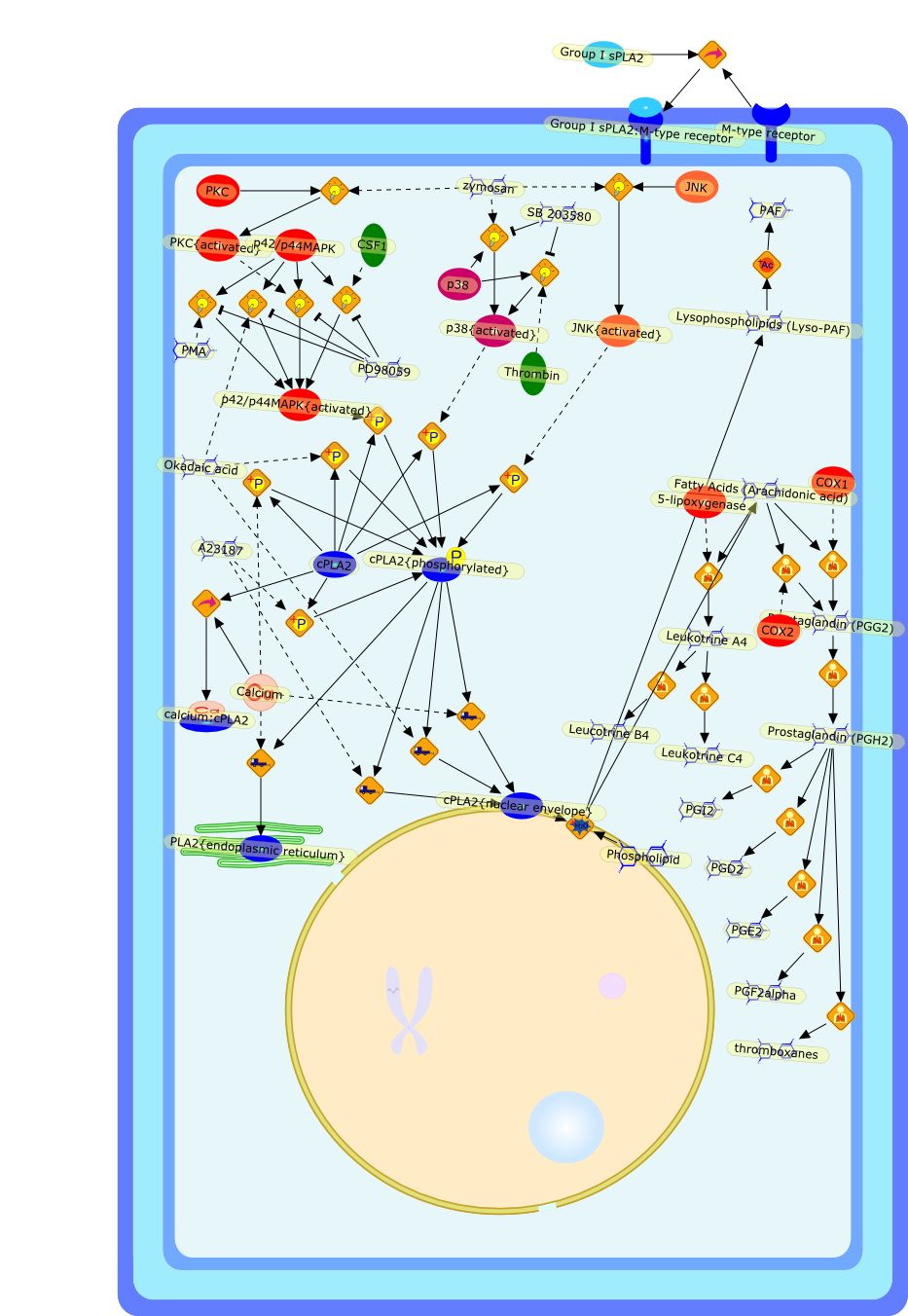
The 85-kDa cytosolic PLA2 (cPLA2) mediates agonist-induced arachidonic acidrelease in many cell models, including mouse peritoneal macrophages. cPLA2 isregulated by an increase in intracellular calcium, which binds to anamino-terminal C2 domain and induces its translocation to the nuclear envelopeand endoplasmic reticulum. Phosphorylation of cPLA2 on S505 by mitogen-activatedprotein kinases (MAPK) also contributes to activation. In macrophages, zymosaninduces a transient increase in intracellular calcium and activation of MAPK,which together fully activate cPLA2 and synergistically promote arachidonic acidrelease. There are alternative pathways for regulating cPLA2 in macrophagesbecause PMA and okadaic acid induce arachidonic acid release without increasingcalcium. The baculovirus expression system is a useful model to study cPLA2activation. Sf9 cells expressing cPLA2 release arachidonic acid to either A23187or okadaic acid. cPLA2 is phosphorylated on multiple sites in Sf9 cells, andphosphorylation of S727 is preferentially induced by okadaic acid. However, thephosphorylation sites are non-essential and only S505 phosphorylation partiallycontributes to cPLA2 activation in this model. Although okadaic acid does notincrease intracellular calcium in Sf9 cells, calcium binding by the C2 domain isnecessary for arachidonic acid release. A23187 and okadaic acid activate cPLA2by different mechanisms, yet both induce translocation to the nuclear envelopein Sf9 cells. The results demonstrate that alternative regulatory pathways canlead to cPLA2 activation and arachidonic acid release.
{kind=link}
{kind=link}
5-lipoxygenase,A23187,COX1,COX2,CSF1,Calcium,Fatty Acids (Arachidonic acid),Group I sPLA2,Group I sPLA2:M-type receptor,JNK,JNK{activated},Leucotrine B4,Leukotrine A4,Leukotrine C4,Lysophospholipids (Lyso-PAF),M-type receptor,Okadaic acid,PAF,PD98059,PGD2,PGE2,PGF2alpha,PGI2,PKC,PKC{activated},PLA2{endoplasmic reticulum},PMA,Phospholipid,Prostaglandin (PGG2),Prostaglandin (PGH2),SB 203580,Thrombin,cPLA2,cPLA2{nuclear envelope},cPLA2{phosphorylated},calcium:cPLA2,p38,p38{activated},p42/p44MAPK,p42/p44MAPK{activated},thromboxanes,zymosan
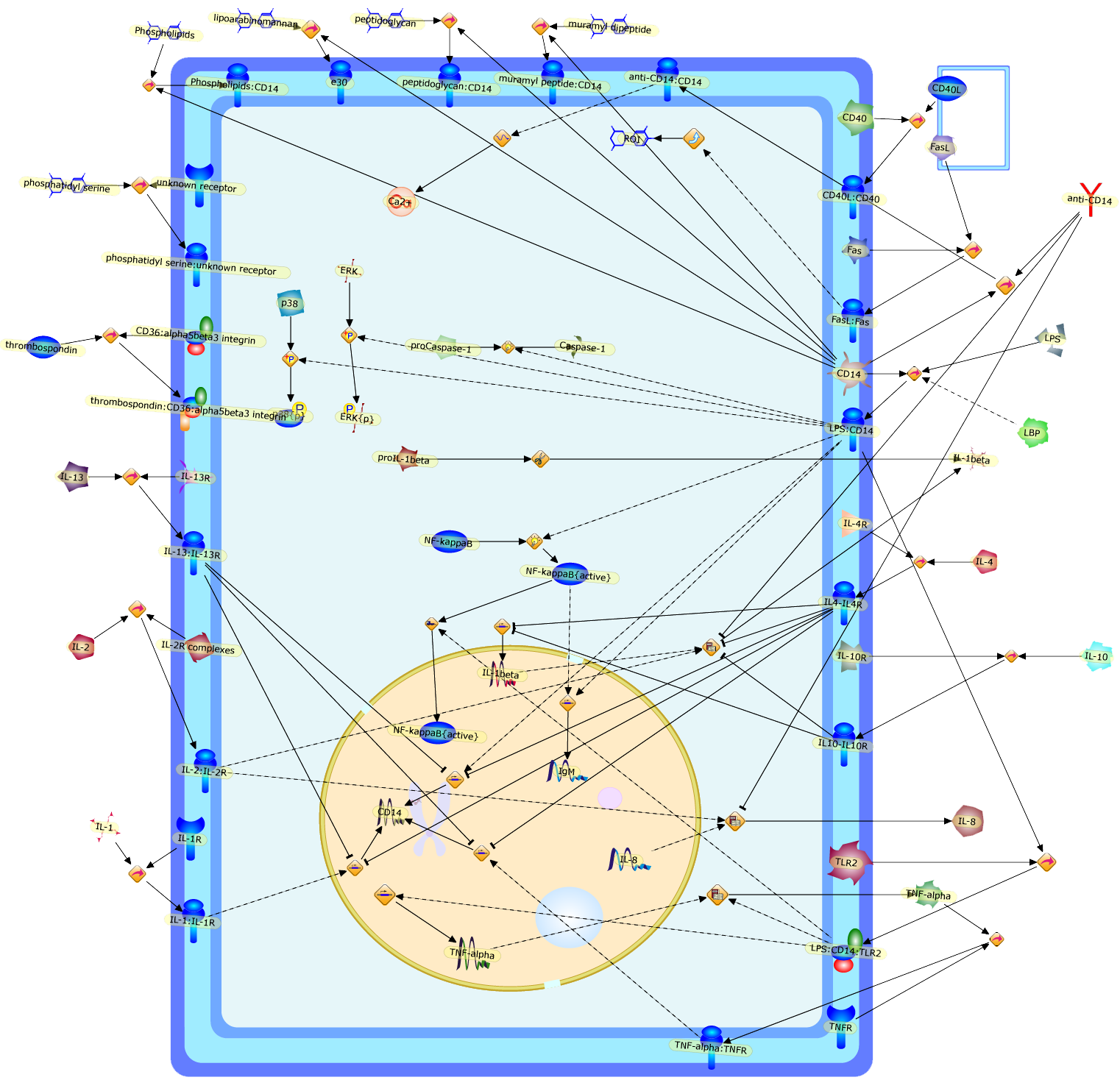
Like all other immune system cells, monocytes and macrophages may undergoapoptotic cell death in response to specific triggers and mediators or as aconsequence of aging. However, factors inducing apoptosis and the involvedcellular and molecular mechanisms are much better investigated and understoodfor lymphocytes. Th2 cell-derived cytokines such as interleukin-4 (IL-4) areable to induce monocyte apoptosis most effectively. This process is preceded bydown-regulation of the CD14 surface receptor. Mediators such aslipopolysaccharide (LPS) suppress and postpone apoptosis in parallel withup-regulation of CD14. Macrophages are rather resistant against apoptoticdamage, and factors able to evoke apoptosis in monocytes are often ineffectivein macrophages. Resistance of macrophages against apoptotic triggers may bebeneficial for inflammatory processes where macrophages are engaged and neededas phagocytes for ingestion and removal of moribund cells. The multifunctionalCD14 receptor of monocytes/macrophages is supposed to be involved in theapoptotic network on both sides: as a surface molecule of monocytes that canpromote survival and antagonize apoptosis and as a recognition receptor ofmacrophages that enables or supports interaction with apoptotic cells.
{kind=link}
{kind=link}
CD14,CD36:alpha5beta3 integrin,CD40,CD40L,CD40L:CD40,Ca2+,Caspase-1,ERK,ERK{p},Fas,FasL,FasL:Fas,IL-1,IL-10,IL-10R,IL-13,IL-13:IL-13R,IL-13R,IL-1:IL-1R,IL-1R,IL-1beta,IL-2,IL-2:IL-2R,IL-2R complexes,IL-4,IL-4R,IL-8,IL10-IL10R,IL4-IL4R,IgM,LBP,LPS,LPS:CD14,LPS:CD14:TLR2,NF-kappaB,NF-kappaB{active},Phospholipids,Phospholipids:CD14,ROI,TLR2,TNF-alpha,TNF-alpha:TNFR,TNFR,anti-CD14,anti-CD14:CD14,e30,lipoarabinomannan,muramyl dipeptide,muramyl peptide:CD14,p38,p38{p},peptidoglycan,peptidoglycan:CD14,phosphatidyl serine,phosphatidyl serine:unknown receptor,proCaspase-1,proIL-1beta,thrombospondin,thrombospondin:CD36:alpha5beta3 integrin,unknown receptor
{kind=link}
{kind=link}
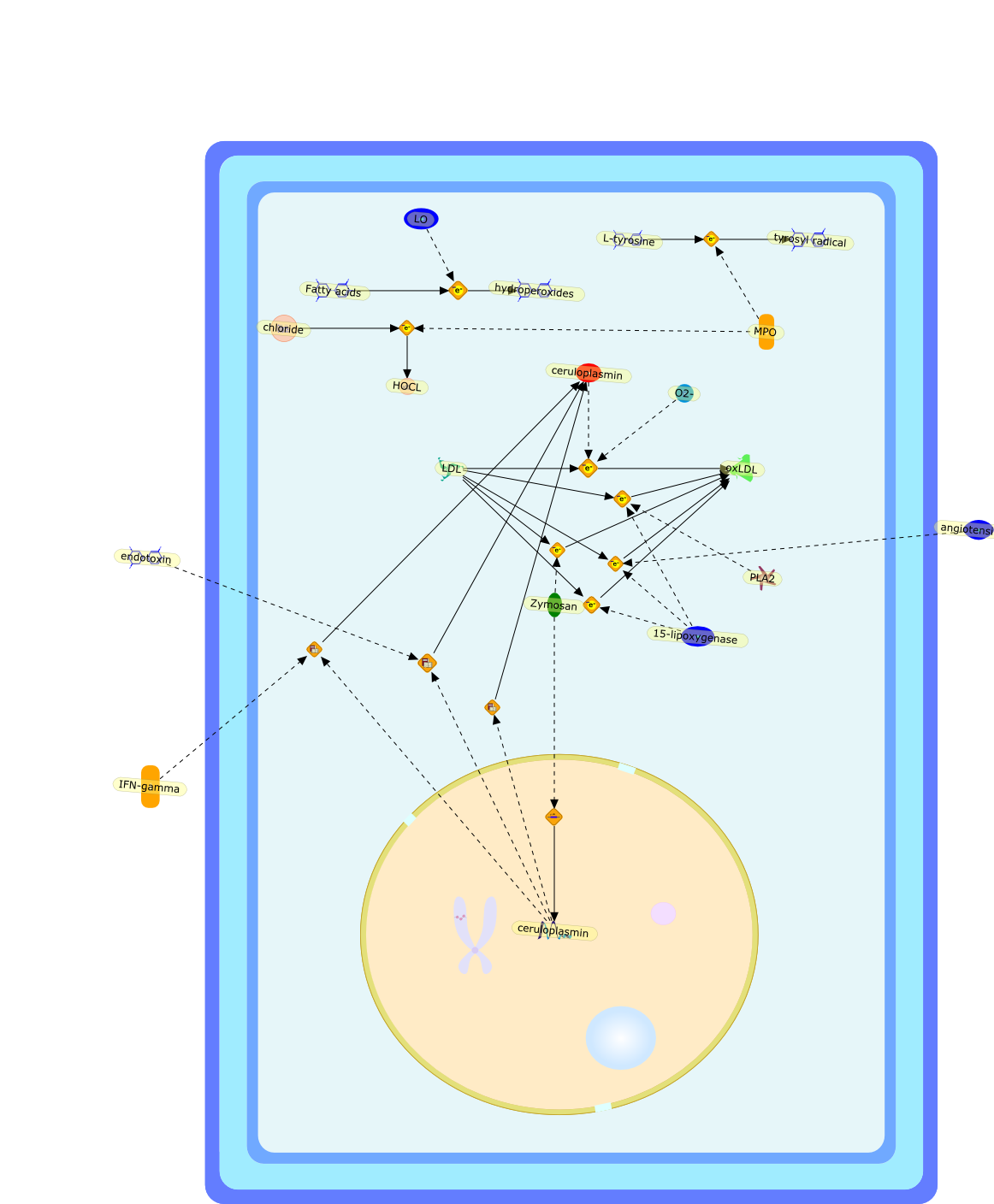
15-lipoxygenase,Fatty acids,HOCL,IFN-gamma,L-tyrosine,LDL,LO,MPO,O2-,PLA2,Zymosan,angiotensin-II,ceruloplasmin,chloride,endotoxin,hydroperoxides,oxLDL,tyrosyl radical
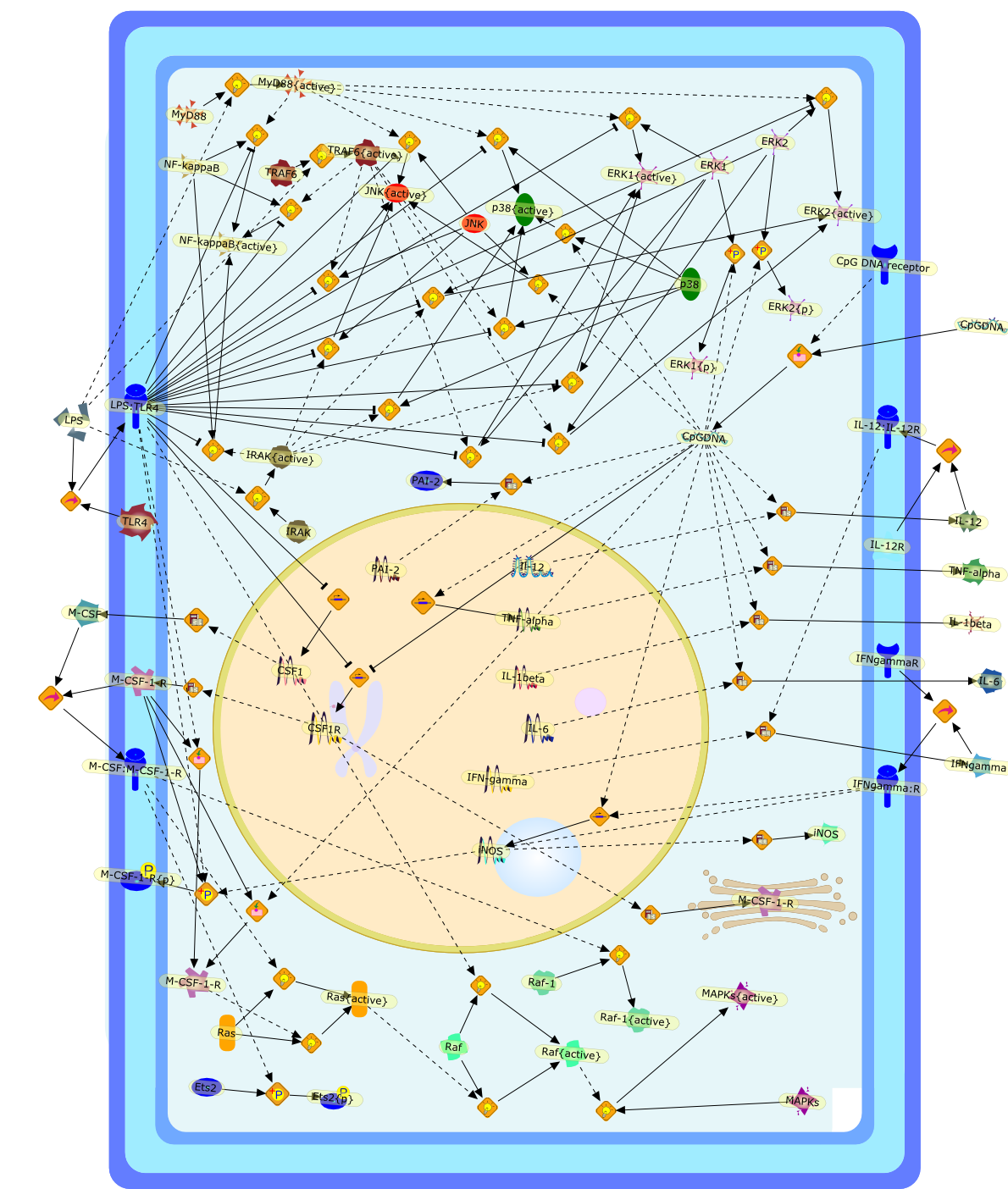
Murine macrophages are able to distinguish bacterial from mammalian DNA. Theresponse is mimicked by single-stranded oligonucleotides containing unmethylatedCG dinucleotides ("CpG" motifs) in specific sequence contexts. The dose-responsecurve for activation is influenced by variation in the sequence flanking thecore CpG motif. CpG or bacterial DNA activates several signaling pathways incommon with bacterial lipopolysaccharide (LPS), leading to induction of cytokinegenes such as tumor necrosis factor alpha. Pretreatment with LPS causesdesensitization to subsequent activation by CpG DNA. Both stimuli also causecell cycle arrest in macrophages proliferating in response to the macrophagegrowth factor colony-stimulating factor-1 (CSF-1), but prevent apoptosis causedby growth factor removal. In part, cell cycle arrest by CpG DNA and LPS may belinked to rapid down-modulation of the CSF-1 receptor from the cell surface, aresponse that occurs in an all-or-nothing manner. The response of macrophages toCpG DNA has aspects in common with the DNA damage response in other cell types,which may provide clues to the underlying mechanism.
{kind=link}
{kind=link}
CSF1,CSF1R,CpG DNA receptor,CpGDNA,ERK1,ERK1{active},ERK1{p},ERK2,ERK2{active},ERK2{p},Ets2,Ets2{p},IFN-gamma,IFNgamma,IFNgamma:R,IFNgammaR,IL-12,IL-12:IL-12R,IL-12R,IL-1beta,IL-6,IRAK,IRAK{active},Il-12,JNK,JNK{active},LPS,LPS:TLR4,M-CSF,M-CSF-1-R,M-CSF-1-R{p},M-CSF:M-CSF-1-R,MAPKs,MAPKs{active},MyD88,MyD88{active},NF-kappaB,NF-kappaB{active},PAI-2,Raf,Raf-1,Raf-1{active},Raf{active},Ras,Ras{active},TLR4,TNF-alpha,TRAF6,TRAF6{active},iNOS,p38,p38{active}
The primary interleukin-4 (IL-4) receptor complex on monocytes (type I IL-4receptor) includes the 140-kDa alpha chain (IL-4R alpha) and the IL-2 receptorgamma chain, gamma(c), which heterodimerize for intracellular signaling,resulting in suppression of lipopolysaccharide (LPS)-inducible inflammatorymediator production. The activity of IL-13 on human monocytes is very similar tothat of IL-4 because the predominant signaling chain (IL-4R alpha) is common toboth receptors. In fact, IL-4R alpha with IL-13R alpha1 is designated both as anIL-13 receptor and the type II IL-4 receptor. When the anti-inflammatoryactivities of IL-4 and IL-13 were investigated on synovial fluid macrophages andcompared with the responses by monocytes isolated from the patients at the sametime as joint drainage, the response profiles differed with some responsessimilar in the two cell populations, others reduced on the inflammatory cells.Similar differences were recorded in the response profiles to IL-4 and IL-13 bymonocytes and monocytes cultured for 7 days in macrophage colony-stimulatingfactor (M-CSF) or granulocyte-macrophage CSF (GM-CSF) (monocyte-derivedmacrophages, MDMac). MDMac have reduced gamma(c) mRNA levels and reducedexpression of the functional 64-kDa gamma(c). There was a similar loss of IL-13Ralpha1 mRNA on monocyte differentiation. In turn, there was a significantreduction in the ability of IL-4 and IL-13 to activate STAT6. These findingssuggest that different functional responses to IL-4 and IL-13 by human monocytesand macrophages may result from reduced expression of gamma(c) and IL-13Ralpha1.
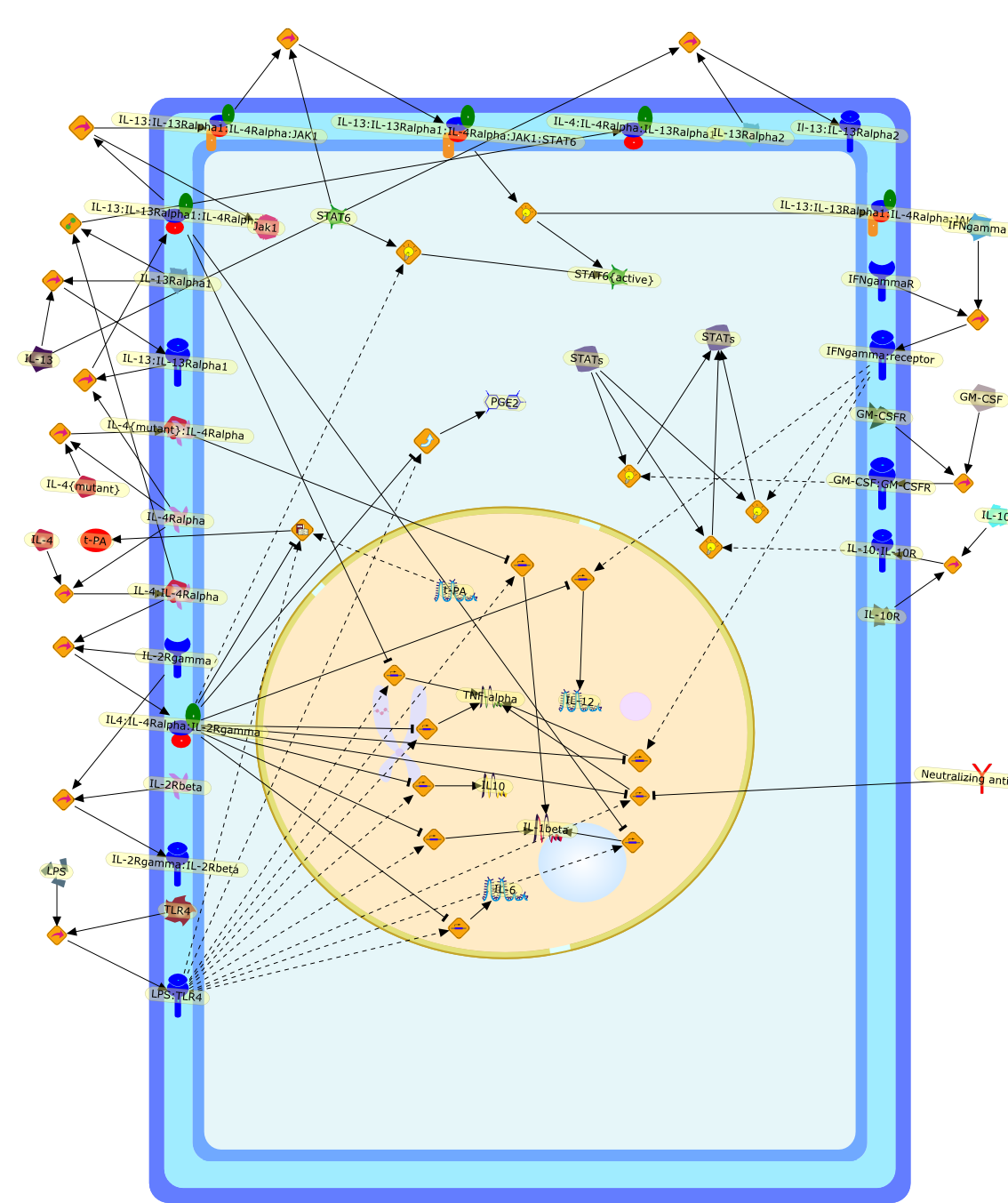{kind=link}
{kind=link}
GM-CSF,GM-CSF:GM-CSFR,GM-CSFR,IFNgamma,IFNgamma:receptor,IFNgammaR,IL-10,IL-10:IL-10R,IL-10R,IL-12,IL-13,IL-13:IL-13Ralpha1,IL-13:IL-13Ralpha1:IL-4Ralpha,IL-13:IL-13Ralpha1:IL-4Ralpha:JAK1,IL-13:IL-13Ralpha1:IL-4Ralpha:JAK1:STAT6,IL-13Ralpha1,IL-13Ralpha2,IL-1beta,IL-2Rbeta,IL-2Rgamma,IL-2Rgamma:IL-2Rbeta,IL-4,IL-4:IL-4Ralpha,IL-4:IL-4Ralpha:IL-13Ralpha1,IL-4Ralpha,IL-4{mutant},IL-4{mutant}:IL-4Ralpha,IL-6,IL10,IL4:IL-4Ralpha:IL-2Rgamma,Il-13:IL-13Ralpha2,Jak1,LPS,LPS:TLR4,Neutralizing antibiody,PGE2,STAT6,STAT6{active},STATs,TLR4,TNF-alpha,t-PA
In the past three years, a novel signal transduction pathway downstream of thetransforming growth factor-beta (TGF-beta) superfamily receptor serine-threoninekinases has been shown to be mediated by a family of latent transcriptionfactors called 'Smads'. These proteins mediate a short-circuited pathway inwhich a set of receptor-activated Smads are phosphorylated directly by thereceptor kinase and then translocate to the nucleus complexed to the commonmediator, Smad4, to participate in transcriptional complexes. Smads 2 and 3mediate signals predominantly from the TGF-beta receptors. Of these, specificroles have been ascribed to Smad3 in control of chemotaxis of neutrophils andmacrophages and the inhibition of Smad3 activity by the oncogene Evi-1 suggeststhat it may play a role in leukemogenesis. Other data, such as the induction bythe inflammatory cytokine interferon-gamma of an inhibitory Smad, Smad7, whichblocks the actions of Smad3, suggest that identification of the specific genetargets of Smad proteins in immune cells will provide new insight into themechanisms of TGF-beta action on these cells.
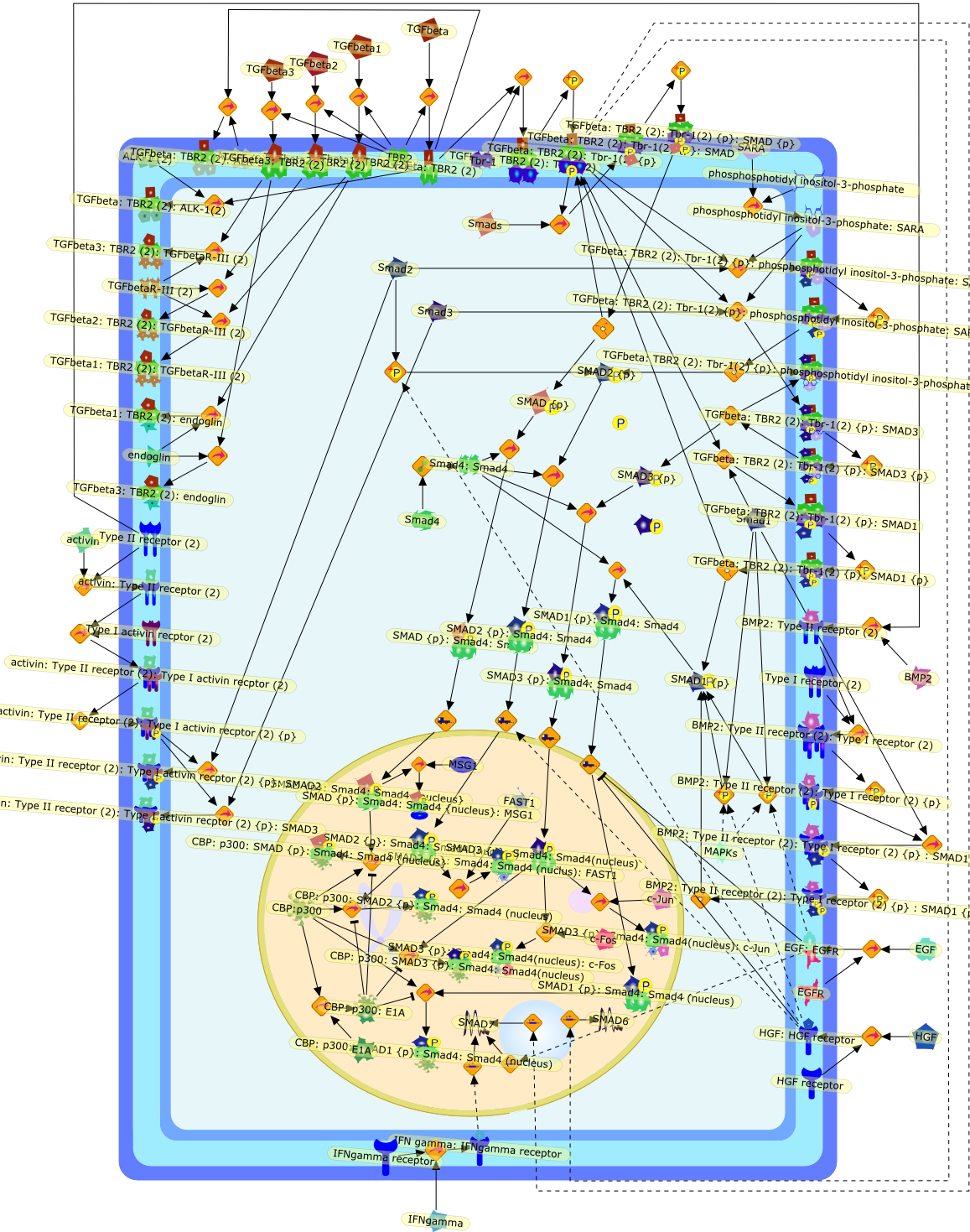{kind=link}
{kind=link}
ALK-1 (2),ALK-5 (2),BMP2,BMP2: Type II receptor (2),BMP2: Type II receptor (2): Type I receptor (2),BMP2: Type II receptor (2): Type I receptor (2) {p},BMP2: Type II receptor (2): Type I receptor (2) {p} : SMAD1,BMP2: Type II receptor (2): Type I receptor (2) {p} : SMAD1 {p},CBP: p300: E1A,CBP: p300: SMAD {p}: Smad4: Smad4 {nucleus},CBP: p300: SMAD1 {p}: Smad4: Smad4 (nucleus),CBP: p300: SMAD2 {p}: Smad4: Smad4 (nucleus),CBP: p300: SMAD3 {p}: Smad4: Smad4(nucleus),CBP:p300,E1A,EGF,EGF: EGFR,EGFR,FAST1,HGF,HGF receptor,HGF: HGF receptor,IFN gamma: IFNgamma receptor,IFNgamma,IFNgamma receptor,MAPKs,MSG1,SARA,SMAD {p},SMAD {p}: Smad4: Smad4,SMAD {p}: Smad4: Smad4 {nucleus},SMAD {p}: Smad4: Smad4 {nucleus}: MSG1,SMAD1 {p},SMAD1 {p}: Smad4: Smad4,SMAD1 {p}: Smad4: Smad4 (nucleus),SMAD2 {p},SMAD2 {p}: Smad4: Smad4,SMAD2 {p}: Smad4: Smad4 (nucleus),SMAD2 {p}: Smad4: Smad4 (nuclus): FAST1,SMAD3 {p},SMAD3 {p}: Smad4: Smad4,SMAD3 {p}: Smad4: Smad4(nucleus),SMAD3 {p}: Smad4: Smad4(nucleus): c-Fos,SMAD3 {p}: Smad4: Smad4(nucleus): c-Jun,SMAD6,SMAD7,Smad1,Smad2,Smad3,Smad4,Smad4: Smad4,Smads,TBR2,TGFbeta,TGFbeta1,TGFbeta1: TBR2 (2),TGFbeta1: TBR2 (2): TGFbetaR-III (2),TGFbeta1: TBR2 (2): endoglin,TGFbeta2,TGFbeta2: TBR2 (2),TGFbeta2: TBR2 (2): TGFbetaR-III (2),TGFbeta3,TGFbeta3: TBR2 (2),TGFbeta3: TBR2 (2): TGFbetaR-III (2),TGFbeta3: TBR2 (2): endoglin,TGFbeta: TBR2 (2),TGFbeta: TBR2 (2): ALK-1(2),TGFbeta: TBR2 (2): ALK-5(2),TGFbeta: TBR2 (2): Tbr-1(2),TGFbeta: TBR2 (2): Tbr-1(2) {p},TGFbeta: TBR2 (2): Tbr-1(2) {p}: SMAD,TGFbeta: TBR2 (2): Tbr-1(2) {p}: SMAD {p},TGFbeta: TBR2 (2): Tbr-1(2) {p}: SMAD1,TGFbeta: TBR2 (2): Tbr-1(2) {p}: SMAD1 {p},TGFbeta: TBR2 (2): Tbr-1(2) {p}: SMAD3,TGFbeta: TBR2 (2): Tbr-1(2) {p}: SMAD3 {p},TGFbeta: TBR2 (2): Tbr-1(2) {p}: phosphosphotidyl inositol-3-phosphate: SARA,TGFbeta: TBR2 (2): Tbr-1(2) {p}: phosphosphotidyl inositol-3-phosphate: SARA: SMAD2,TGFbeta: TBR2 (2): Tbr-1(2) {p}: phosphosphotidyl inositol-3-phosphate: SARA: SMAD2 {p},TGFbetaR-III (2),Tbr-1,Type I activin recptor (2),Type I receptor (2),Type II receptor (2),activin,activin: Type II receptor (2),activin: Type II receptor (2): Type I activin recptor (2),activin: Type II receptor (2): Type I activin recptor (2) {p},activin: Type II receptor (2): Type I activin recptor (2) {p}: SMAD2,activin: Type II receptor (2): Type I activin recptor (2) {p}: SMAD3,c-Fos,c-Jun,endoglin,phosphosphotidyl inositol-3-phosphate,phosphosphotidyl inositol-3-phosphate: SARA
Lipopolysaccharide (LPS), an abundant glycolipid of the outer membrane ofgram-negative bacteria, is able to provoke a generalized proinflammatoryresponse in the infected host. Genetic regulation of this trait has beenlocalized to the Lps locus on mouse chromosome 4. Several inbred mouse strains,including C3H/HeJ, C57BL/10ScNCr and C57BL/10ScCr, bear mutations at the Lpslocus (Lps(d)) that confer hyporesponsiveness to the immunostimulatoryproperties of LPS and susceptibility to overwhelming gram-negative bacterialinfection. The phenotypic expression of Lps(d) is pleiotropic, affecting severalcell types crucial to host defense, including the macrophage. By positionalcloning, Toll-like receptor 4 (Tlr4), a transmembrane protein with a cytoplasmicdomain that bears homology to the Interleukin-1 receptor, has been identified asthe gene encoded by Lps. Tlr4 is a member of a novel gene family thatparticipates in host defense against microbial infection in plants,invertebrates and mammals. Discovery of the molecular basis of the Lps mutationrepresents a significant advance in defining the fundamental mechanisms ofcellular activation by LPS.
{kind=link}
{kind=link}
B7.1,Dorsal,IKB,IL-1,IL-1R,IL-1R:IL-1,IL-6,IL-8,LBP,LPS,LPS:LBP,LPS:LBP:CD14:TLR4,LPS:LBP:CD14:TLR4:MYD88,LPS:LBP:mCD14,LPS:LBP:sCD14,MYD88,NFKB:IKB,NFKB[active},NFKB{active},SIIK,SIIK{active},TLR4,TOLL,TOLL:Spatzle,TRAF6,TRAF6{active},mCD14,mRNA,sCD14,spatzle
Innate immune response to microbes sometimes determines the nature of thefollowing specific immune response. Kupffer cells, a potent constituent ofinnate immunity, play a key role in developing the type 1 immune response byinterleukin (IL)-12 production. Furthermore, Kupffer cells have the potential toinduce liver injury by production of IL-18. Propionibacterium acnes-primedlipopolysaccharide (LPS)-challenged liver injury is the prototype ofIL-18-induced tissue injury, in which IL-18 acts on natural killer cells toincrease Fas ligand (FasL) that causes liver injury by induction ofFas-dependent hepatocyte apoptosis. LPS induces IL-18 secretion from Kupffercells in a caspase-1-dependent manner. Indeed, caspase-1-deficient mice areresistant to P. acnes and LPS-induced liver injury. However, administration ofsoluble FasL induces acute liver injury in P. acnes-primed caspase-1-deficientmice but does not do so in IL-18-deficient mice, indicating that IL-18 releasein a caspase-1-independent fashion is essential for this liver injury.Therefore, a positive feedback loop between FasL and IL-18 plays an importantrole in the pathogenesis of endotoxin-induced liver injury.
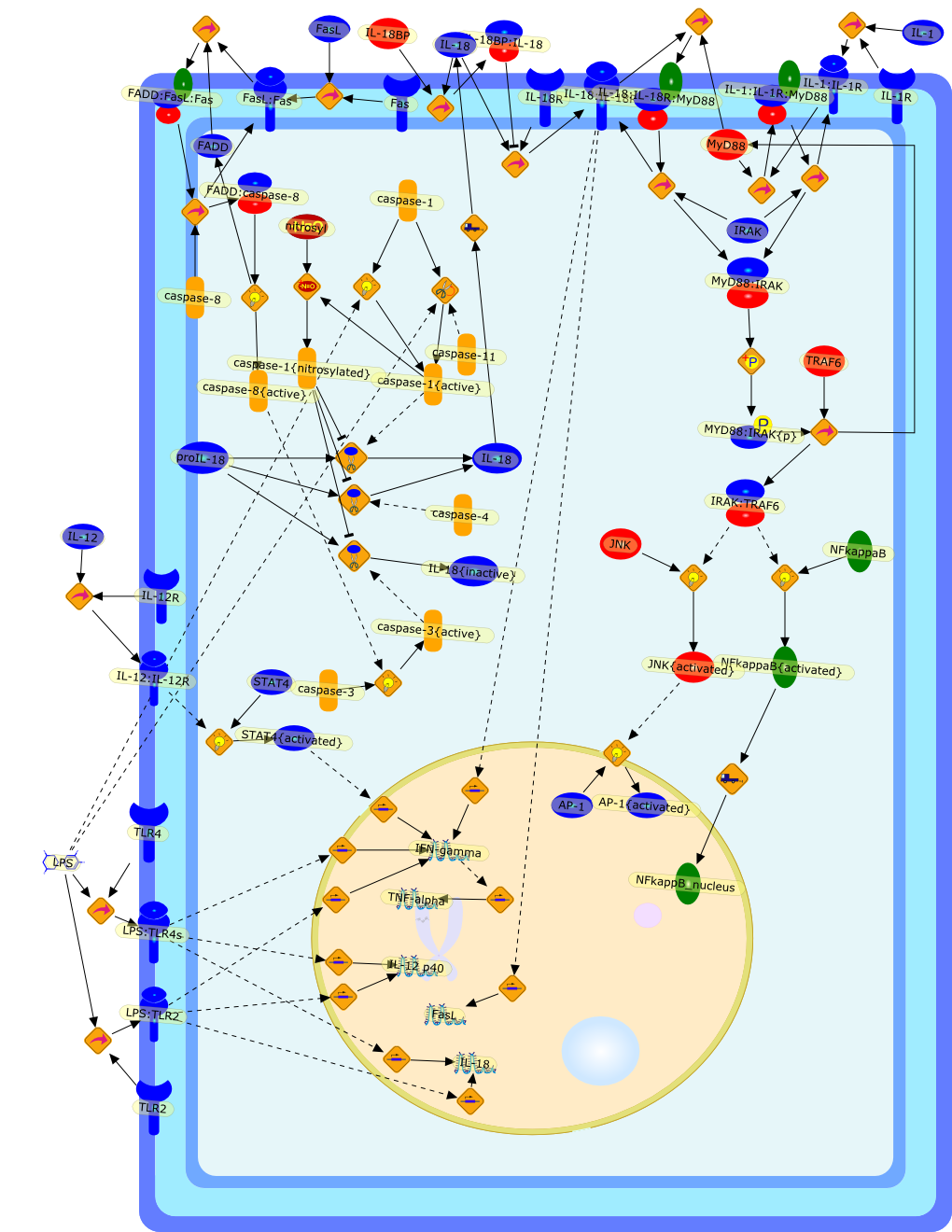{kind=link}
{kind=link}
AP-1,AP-1{activated},FADD,FADD:FasL:Fas,FADD:caspase-8,Fas,FasL,FasL:Fas,IFN-gamma,IL-1,IL-12,IL-12 p40,IL-12:IL-12R,IL-12R,IL-18,IL-18:IL-18R,IL-18:IL-18R:MyD88,IL-18BP,IL-18BP:IL-18,IL-18R,IL-18{inactive},IL-1:IL-1R,IL-1:IL-1R:MyD88,IL-1R,IRAK,IRAK:TRAF6,JNK,JNK{activated},LPS,LPS:TLR2,LPS:TLR4s,MYD88:IRAK{p},MyD88,MyD88:IRAK,NFkappB_nucleus,NFkappaB,NFkappaB{activated},STAT4,STAT4{activated},TLR2,TLR4,TNF-alpha,TRAF6,caspase-1,caspase-11,caspase-1{active},caspase-1{nitrosylated},caspase-3,caspase-3{active},caspase-4,caspase-8,caspase-8{active},nitrosyl,proIL-18
Whereas most cells in adult tissues are fixed in place by cell junctions,leukocytes are motile and able to migrate actively through the walls of bloodvessels into surrounding tissues. The actin cytoskeleton of these cells plays acentral role in locomotion, phagocytosis, and the regulation of cell shape thatare crucial elements of neutrophil and monocyte/macrophage function. This reviewwill concentrate on how macrophages in particular control the actin cytoskeletonto generate cell movement and the shape changes required for chemotaxis. It hasrecently become evident that a complex of seven proteins known as the Arp2/3complex regulates the assembly of new actin filament networks at the leadingfront of moving cells. Proteins of the Wiskott-Aldrich Syndrome Protein (WASP)family bind directly to the Arp2/3 complex and stimulate its ability to promotethe nucleation of new actin filaments. Upstream of the WASP family proteins,receptor tyrosine kinases, G-protein-coupled receptors, phosphoinositide-3-OHkinase (PI 3-kinase), and the Rho family of GTPases receive and transduce thesignals that lead to actin nucleation through WASP-Arp2/3 action. Although manygaps remain in our understanding, we are now in a position to considercompleting signaling pathways that are initiated from outside the cell to theactin rearrangements that drive cell motility and chemotaxis.
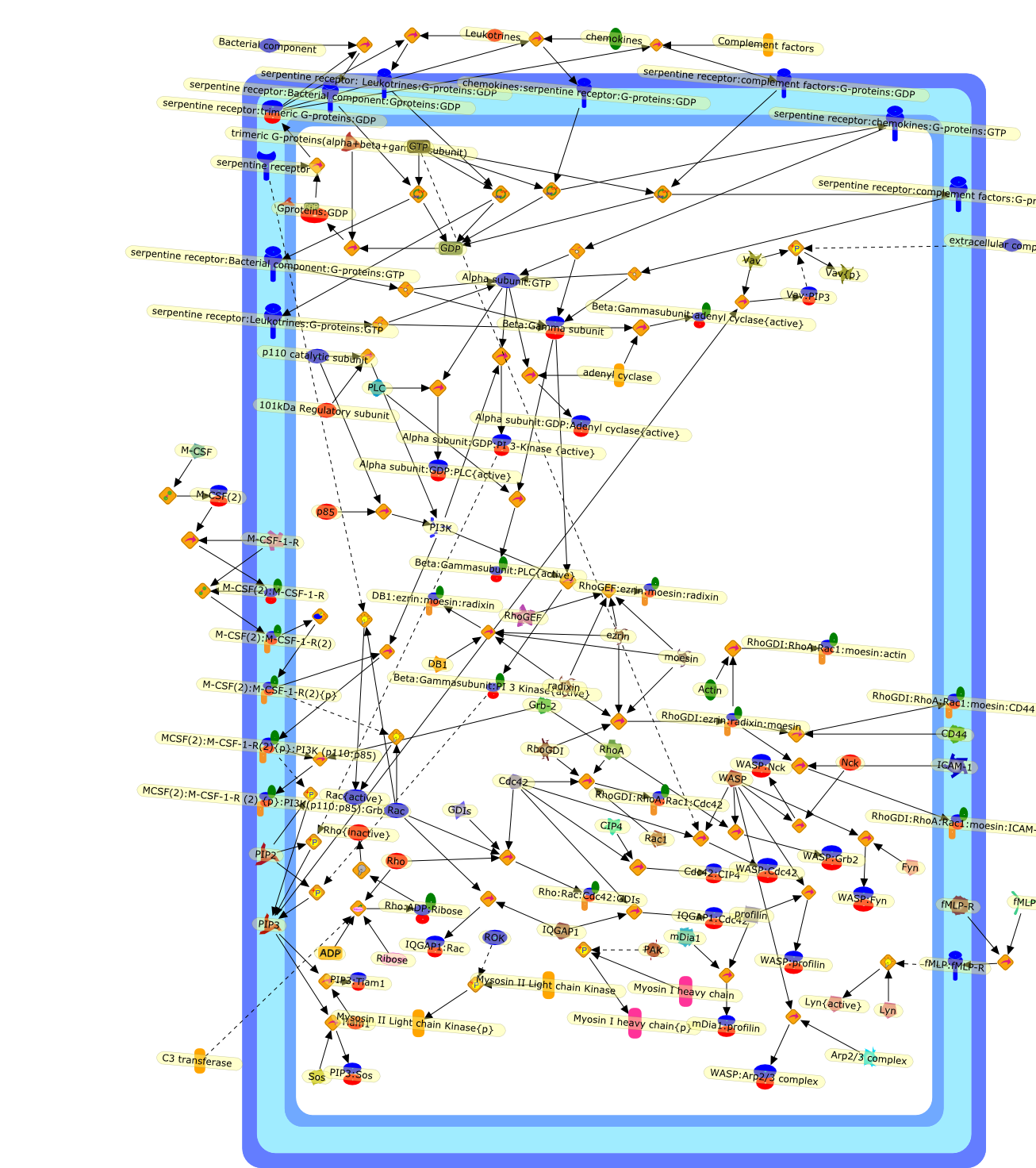{kind=link}
{kind=link}
101kDa Regulatory subunit,ADP,Actin,Alpha subunit:GDP:Adenyl cyclase{active},Alpha subunit:GDP:PI 3-Kinase {active},Alpha subunit:GDP:PLC{active},Alpha subunit:GTP,Arp2/3 complex,Bacterial component,Beta:Gamma subunit,Beta:Gammasubunit:PI 3 Kinase{active},Beta:Gammasubunit:PLC{active},Beta:Gammasubunit:adenyl cyclase{active},C3 transferase,CD44,CIP4,Cdc42,Cdc42:CIP4,Complement factors,DB1,DB1:ezrin:moesin:radixin,Fyn,GDIs,GDP,GTP,Gproteins:GDP,Grb-2,ICAM-1,IQGAP1,IQGAP1:Cdc42,IQGAP1:Rac,Leukotrines,Lyn,Lyn{active},M-CSF,M-CSF(2),M-CSF(2):M-CSF-1-R,M-CSF(2):M-CSF-1-R(2),M-CSF(2):M-CSF-1-R(2){p},M-CSF-1-R,MCSF(2):M-CSF-1-R (2) {p}:PI3K(p110:p85):Grb2,MCSF(2):M-CSF-1-R(2){p}:PI3K (p110:p85),Myosin I heavy chain,Myosin I heavy chain{p},Mysosin II Light chain Kinase,Mysosin II Light chain Kinase{p},Nck,PAK,PI3K,PIP2,PIP3,PIP3:Sos,PIP3:Tiam1,PLC,ROK,Rac,Rac1,Rac{active},Rho,Rho:ADP:Ribose,Rho:Rac:Cdc42:GDIs,RhoA,RhoGDI,RhoGDI:RhoA:Rac1:Cdc42,RhoGDI:RhoA:Rac1:moesin:CD44,RhoGDI:RhoA:Rac1:moesin:ICAM-1,RhoGDI:RhoA:Rac1:moesin:actin,RhoGDI:ezrin:radixin:moesin,RhoGEF,RhoGEF:ezrin:moesin:radixin,Rho{inactive},Ribose,Sos,Tiam1,Vav,Vav:PIP3,Vav{p},WASP,WASP:Arp2/3 complex,WASP:Cdc42,WASP:Fyn,WASP:Grb2,WASP:Nck,WASP:profilin,adenyl cyclase,chemokines,chemokines:serpentine receptor:G-proteins:GDP,extracellular components,ezrin,fMLP,fMLP-R,fMLP:fMLP-R,mDia1,mDia1:profilin,moesin,p110 catalytic subunit,p85,profilin,radixin,serpentine receptor,serpentine receptor: Leukotrines:G-proteins:GDP,serpentine receptor:Bacterial component:G-proteins:GTP,serpentine receptor:Bacterial component:Gproteins:GDP,serpentine receptor:Leukotrines:G-proteins:GTP,serpentine receptor:chemokines:G-proteins:GTP,serpentine receptor:complement factors:G-proteins:GDP,serpentine receptor:complement factors:G-proteins:GTP,serpentine receptor:trimeric G-proteins:GDP,trimeric G-proteins(alpha+beta+gamma subunit)
Lipopolysaccharide (LPS), also known as endotoxin, is a compound of the cellwall of Gram-negative bacteria, which has been demonstrated to induceinflammatory reactions in vitro as well as in vivo, including lethal shock. Agreat number of different cells have been documented to be reactive to LPS, e.g.monocytes/macrophages, vascular cells, polymorphonuclear cells, and even Blymphocytes. We have now established that T lymphocytes could also contribute toan inflammatory reaction to LPS. LPS is a potent inducer of human T-lymphocyteproliferation and cytokine production. The activation of T lymphocytes by LPSrequires direct cell-to-cell contact with viable accessory monocytes. Thisinteraction was found to be MHC-unrestricted, but strongly dependent oncostimulatory signals provided by B7/CD28 interactions. The frequency ofresponding T lymphocytes is less than 1:1000. A very exciting finding was thatnot only monocytes, but also CD34+ hematopoietic stem cells, which circulate inperipheral blood in very low frequency, exert essential accessory cell activityduring stimulation of T lymphocytes by LPS. In contrast, the response of Tlymphocytes to conventional recall antigens is not controlled by blood stemcells. These conclusions are based on the observation that depletion ofCD34-positive blood stem cells resulted in a complete loss of LPS-inducedT-lymphocyte stimulation. Addition of CD34-enriched blood stem cells led to arecovery of reactivity of T lymphocyte to LPS. The characteristics ofT-lymphocyte activation indicate that LPS is neither active as a mitogen, or asa superantigen, or as a classical antigen, but may activate T lymphocyte througha new, so far undescribed, mechanism. Furthermore, the involvement ofhematopoietic blood stem cells in the activation of T lymphocytes by LPSdemonstrates a role of these cells in inflammatory and immunological events.
{kind=link}
{kind=link}
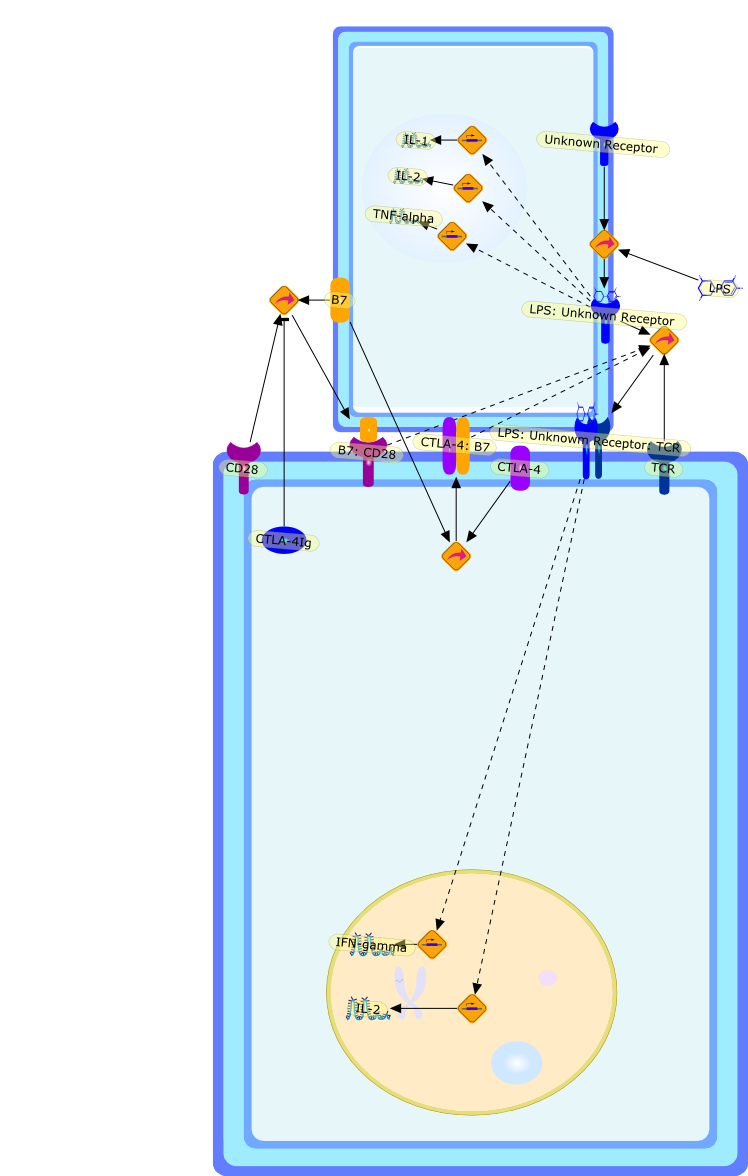
B7,B7: CD28,CD28,CTLA-4,CTLA-4: B7,CTLA-4Ig,IFN-gamma,IL-1,IL-2,LPS,LPS: Unknowm Receptor: TCR,LPS: Unknown Receptor,TCR,TNF-alpha,Unknown Receptor
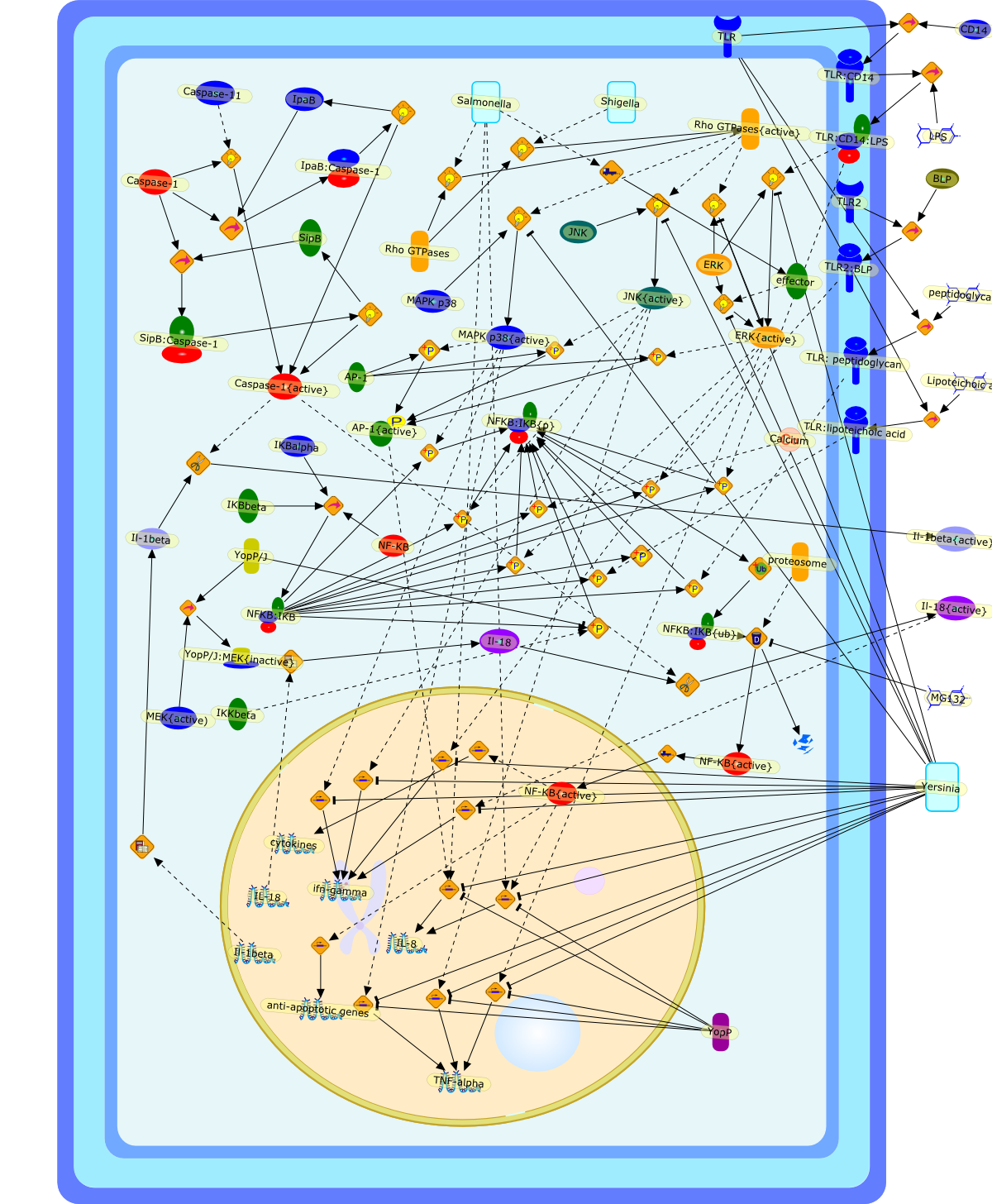
Microbe-macrophage interactions play a central role in the pathogenesis of manyinfections. Several bacterial pathogens induce apoptosis specifically inmacrophages, but the mechanisms by which it occurs differ, and the resultingpathology can take different courses. Macrophage death caused by Shigellaflexneri and Salmonella spp. has been shown to result in the release ofpro-inflammatory cytokines. Conversely, Yersinia spp. induce apoptosis bysuppressing the signalling pathways that lead to the production of tumournecrosis factor (TNF)-alpha, a cytokine essential for the control of thisinfection. It is likely that there are a variety of reasons why macrophages areparticularly susceptible to pathogen-induced apoptosis. One reason may be theexpression of surface receptors that recognize highly conserved bacterialcomponents, such as lipopolysaccharide (LPS) and bacterial lipoproteins (BLPs).These receptors have recently been shown to activate pro-apoptotic signallingpathways. The roles of macrophage apoptosis in different disease processes arediscussed.
{kind=link}
{kind=link}
AP-1,AP-1{active},BLP,CD14,Calcium,Caspase-1,Caspase-11,Caspase-1{active},ERK,ERK{active},IKBalpha,IKBbeta,IKKbeta,IL-18,IL-8,Il-18,Il-18{active},Il-1beta,Il-1beta{active},IpaB,IpaB:Caspase-1,JNK,JNK{active},LPS,Lipoteichoic acids,MAPK p38,MAPK p38{active},MEK{active),MG132,NF-KB,NF-KB{active},NFKB:IKB,NFKB:IKB{p},NFKB:IKB{ub},Rho GTPases,Rho GTPases{active},Salmonella,Shigella,SipB,SipB:Caspase-1,TLR,TLR2,TLR2:BLP,TLR: peptidoglycan,TLR:CD14,TLR:CD14:LPS,TLR:lipoteichoic acid,TNF-alpha,Yersinia,YopP,YopP/J,YopP/J:MEK{inactive},anti-apoptotic genes,cytokines,effector,ifn-gamma,peptidoglycan,protein remnants,proteosome
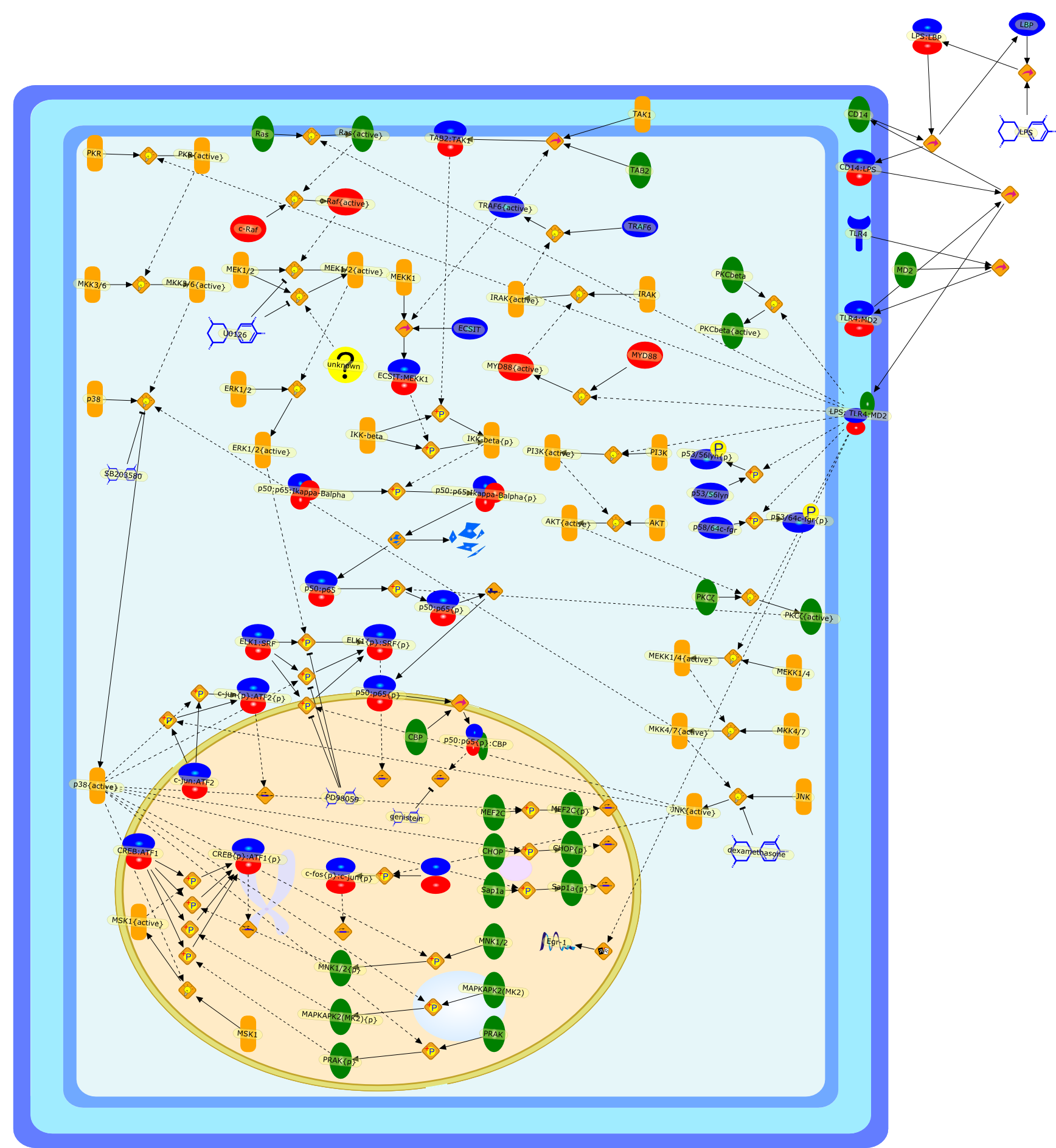
Lipopolysaccharide (LPS [endotoxin]) is the principal component of the outermembrane of Gram-negative bacteria. Recent studies have elucidated how LPS isrecognized by monocytes and macrophages of the innate immune system. Humanmonocytes are exquisitely sensitive to LPS and respond by expressing manyinflammatory cytokines. LPS binds to LPS-binding protein (LBP) in plasma and isdelivered to the cell surface receptor CD14. Next, LPS is transferred to thetransmembrane signaling receptor toll-like receptor 4 (TLR4) and its accessoryprotein MD2. LPS stimulation of human monocytes activates several intracellularsignaling pathways that include the IkappaB kinase (IKK)-NF-kappaB pathway andthree mitogen-activated protein kinase (MAPK) pathways: extracellularsignal-regulated kinases (ERK) 1 and 2, c-Jun N-terminal kinase (JNK) and p38.These signaling pathways in turn activate a variety of transcription factorsthat include NF-kappaB (p50/p65) and AP-1 (c-Fos/c-Jun), which coordinate theinduction of many genes encoding inflammatory mediators.
{kind=link}
{kind=link}
AKT,AKT{active},CBP,CD14,CD14:LPS,CHOP,CHOP{p},CREB:ATF1,CREB{p}:ATF1{p},ECSIT,ECSIT:MEKK1,ELK1:SRF,ELK1{p}:SRF{p},ERK1/2,ERK1/2{active},Egr-1,IKK-beta,IKK-beta{p},IRAK,IRAK{active},JNK,JNK{active},LBP,LPS,LPS: TLR4:MD2,LPS:LBP,MAPKAPK2(MK2),MAPKAPK2(MK2){p},MD2,MEF2C,MEF2C{p},MEK1/2,MEK1/2{active},MEKK1,MEKK1/4,MEKK1/4{active},MKK3/6,MKK3/6{active},MKK4/7,MKK4/7{active},MNK1/2,MNK1/2{p},MSK1,MSK1{active},MYD88,MYD88{active},PD98059,PI3K,PI3K{active},PKCbeta,PKCbeta{active},PKC�Ύ�,PKC�Ύ�{active},PKR,PKR{active},PRAK,PRAK{p},Ras,Ras{active},SB203580,Sap1a,Sap1a{p},TAB2,TAB2:TAK1,TAK1,TLR4,TLR4:MD2,TRAF6,TRAF6{active},U0126,c-Raf,c-Raf{active},c-fos{p}:c-jun{p},c-jun:ATF2,c-jun{p}:ATF2{p},dexamethasone,genistein,p38,p38{active},p50:p65,p50:p65:Ikappa-Balpha,p50:p65:Ikappa-Balpha{p},p50:p65{p},p50:p65{p}:CBP,p53/56lyn,p53/56lyn{p},p53/64c-fgr{p},p58/64c-fgr,protein reminant of Ikappa-Balpha,unknown
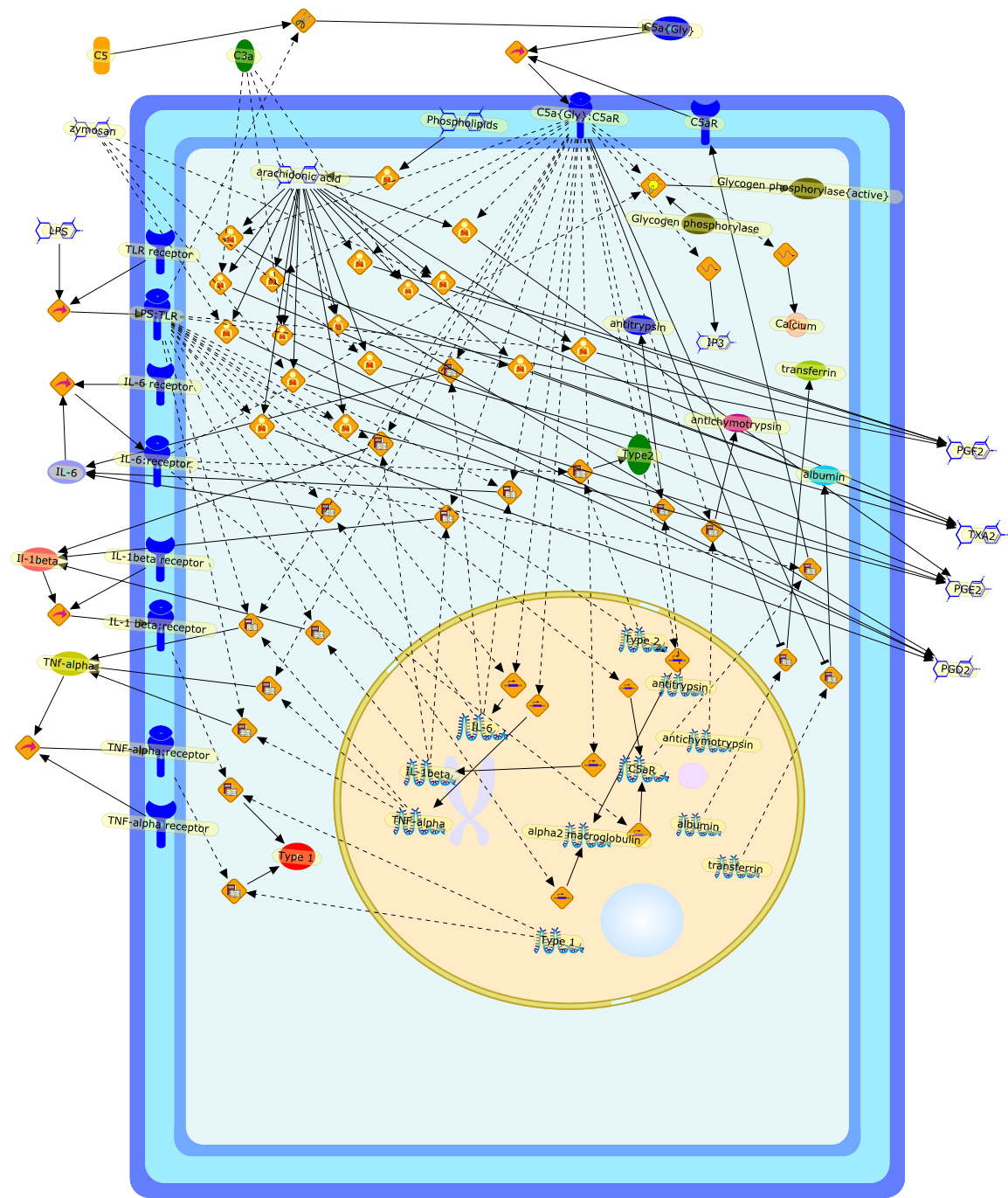
Growing evidence obtained in recent years indicates that anaphylatoxin C5areceptors (C5aR) are not restricted to myeloid cells but are also expressed onnonmyeloid cells in different tissues such as brain, lung, skin and liver. Incontrast to its well-defined systemic functions, the actions of anaphylatoxinsin these organs are poorly characterized. The liver can be a primary targetorgan for the C5a anaphylatoxin since the liver is directly connected to thegut, via the mesenteric veins and portal vein which is a main source ofcomplement activating lipopolysaccharides (LPS). In the normal rat liver, theC5aR is only expressed by nonparenchymal cells, i.e. strongly by Kupffer cells(KC) and hepatic stellate cells (HSC) and weakly by sinusoidal endothelial cells(SEC), but not expressed by the parenchymal hepatocytes (HC). Accordingly,direct effects of C5a were only found in the C5aR-expressing KC and HSC: C5ainduced the release of prostanoids from KC and HSC and enhanced theLPS-dependent release of interleukin-6 from KC. These soluble mediatorsindirectly influenced effector functions of the C5aR-free HC. C5a enhanced theglycogen phosphorylase activity and thus the glucose output from HC indirectlyvia prostanoids released from KC and HSC. Glucose can serve as an energysubstrate as well as an electron donor for the synthesis of reactive oxygenintermediates by KC. Moreover, C5a also enhanced transcription of the gene forthe type-2 acute phase protein alpha 2-macroglobulin in HC indirectly byincreasing LPS-dependent IL-6 release from KC. Under pathological conditions,C5aR was found to be upregulated in various organs including the liver.Simulation of inflammatory conditions by treatment of rats with IL-6, a maininflammatory mediator in the liver, caused a de novo expression of functionalC5aR in HC. In livers of IL-6-treated rats, C5a initiated glucose output from HCand perhaps other HC-specific defense reactions directly without theintervention of soluble mediators from nonparenchymal cells.
{kind=link}
{kind=link}
C3a,C5,C5aR,C5a{Gly},C5a{Gly}:C5aR,Calcium,Glycogen phosphorylase,Glycogen phosphorylase{active},IL-1 beta:receptor,IL-1beta,IL-1beta receptor,IL-6,IL-6 receptor,IL-6:receptor,IP3,Il-1beta,LPS,LPS:TLR,PGD2,PGE2,PGF2,Phospholipids,TLR receptor,TNF-alpha,TNF-alpha receptor,TNF-alpha:receptor,TNf-alpha,TXA2,Type 1,Type 2,Type2,albumin,alpha2 macroglobulin,antichymotrypsin,antitrypsin,arachidonic acid,transferrin,zymosan
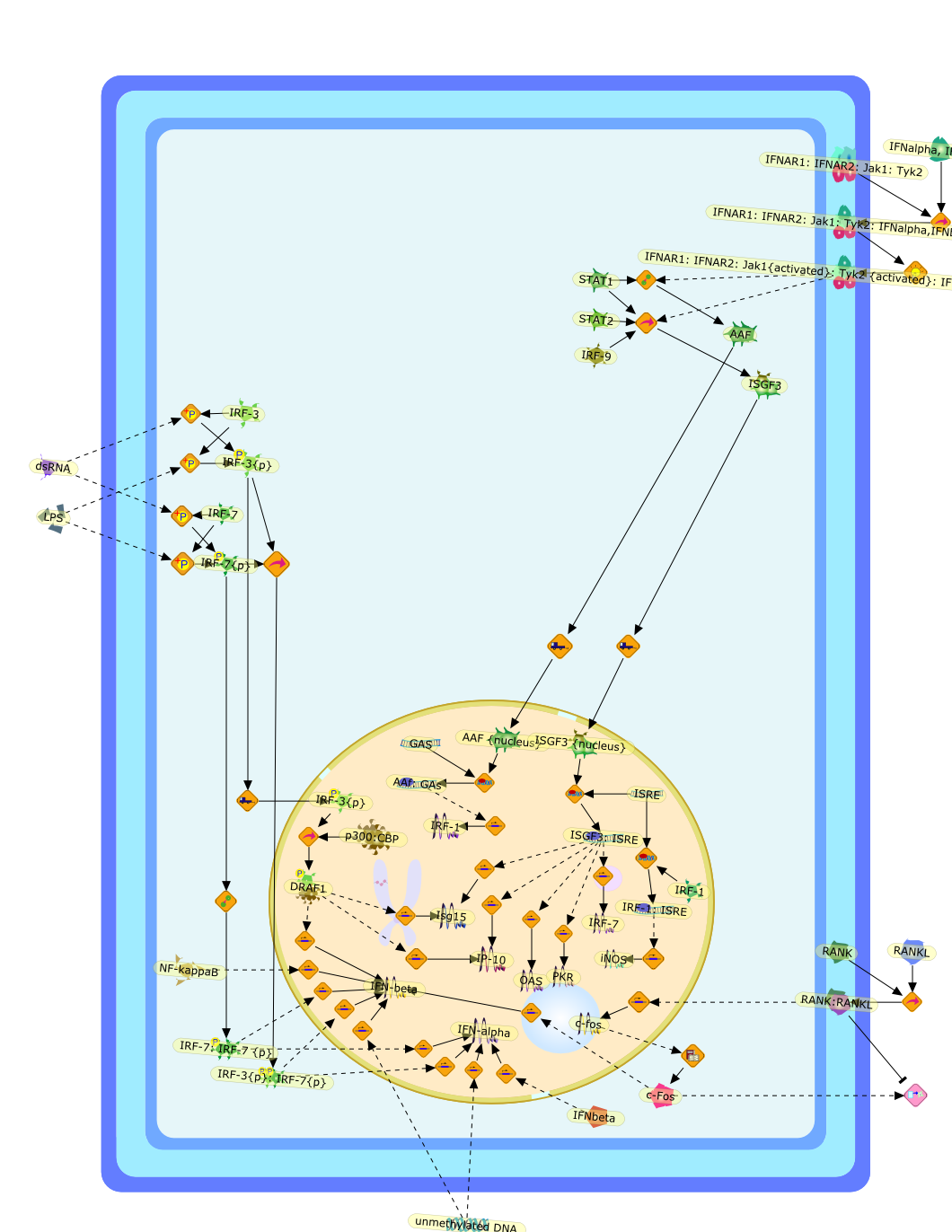
The efficient induction of interferons alpha and beta (IFN-alpha/beta) invirus-infected cells is central to the antiviral response of a host and isregulated mainly at the level of gene transcription. Once produced,IFN-alpha/beta transmit signals to the cell interior via a specific receptorcomplex to induce an antiviral response. Recently, the auto-amplificationmechanism of the IFN-alpha/beta system that follows viral infection has beenidentified. This mechanism is mediated by transcription factors of the IFNregulatory factor family and, in fact, may have evolved to render the systemmore robust in antiviral responses.
{kind=link}
{kind=link}
AAF,AAF {nucleus},AAf: GAs,DRAF1,GAS,IFN-alpha,IFN-beta,IFNAR1: IFNAR2: Jak1: Tyk2,IFNAR1: IFNAR2: Jak1: Tyk2: IFNalpha,IFNbeta,IFNAR1: IFNAR2: Jak1{activated}: Tyk2 {activated}: IFNalpha,IFNbeta,IFNalpha, IFNbeta,IFNbeta,IP-10,IRF-1,IRF-1: ISRE,IRF-3,IRF-3{p},IRF-3{p}: IRF-7{p},IRF-7,IRF-7: IRF-7 {p},IRF-7{p},IRF-9,ISGF3,ISGF3 {nucleus},ISGF3: ISRE,ISRE,Isg15,LPS,NF-kappaB,OAS,PKR,RANK,RANK:RANKL,RANKL,STAT1,STAT2,c-Fos,c-fos,dsRNA,iNOS,p300:CBP,unmethylated DNA
Sustained TNFalpha induction is central to the pathogenesis of chronic liverdisease including alcoholic liver disease (ALD). However, molecularunderstanding of this abnormality at the cellular level remains elusive. Redoxregulation of NF-kappaB is critical in the transcriptional control of TNFalphaexpression. Evidence supports that increased iron storage in hepatic macrophages(HM) is causally associated with accentuated and sustained NF-kappaB activationin these cells in ALD. Treatment of cultured HM with a lipophilic iron chelator(deferiprone) abrogates LPS-induced NF-kappaB activation. HM from an animalmodel of ALD have increased nonheme iron content accompanied by increasedgeneration of EPR-detected radicals, NF-kappaB activation, and TNFalphainduction, all of which are normalized by ex vivo treatment of the cells withdeferiprone. A moderate increase in the nonheme iron content in HM byerythrophagocytosis, promotes subsequent LPS-stimulated NF-kappaB activation ina hemeoxygenase-dependent manner. Recent evidence also suggests a role ofintracellular low molecular weight iron in the early signal transduction forLPS-mediated NF-kappaB activation.
{kind=link}
{kind=link}
4Fe-S4,Ferritin,H2O2,IKK,IKK{activated},IL-10,IL-4,IRP-1,Iron,LPS,NADPH oxidase,NADPH oxidase{activated},NFkappaB,NFkappaB{activated},NO,O2-,OH-,ONOO-,PMA,TNF-alpha,deferiprone,e17,iNOS,iNOS{activated},iron:nitrosyl,nitrosyl,zinc protoporphyrin
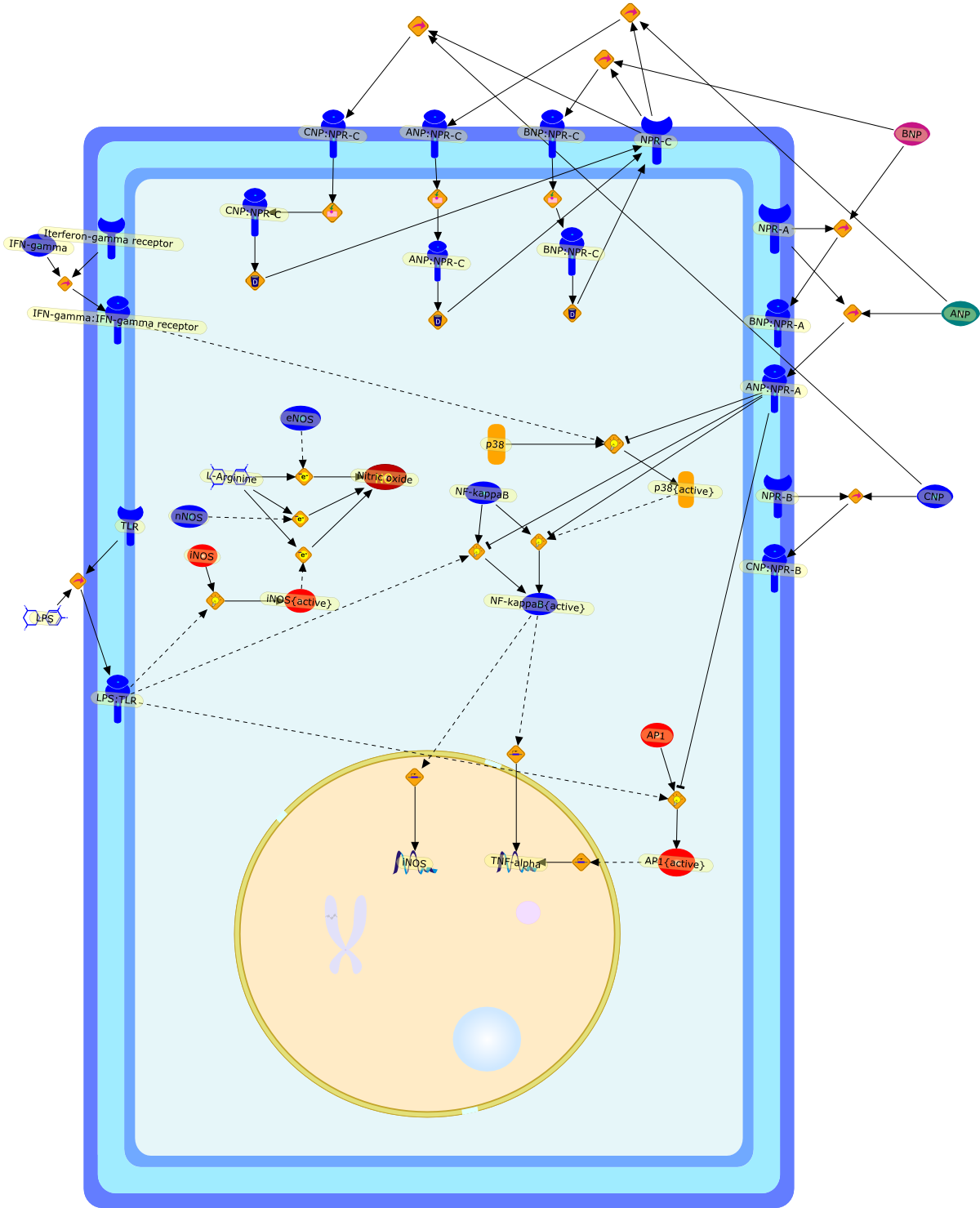
The atrial natriuretic peptide (ANP), a member of the natriuretic peptidefamily, is a cardiovascular hormone which possesses well defined natriuretic,diuretic, and vasodilating properties. Most of the biological effects of ANParemediated through its guanylyl cyclase coupled A receptor. Because ANP and itsreceptors have been shown to be expressed and differentially regulated in theimmune system, it has been suggested that ANP has an immunomodulatory potency.Much investigation of the effects of ANP on the activation of macrophages hasbeen carried out. ANP was shown to inhibit the lipopolysaccharide (LPS)-inducedexpression of inducible nitric oxide synthase (iNOS) in macrophages in anautocrine fashion. ANP in this context was shown to reduce significantly theactivation of NF-kappaB and to destabilise iNOS mRNA. ANP, furthermore, cansignificantly reduce the LPS-induced secretion of tumour necrosis factor alpha(TNFalpha) in macrophages. The relevance of these findings on a regulatory rolefor ANP on TNFalpha in humans was shown by the fact that ANP significantlyreduces the release of TNFalpha in whole human blood. It was furthermore shownto attenuate the release of interleukin 1beta (IL1beta). Interestingly, ANP didnot affect the secretion of the anti-inflammatory cytokines IL10 and IL1receptor antagonist (IL1ra). In summary, ANP was shown to reduce the secretionof inflammatory mediators in macrophages. Therefore, this cardiovascular hormonemay possess anti-inflammatory potential.
{kind=link}
{kind=link}
ANP,ANP:NPR-A,ANP:NPR-C,AP1,AP1{active},BNP,BNP:NPR-A,BNP:NPR-C,CNP,CNP:NPR-B,CNP:NPR-C,IFN-gamma,IFN-gamma:IFN-gamma receptor,Iterferon-gamma receptor,L-Arginine,LPS,LPS:TLR,NF-kappaB,NF-kappaB{active},NPR-A,NPR-B,NPR-C,Nitric oxide,TLR,TNF-alpha,eNOS,iNOS,iNOS{active},nNOS,p38,p38{active}
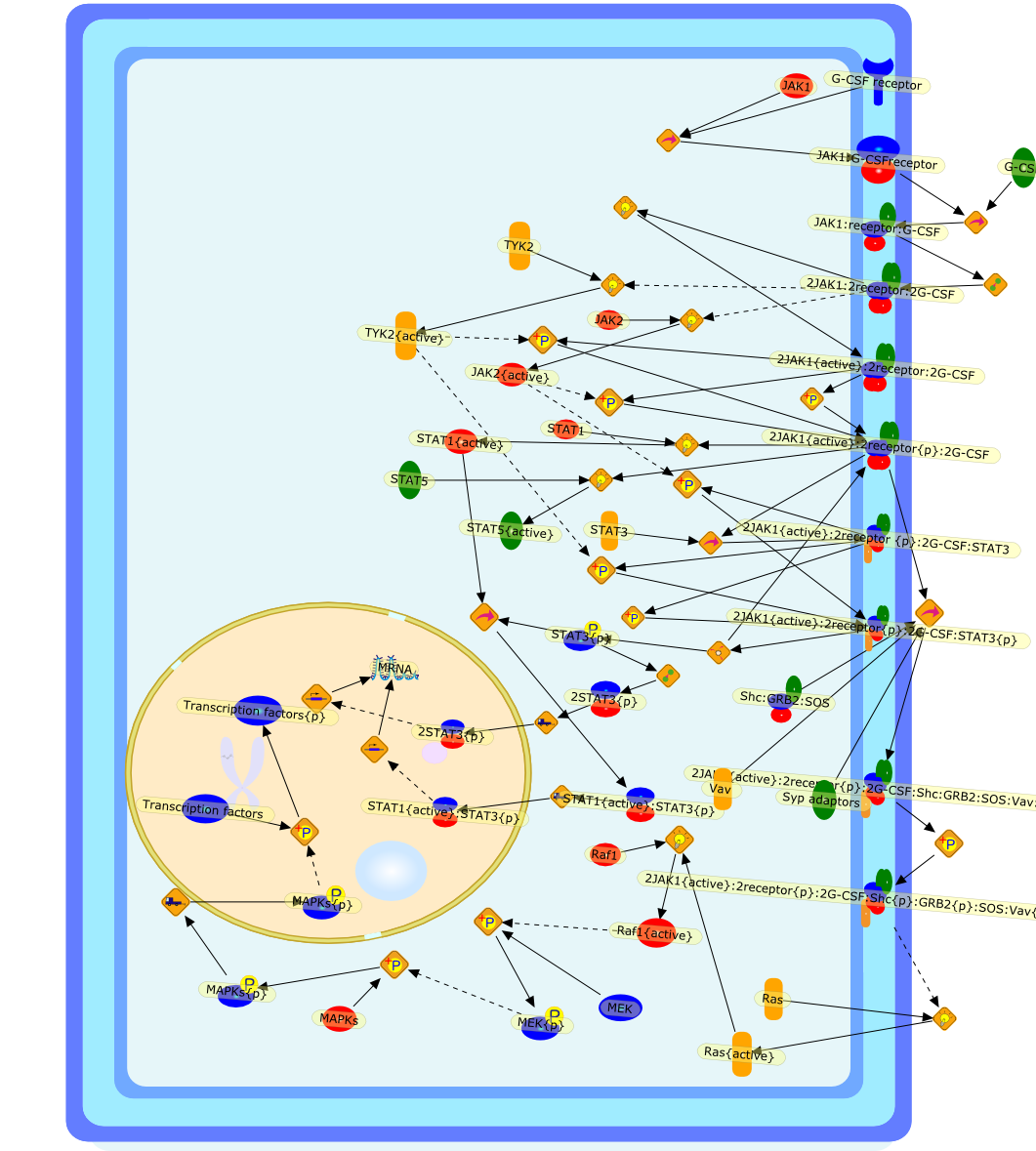
Granulocyte colony-stimulating factor (G-CSF) is a cytokine which stimulates theproduction of neutrophils in the bone marrow and modulates cellular functions ofmature neutrophils. Besides neutrophils and their precursors, monocytes aredirect target cells of G-CSF action. G-CSF influences monocyte functions in ananti-inflammatory way: The stimulation of monocytes with G-CSF results in anattenuation of LPS-induced release of IL-1beta, TNF-alpha, IL-12 and IL-18.G-CSF exerts its biological functions on neutrophils and monocytes viamembrane-bound receptors. Seven different human G-CSF receptor isoforms havebeen described which are generated by alternative splicing. The physiologicroles of these isoforms and the expression pattern on various cell types arestill unknown. The signal transduction pathway of G-CSF receptors involves theactivation of JAK tyrosine kinases and STAT transcription factors and theras/mitogen-activated protein kinase pathway.
{kind=link}
{kind=link}
2JAK1:2receptor:2G-CSF,2JAK1{active}:2receptor {p}:2G-CSF:STAT3,2JAK1{active}:2receptor:2G-CSF,2JAK1{active}:2receptor{p}:2G-CSF,2JAK1{active}:2receptor{p}:2G-CSF:STAT3{p},2JAK1{active}:2receptor{p}:2G-CSF:Shc:GRB2:SOS:Vav:Syp,2JAK1{active}:2receptor{p}:2G-CSF:Shc{p}:GRB2{p}:SOS:Vav{p}:Syp{p},2STAT3{p},G-CSF,G-CSF receptor,JAK1,JAK1:G-CSFreceptor,JAK1:receptor:G-CSF,JAK2,JAK2{active},MAPKs,MAPKs{p},MEK,MEK{p},MRNA,Raf1,Raf1{active},Ras,Ras{active},STAT1,STAT1{active},STAT1{active}:STAT3{p},STAT3,STAT3{p},STAT5,STAT5{active},Shc:GRB2:SOS,Syp adaptors,TYK2,TYK2{active},Transcription factors,Transcription factors{p},Vav
This review will focus on the molecular mechanisms of macrophage activation anddesensitization by bacterial lipopolysaccharide (LPS). The most recent advancesin the understanding of the function of the LPS receptor complex and its role inthe development of the septic shock syndrome and endotoxin tolerance will bediscussed.
{kind=link}
{kind=link}
AP-1,AP-1{active},CD11b:CD18,CD11c:CD18,CD14,CXCR4:GDF5:HSP90:HSP70,ERK1,ERK1{active},ERK2,ERK2{active},I-kappaB,ICSBP,IFN-gamma,IKKalpha:IKKbeta:IKKgamma,IKKalpha:Ikkbeta:IKKgamma{active},IL-12p35,IL-12p40,IL-8,IP-10,IRF-1,Ikappa-B:NF-kappaB,Ikappa-B{p}:NF-kappaB,Ikappa-B{ub}{NF-kappaB,JNK1,JNK1{active},JNK2,JNK2{active},LAM,LAM:TLR2,LPS,LPS:CD11b:CD18,LPS:CD11c:CD18,LPS:CD14,LPS:CD14:TLR4:MD2,LPS:CXCXR4:GDF5:HSP90:HSP70,LPS:Nod1,LPS:TLR2,MCP-5,MD2,NF-kappaB:,NF-kappaB{active},Nod1,STF,STF:TLR2,TLR2,TLR4,TLR4:MD2,TNF-alpha,cox2,p19/IL-13,p38,p38{active},protein remnants
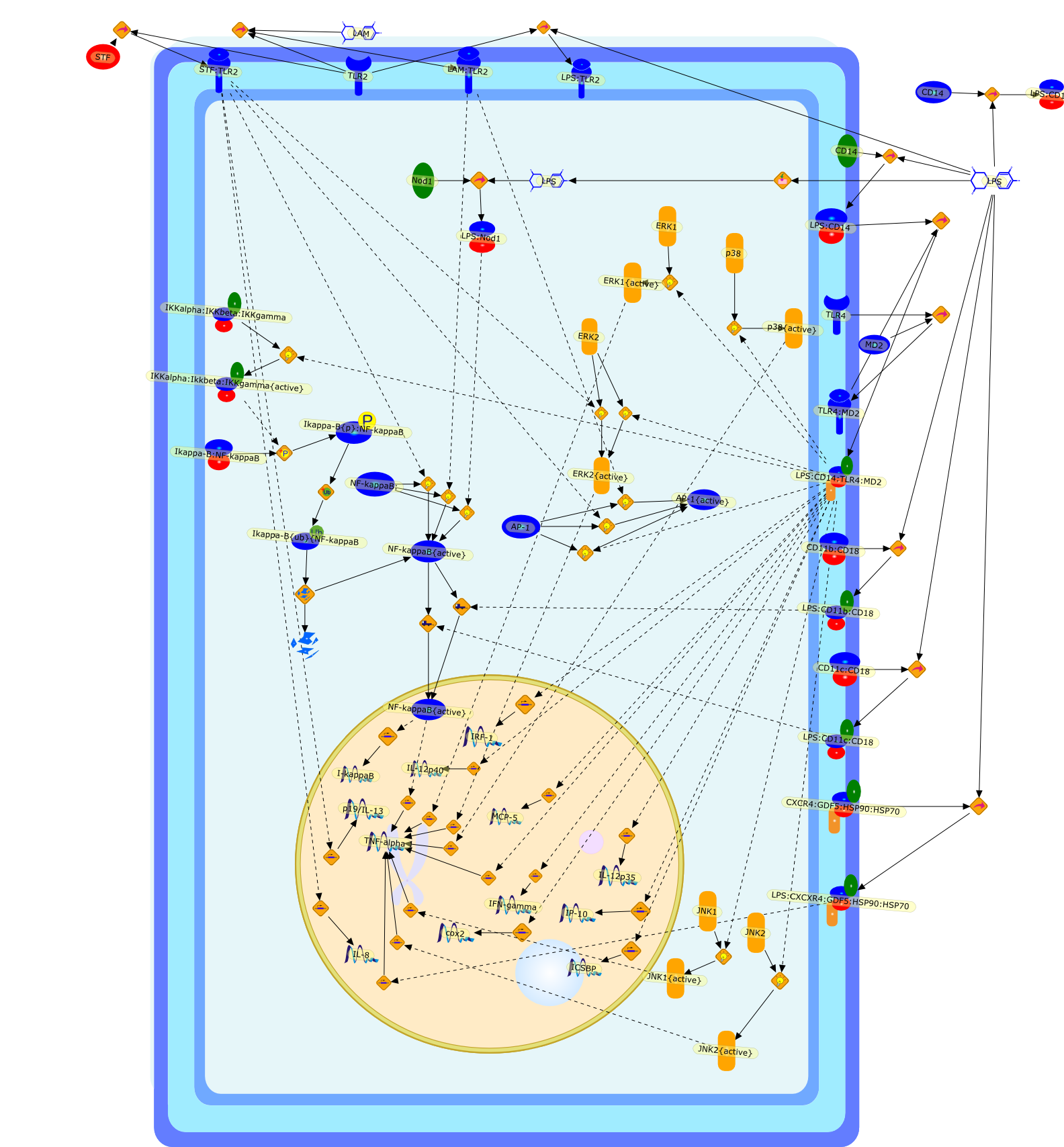
Bacterial lipopolysaccharides (LPSs) are powerful immunomodulators in infectedhosts, and may cause endotoxic shock. Most of them share a common architecturebut vary considerably in structural motifs from one genus, species, and strainto another. Cells of the innate immune response recognize evolutionarilyconserved LPS molecular patterns of endotoxins and structural details therebygreatly influencing their response.
{kind=link}
{kind=link}
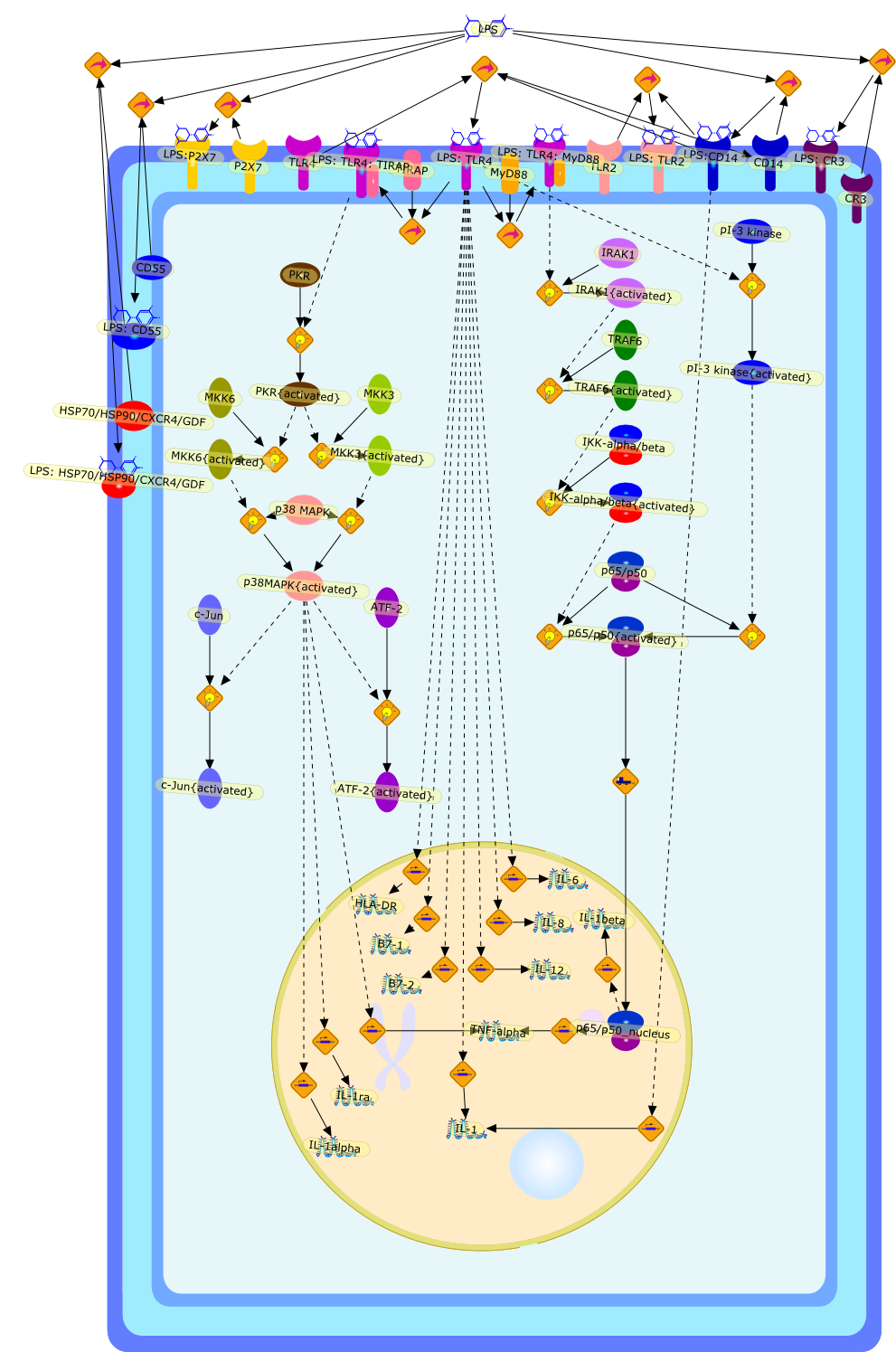
ATF-2,ATF-2{activated},B7-1,B7-2,CD14,CD55,CR3,HLA-DR,HSP70/HSP90/CXCR4/GDF,IKK-alpha/beta,IKK-alpha/beta{activated},IL-1,IL-12,IL-1alpha,IL-1beta,IL-1ra,IL-6,IL-8,IRAK1,IRAK1{activated},LPS,LPS: CD55,LPS: CR3,LPS: HSP70/HSP90/CXCR4/GDF,LPS: TLR2,LPS: TLR4,LPS: TLR4: MyD88,LPS: TLR4: TIRAP,LPS:CD14,LPS:P2X7,MKK3,MKK3{activated},MKK6,MKK6{activated},MyD88,P2X7,PKR,PKR{activated},TIRAP,TLR2,TLR4,TNF-alpha,TRAF6,TRAF6{activated},c-Jun,c-Jun{activated},p38 MAPK,p38MAPK{activated},p65/p50,p65/p50_nucleus,p65/p50{activated},pI-3 kinase,pI-3 kinase{activated}
Virus infection of susceptible cells activates multiple signaling pathways thatorchestrate the activation of genes, such as cytokines, involved in theantiviral and innate immune response. Among the kinases induced are themitogen-activated protein (MAP) kinases, Jun-amino terminal kinases (JNK) andp38, the IkappaB kinase (IKK) and DNA-PK. In addition, virus infection alsoactivates an uncharacterized VAK responsible for the C-terminal phosphorylationand subsequent activation of interferon regulatory factor 3 (IRF-3).Virus-mediated activation of IRF-3 through VAK is dependent on viral entry andtranscription, since replication deficient virus failed to induce IRF-3activity. The pathways leading to VAK activation are not well characterized, butIRF-3 appears to represent a novel cellular detection pathway that recognizesviral nucleocapsid (N) structure. Recently, the range of inducers responsiblefor IRF-3 activation has increased. In addition to virus infection, recognitionof bacterial infection mediated through lipopolysaccharide by Toll-like receptor4 has also been reported. Furthermore, MAP kinase kinase kinase (MAPKKK)-related pathways and DNA-PK induce N-terminal phosphorylation of IRF-3.This review summarizes recent observations in the identification of novelsignaling pathways leading to IRF-3 activation.
{kind=link}
{kind=link}
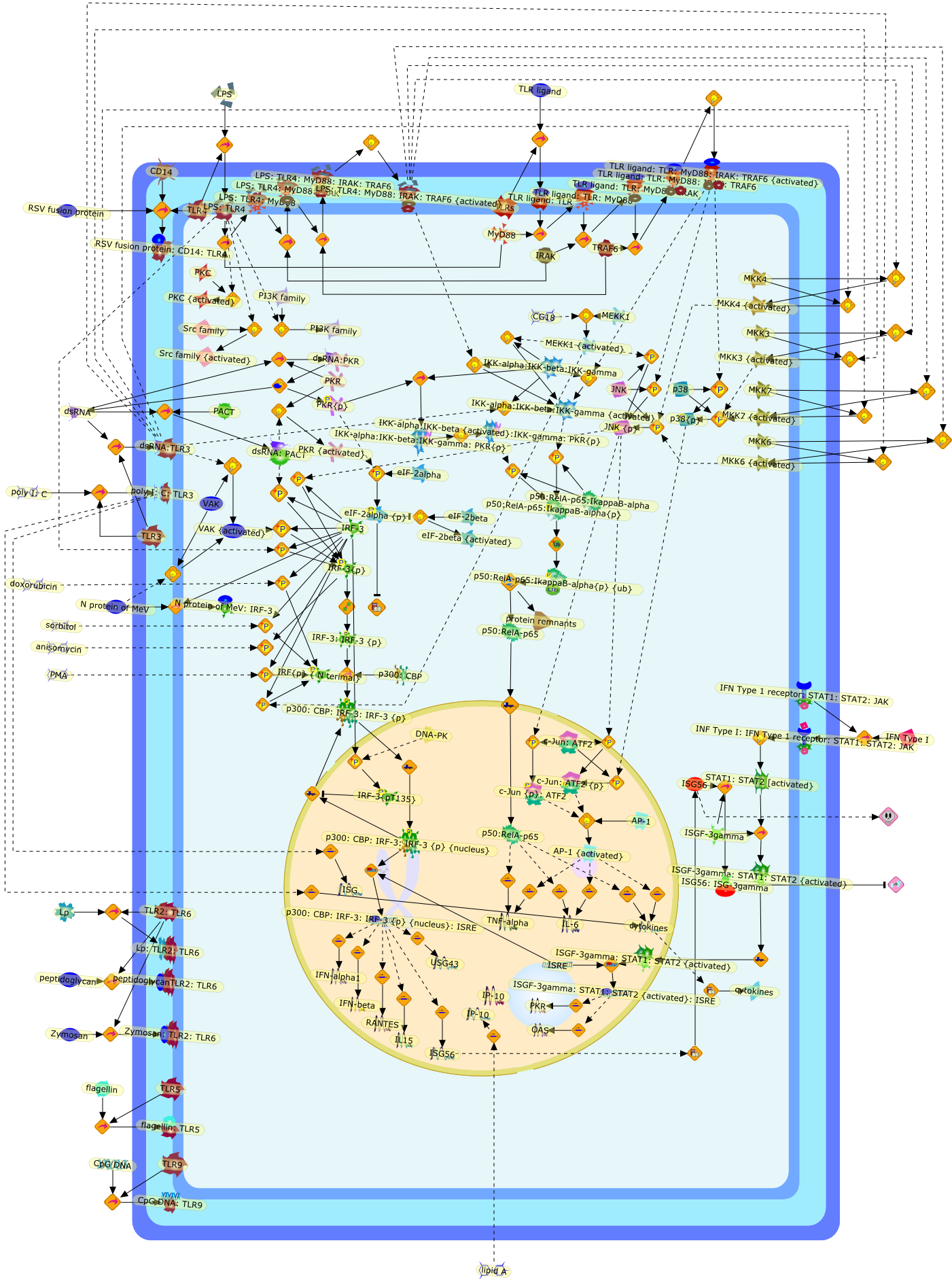
AP-1,AP-1 {activated},CD14,CG18,CpG DNA,CpG DNA: TLR9,DNA-PK,IFN Type 1 receptor: STAT1: STAT2: JAK,IFN Type I,IFN-alpha1,IFN-beta,IKK-alpha:IKK-beta {activated}:IKK-gamma: PKR{p},IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma {activated},IKK-alpha:IKK-beta:IKK-gamma: PKR{p},IL-6,IL15,INF Type I: IFN Type 1 receptor: STAT1: STAT2: JAK,IP-10,IRAK,IRF-3,IRF-3: IRF-3 {p},IRF-3{pT135},IRF-3{p},IRF{p} { N terimal},ISG,ISG56,ISG56: ISG-3gamma,ISGF-3gamma,ISGF-3gamma: STAT1: STAT2 {activated},ISGF-3gamma: STAT1: STAT2 {activated}: ISRE,ISRE,JNK,JNK {p},LPS,LPS: TLR4,LPS: TLR4: MyD88,LPS: TLR4: MyD88: IRAK,LPS: TLR4: MyD88: IRAK: TRAF6,LPS: TLR4: MyD88: IRAK: TRAF6 {activated},Lp,Lp: TLR2: TLR6,MEKK1,MEKK1 {activated},MKK3,MKK3 {activated},MKK4,MKK4 {activated},MKK6,MKK6 {activated},MKK7,MKK7 {activated},MyD88,N protein of MeV,N protein of MeV: IRF-3,OAS,PACT,PI3K family,PKC,PKC {activated},PKR,PKR {activated},PKR{p},PMA,RANTES,RSV fusion protein,RSV fusion protein: CD14: TLR4,STAT1: STAT2 [activated},Src family,Src family {activated},TLR ligand,TLR ligand: TLR,TLR ligand: TLR: MyD88,TLR ligand: TLR: MyD88: IRAK,TLR ligand: TLR: MyD88: IRAK: TRAF6,TLR ligand: TLR: MyD88: IRAK: TRAF6 {activated},TLR2: TLR6,TLR3,TLR4,TLR5,TLR9,TLRs,TNF-alpha,TRAF6,USG43,VAK,VAK {activated},Zymosan,Zymosan: TLR2: TLR6,anisomycin,c-Jun {p}: ATF2,c-Jun: ATF2,c-Jun: ATF2 {p},cytokines,doxorubicin,dsRNA,dsRNA: PACT,dsRNA:PKR,dsRNA:TLR3,eIF-2alpha,eIF-2alpha {p},eIF-2beta,eIF-2beta {activated},flagellin,flagellin: TLR5,lipid A,p300: CBP,p300: CBP: IRF-3: IRF-3 {p},p300: CBP: IRF-3: IRF-3 {p} {nucleus},p300: CBP: IRF-3: IRF-3 {p} {nucleus}: ISRE,p38,p38{p},p50:RelA-p65,p50:RelA-p65:IkappaB-alpha,p50:RelA-p65:IkappaB-alpha{p},p50:RelA-p65:IkappaB-alpha{p} {ub},peptidoglycan,peptidoglycanTLR2: TLR6,poly I: C,poly I: C: TLR3,protein remnants,sorbitol
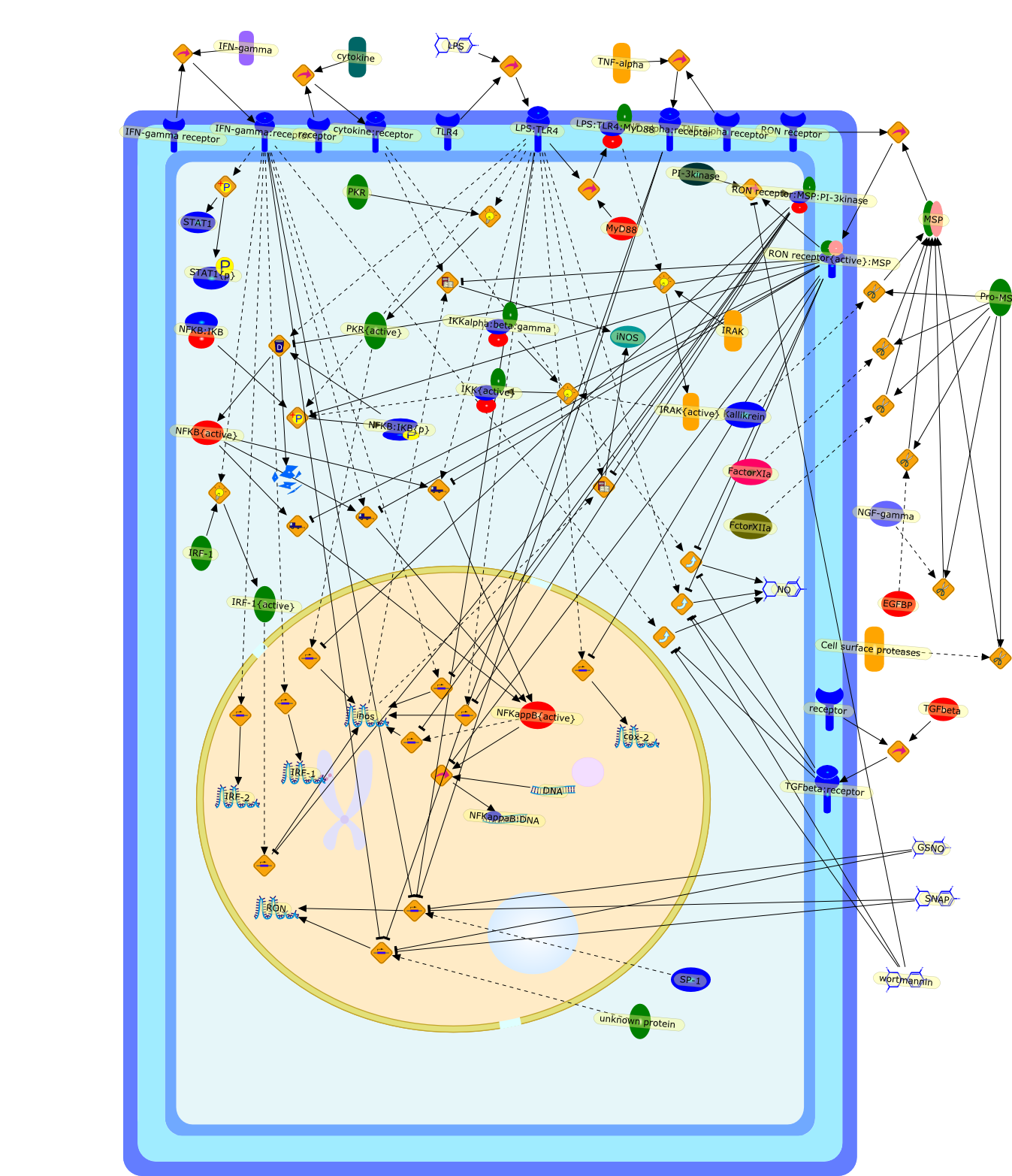
Macrophage-stimulating protein (MSP) is a serum protein belonging to theplasminogen-related growth factor family. The specific receptor for MSP is theRON (recepteur d'origine nantais) receptor tyrosine kinase - a member of the METproto-oncogene family. Activation of RON by MSP exerts dual functions onmacrophages. The stimulatory activities include the induction of macrophagespreading, migration and phagocytosis. However, MSP also inhibitslipopolysaccharide (LPS)-induced production of inflammatory mediators, includinginducible nitric oxide and prostaglandins. These suppressive effects aremediated by RON-transduced signals that block LPS-induced enzymatic cascadesthat activate nuclear factor kappa-B (NFkappaB) pathways. Recent in vivo studiesdemonstrated that inactivation of the RON gene results in increased inflammatoryresponses and susceptibility to LPS-induced septic death in mice, suggestingthat RON expression is required for attenuating the extent of inflammatoryresponses in vivo. Thus, MSP and RON are potential regulators that controlmacrophage activities during bacterial infection in vivo.
{kind=link}
{kind=link}
Cell surface proteases,DNA,EGFBP,FactorXIa,FctorXIIa,GSNO,IFN-gamma,IFN-gamma receptor,IFN-gamma:receptor,IKKalpha:beta:gamma,IKK{active},IRAK,IRAK{active},IRF-1,IRF-1{active},IRF-2,Kallikrein,LPS,LPS:TLR4,LPS:TLR4:MyD88,MSP,MyD88,NFKB:IKB,NFKB:IKB{p},NFKB{active},NFKappB{active},NFKappaB:DNA,NGF-gamma,NO,PI-3kinase,PKR,PKR{active},Pro-MSP,RON,RON receptor,RON receptor:MSP:PI-3kinase,RON receptor{active}:MSP,SNAP,SP-1,STAT1,STAT1{p},TGFbeta,TGFbeta:receptor,TLR4,TNF-alpha,TNF-alpha receptor,TNF_alpha:receptor,cox-2,cytokine,cytokine:receptor,iNOS,inos,protein remnants,receptor,unknown protein,wortmannin
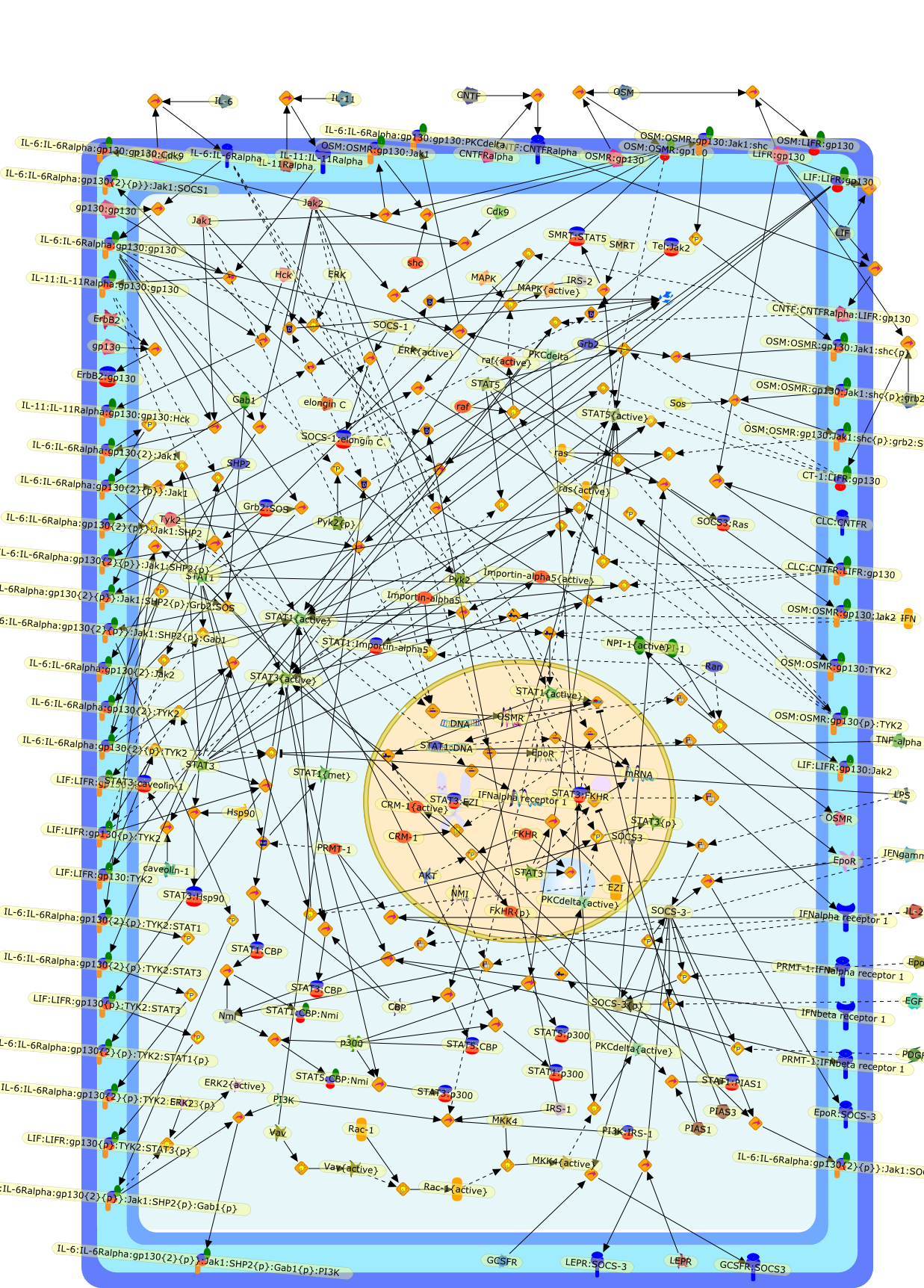
The IL (interleukin)-6-type cytokines IL-6, IL-11, LIF (leukaemia inhibitoryfactor), OSM (oncostatin M), ciliary neurotrophic factor, cardiotrophin-1 andcardiotrophin-like cytokine are an important family of mediators involved in theregulation of the acute-phase response to injury and infection. Besides theirfunctions in inflammation and the immune response, these cytokines play also acrucial role in haematopoiesis, liver and neuronal regeneration, embryonaldevelopment and fertility. Dysregulation of IL-6-type cytokine signallingcontributes to the onset and maintenance of several diseases, such as rheumatoidarthritis, inflammatory bowel disease, osteoporosis, multiple sclerosis andvarious types of cancer (e.g. multiple myeloma and prostate cancer). IL-6-typecytokines exert their action via the signal transducers gp (glycoprotein) 130,LIF receptor and OSM receptor leading to the activation of the JAK/STAT (Januskinase/signal transducer and activator of transcription) and MAPK(mitogen-activated protein kinase) cascades. This review focuses on recentprogress in the understanding of the molecular mechanisms of IL-6-type cytokinesignal transduction. Emphasis is put on the termination and modulation of theJAK/STAT signalling pathway mediated by tyrosine phosphatases, the SOCS(suppressor of cytokine signalling) feedback inhibitors and PIAS (proteininhibitor of activated STAT) proteins. Also the cross-talk between the JAK/STATpathway with other signalling cascades is discussed.
{kind=link}
{kind=link}
AKT,CBP,CLC:CNTFR,CLC:CNTFR:LIFR:gp130,CNTF,CNTF:CNTFRalpha,CNTF:CNTFRalpha:LIFR:gp130,CNTFRalpha,CRM-1,CRM-1{active},CT-1,CT-1:LIFR:gp130,Cdk9,DNA,EGF,ERK,ERK2,ERK2{active},ERK{active},EZI,Epo,EpoR,EpoR:SOCS-3,ErbB2,ErbB2:gp130,FKHR,FKHR{p},GCSFR,GCSFR:SOCS3,Gab1,Grb2,Grb2:SOS,Hck,Hsp90,IFN,IFNalpha receptor 1,IFNalpha receptor 1 ,IFNbeta receptor 1,IFNgamma,IL-11,IL-11:IL-11Ralpha,IL-11:IL-11Ralpha:gp130:gp130,IL-11:IL-11Ralpha:gp130:gp130:Hck,IL-11Ralpha,IL-2,IL-6,IL-6:IL-6Ralpha,IL-6:IL-6Ralpha:gp130:gp130,IL-6:IL-6Ralpha:gp130:gp130:Cdk9,IL-6:IL-6Ralpha:gp130:gp130:PKCdelta,IL-6:IL-6Ralpha:gp130{2}:Jak1,IL-6:IL-6Ralpha:gp130{2}:Jak2,IL-6:IL-6Ralpha:gp130{2}:TYK2,IL-6:IL-6Ralpha:gp130{2}{p}:TYK2,IL-6:IL-6Ralpha:gp130{2}{p}:TYK2:STAT1,IL-6:IL-6Ralpha:gp130{2}{p}:TYK2:STAT1{p},IL-6:IL-6Ralpha:gp130{2}{p}:TYK2:STAT3,IL-6:IL-6Ralpha:gp130{2}{p}:TYK2:STAT3{p},IL-6:IL-6Ralpha:gp130{2}{p}}:Jak1,IL-6:IL-6Ralpha:gp130{2}{p}}:Jak1:SHP2,IL-6:IL-6Ralpha:gp130{2}{p}}:Jak1:SHP2{p},IL-6:IL-6Ralpha:gp130{2}{p}}:Jak1:SHP2{p}:Gab1,IL-6:IL-6Ralpha:gp130{2}{p}}:Jak1:SHP2{p}:Gab1{p},IL-6:IL-6Ralpha:gp130{2}{p}}:Jak1:SHP2{p}:Gab1{p}:PI3K,IL-6:IL-6Ralpha:gp130{2}{p}}:Jak1:SHP2{p}:Grb2:SOS,IL-6:IL-6Ralpha:gp130{2}{p}}:Jak1:SOCS1,IL-6:IL-6Ralpha:gp130{2}{p}}:Jak1:SOCS3,IL-6Ralpha,IRS-1,IRS-2,Importin-alpha5,Importin-alpha5{active},Jak1,Jak2,LEPR,LEPR:SOCS-3,LIF,LIF:LIFR:gp130,LIF:LIFR:gp130:Jak1,LIF:LIFR:gp130:Jak2,LIF:LIFR:gp130:TYK2,LIF:LIFR:gp130{p}:TYK2,LIF:LIFR:gp130{p}:TYK2:STAT3,LIF:LIFR:gp130{p}:TYK2:STAT3{p},LIFR:gp130,LPS,MAPK,MAPK{active},MKK4,MKK4{active},NMI,NPI-1,NPI-1{active},Nmi,OSM,OSM:LIFR:gp130,OSM:OSMR:gp130,OSM:OSMR:gp130:Jak1,OSM:OSMR:gp130:Jak1:shc,OSM:OSMR:gp130:Jak1:shc{p},OSM:OSMR:gp130:Jak1:shc{p}:grb2,OSM:OSMR:gp130:Jak1:shc{p}:grb2:SOS,OSM:OSMR:gp130:Jak2,OSM:OSMR:gp130:TYK2,OSM:OSMR:gp130{p}:TYK2,OSMR,OSMR:gp130,PDGF,PI3K,PI3K:IRS-1,PIAS1,PIAS3,PKCdelta,PKCdelta{active},PRMT-1,PRMT-1:IFNalpha receptor 1,PRMT-1:IFNbeta receptor 1,Pyk2,Pyk2{p},Rac-1,Rac-1{active},Ran,SHP2,SMRT,SMRT:STAT5,SOCS-1,SOCS-1:elongin C,SOCS-3,SOCS-3{p},SOCS3,SOCS3:Ras,STAT1,STAT1:CBP,STAT1:CBP:Nmi,STAT1:DNA,STAT1:Importin-alpha5,STAT1:PIAS1,STAT1:p300,STAT1{active},STAT1{met},STAT3,STAT3:CBP,STAT3:EZI,STAT3:FKHR,STAT3:Hsp90,STAT3:caveolin-1,STAT3:p300,STAT3{active},STAT3{p},STAT5,STAT5:CBP,STAT5:CBP:Nmi,STAT5:p300,STAT5{active},Sos,TNF-alpha,Tel:Jak2,Tyk2,Vav,Vav{active},caveolin-1,degradants,elongin C,gp130,gp130:gp130,mRNA,p300,raf,raf{active},ras,ras{active},shc
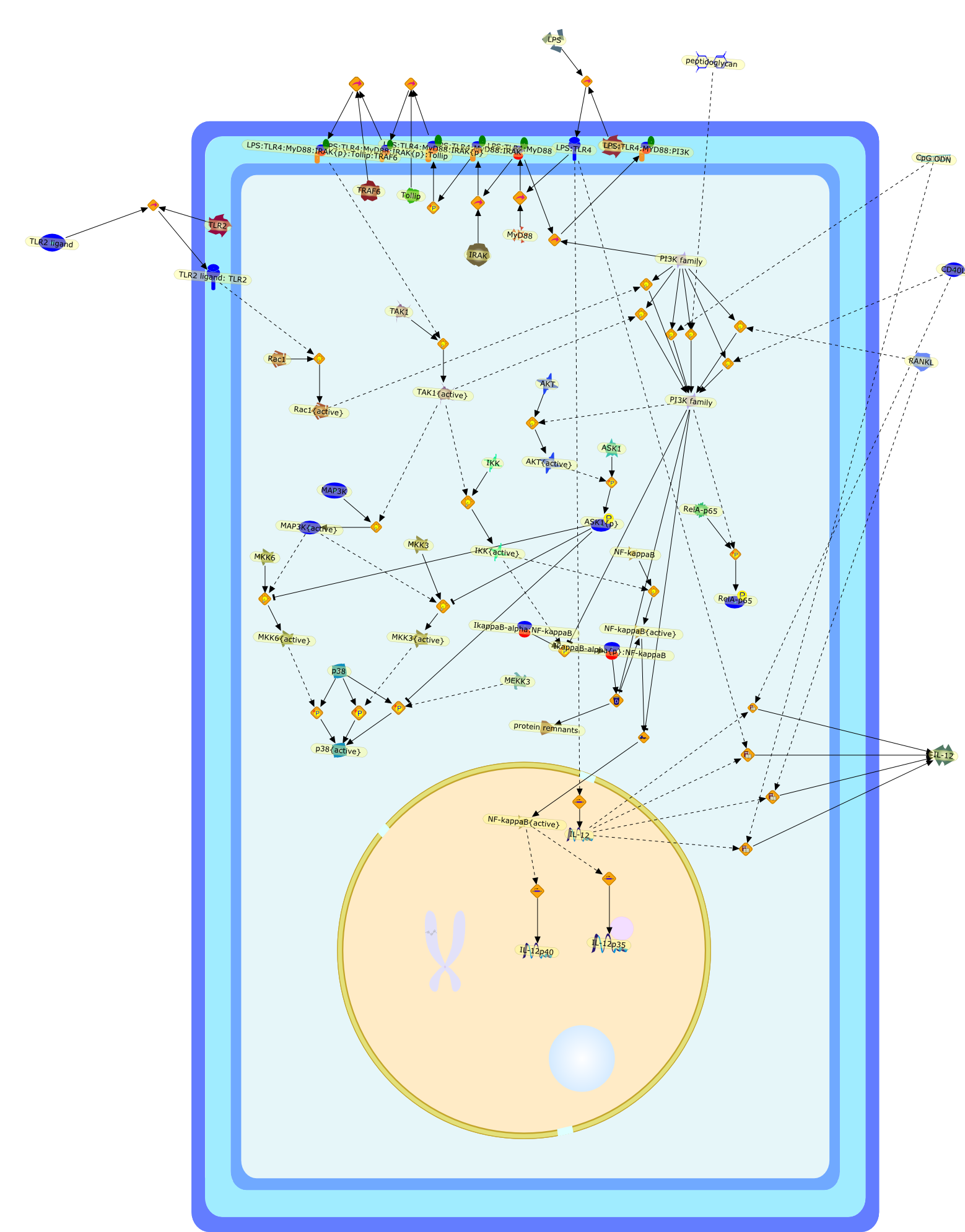
Excessive immune responses are detrimental to the host and negative feedbackregulation is crucial for the maintenance of immune-system integrity. Recentstudies have shown that phosphoinositide 3-kinase (PI3K) is an endogenoussuppressor of interleukin-12 (IL-12) production triggered by Toll-like receptor(TLR) signaling and limits excessive Th1 polarization. Unlike IRAK-M (IL-1receptor-associated kinase-M) and SOCS-1 (suppressor of cytokine signaling-1)that are induced by TLR signaling and function during the second or continuousexposure to stimulation, PI3K functions at the early phase of TLR signaling andmodulates the magnitude of the primary activation. Thus, PI3K, IRAK-M and SOCS-1have unique roles in the gate-keeping system, preventing excessive innate immuneresponses.
{kind=link}
{kind=link}
AKT,AKT{active},ASK1,ASK1{p},CD40L,CpG ODN,IKK,IKK{active},IL-12,IL-12p35,IL-12p40,IRAK,IkappaB-alpha:NF-kappaB,IkappaB-alpha{p}:NF-kappaB,LPS,LPS:TLR4,LPS:TLR4:MYD88:PI3K,LPS:TLR4:MyD88,LPS:TLR4:MyD88:IRAK,LPS:TLR4:MyD88:IRAK{p},LPS:TLR4:MyD88:IRAK{p}:Tollip,LPS:TLR4:MyD88:IRAK{p}:Tollip:TRAF6,MAP3K,MAP3K{active},MEKK3,MKK3,MKK3{active},MKK6,MKK6{active},MyD88,NF-kappaB,NF-kappaB{active},PI3K family,RANKL,Rac1,Rac1{active},RelA-p65,TAK1,TAK1{active},TLR2,TLR2 ligand,TLR2 ligand: TLR2,TLR4,TRAF6,Tollip,p38,p38{active},peptidoglycan,protein remnants
This short review focuses on the role of central nervous system (CNS)perivascular macrophages as targets of productive infection of the CNS. Datadiscussed include the importance of these cells as early targets of infectionand their productive infection with AIDS. Many of the immune molecules onperivascular macrophages are also found on subsets of bloodmonocyte/macrophages, some of which are expanded during human immunodeficiencyvirus (HIV) infection. These observations paired with the known bone marrow (BM)origin of perivascular macrophages and the BM as a site of HIV infectionunderscore the importance of the study of monocyte populations in the BM andblood, which are activated and infected as a source of virus that enters theCNS. Data presented and discussed herein suggest a role of HIV-infectedBM-derived monocytes as "Trojan horse" cells that traffic to the CNS to becomeperivascular macrophages. The study of such cells including their timing ofinfection, activation, and traffic and the role of HIV-specific immune responsescontrolling their accumulation in the CNS warrant study with regard to CNSneuropathogenesis.
{kind=link}
{kind=link}
CD14,CD16,CD45,CD69,HIV protein,HLA-DRA,M-CSF,PCNA,SIV RNA,gp120
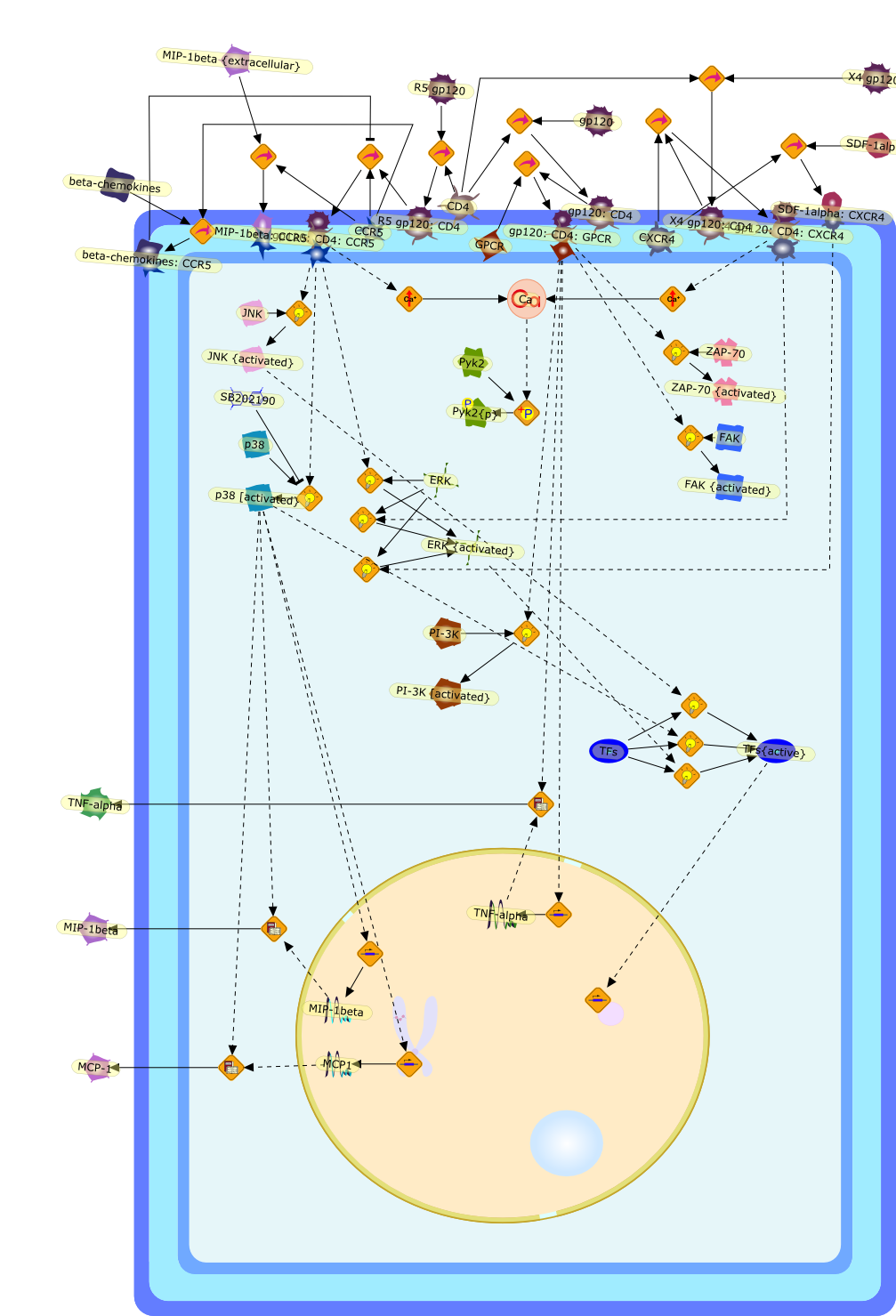
Macrophages are major targets for infection by human immunodeficiency virus type1 (HIV-1). In addition to their role as productive viral reservoirs,inappropriate activation of infected and uninfected macrophages appears tocontribute to pathogenesis. HIV-1 infection requires initial interactionsbetween the viral envelope surface glycoprotein gp120, the cell-surface proteinCD4, and a chemokine receptor CCR5 or CXCR4. Besides their role in HIV-1 entry,CCR5 and CXCR4 are G protein-coupled receptors that can activate multipleintracellular signaling pathways. HIV-1 gp120 has been shown to activatesignaling pathways through the chemokine receptors in several cell typesincluding lymphocytes, neurons, and astrocytes. In some cell types, theseconsequences may cause cellular injury. In this review, we highlight our datademonstrating diverse signaling events that occur in primary human macrophagesin response to gp120/chemokine receptor interactions. These responses includeK+, Cl-, and nonselective cation currents, intracellular Ca2+ increases, andactivation of several kinases including the focal adhesion-related tyrosinekinase Pyk2, mitogen-activated protein kinases (MAPK), and phosphoinositol-3kinase. Activation of the MAPK leads to gp120-induced expression of chemokinessuch as monocyte chemoattractant protein-1 and macrophage-inflammatoryprotein-1beta and the proinflammatory cytokine tumor necrosis factor alpha.These responses establish a complex cytokine network, which may enhance orsuppress HIV-1 replication. In addition, dysregulation of macrophage function bygp120/chemokine receptor signaling may contribute to local inflammation andinjury and further recruit additional inflammatory and/or target cells.Targeting these cellular signaling pathways may have benefit in controllinginflammatory sequelae of HIV infection such as in neurological disease.
{kind=link}
{kind=link}
CCR5,CD4,CXCR4,Ca,ERK,ERK {activated},FAK,FAK {activated},GPCR,JNK,JNK {activated},MCP-1,MCP1,MIP-1beta,MIP-1beta {extracellular},MIP-1beta: CCR5,PI-3K,PI-3K [activated},Pyk2,Pyk2{p},R5 gp120,R5 gp120: CD4,R5 gp120: CD4: CCR5,SB202190,SDF-1alpha,SDF-1alpha: CXCR4,TFs,TFs{active},TNF-alpha,X4 gp120,X4 gp120: CD4,X4 gp120: CD4: CXCR4,ZAP-70,ZAP-70 {activated},beta-chemokines,beta-chemokines: CCR5,gp120,gp120: CD4,gp120: CD4: GPCR,p38,p38 [activated}
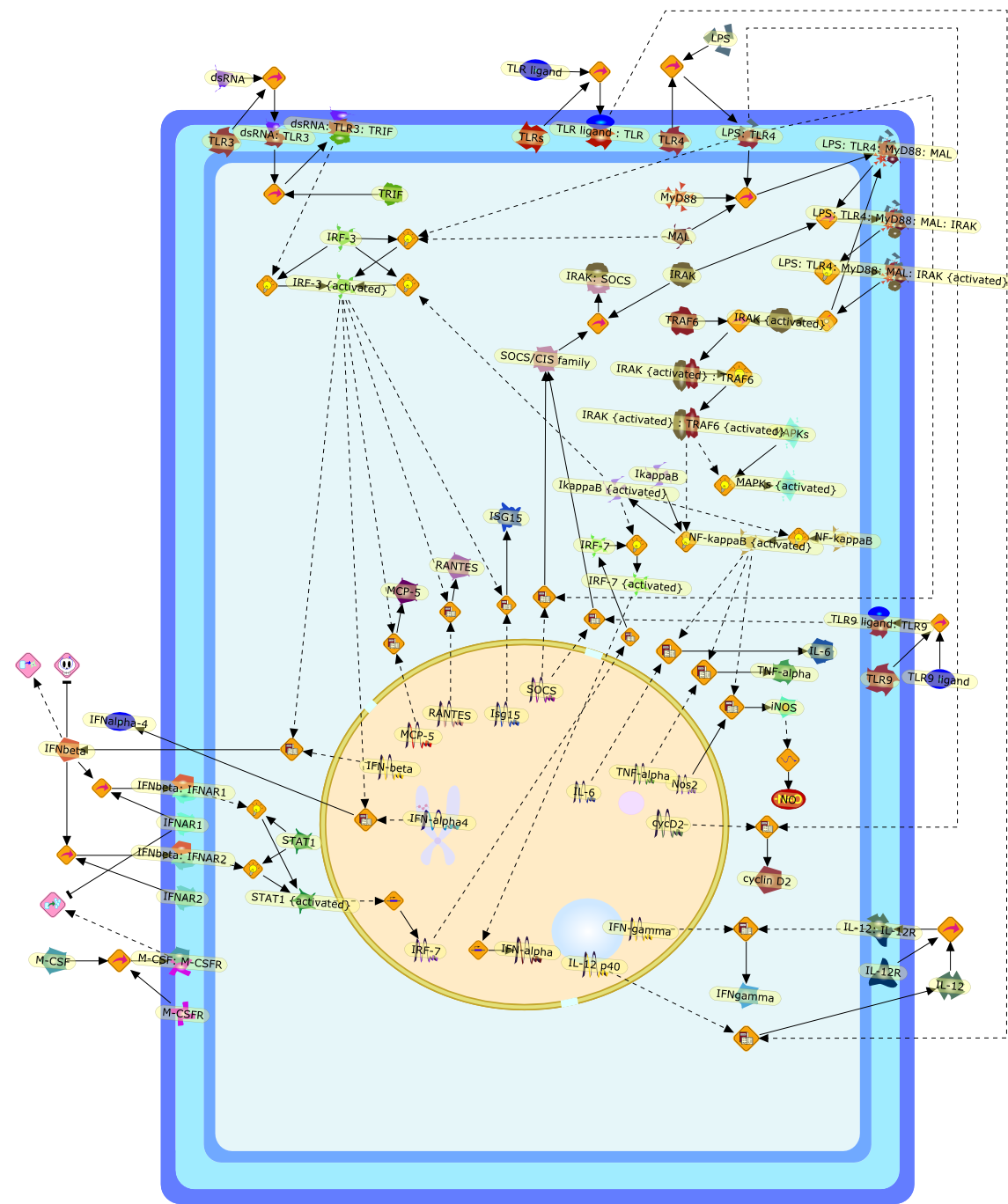
The Toll-like receptor (TLR) system is responsible for the recognition ofinfectious agents leading to initiation of the primary innate, and lateradaptive, immune response. Genetic technologies have enabled the discovery ofnew factors involved in these systems, their genetic manipulation and the globalanalyses of their effects on gene expression. Furthermore, this increasedunderstanding has resulted in the need to reassess our preconceptions about thefunctions of well-known molecules. For example, type I interferons (IFNs), whichwere discovered as antiviral proteins, are now known to be produced in responseto TLR activation by many pathogens, including bacteria. Should we be surprised?Has the inflammatory response unexpectedly highjacked the body's antiviralsystem? Or are we too easily blinkered by preconceptions from how a compound wasdiscovered?
{kind=link}
{kind=link}
IFN-alpha,IFN-alpha4,IFN-beta,IFN-gamma,IFNAR1,IFNAR2,IFNalpha-4,IFNbeta,IFNbeta: IFNAR1,IFNbeta: IFNAR2,IFNgamma,IL-12,IL-12 p40,IL-12: IL-12R,IL-12R,IL-6,IRAK,IRAK {activated},IRAK {activated} : TRAF6,IRAK {activated} : TRAF6 {activated},IRAK: SOCS,IRF-3,IRF-3 {activated},IRF-7,IRF-7 {activated},ISG15,IkappaB,IkappaB {activated},Isg15,LPS,LPS: TLR4,LPS: TLR4: MyD88: MAL,LPS: TLR4: MyD88: MAL: IRAK,LPS: TLR4: MyD88: MAL: IRAK {activated},M-CSF,M-CSF: M-CSFR,M-CSFR,MAL,MAPKs,MAPKs {activated},MCP-5,MyD88,NF-kappaB,NF-kappaB {activated},NO,Nos2,RANTES,SOCS,SOCS/CIS family,STAT1,STAT1 {activated},TLR ligand,TLR ligand : TLR,TLR3,TLR4,TLR9,TLR9 ligand,TLR9 ligand: TLR9,TLRs,TNF-alpha,TRAF6,TRIF,cycD2,cyclin D2,dsRNA,dsRNA: TLR3,dsRNA: TLR3: TRIF,iNOS
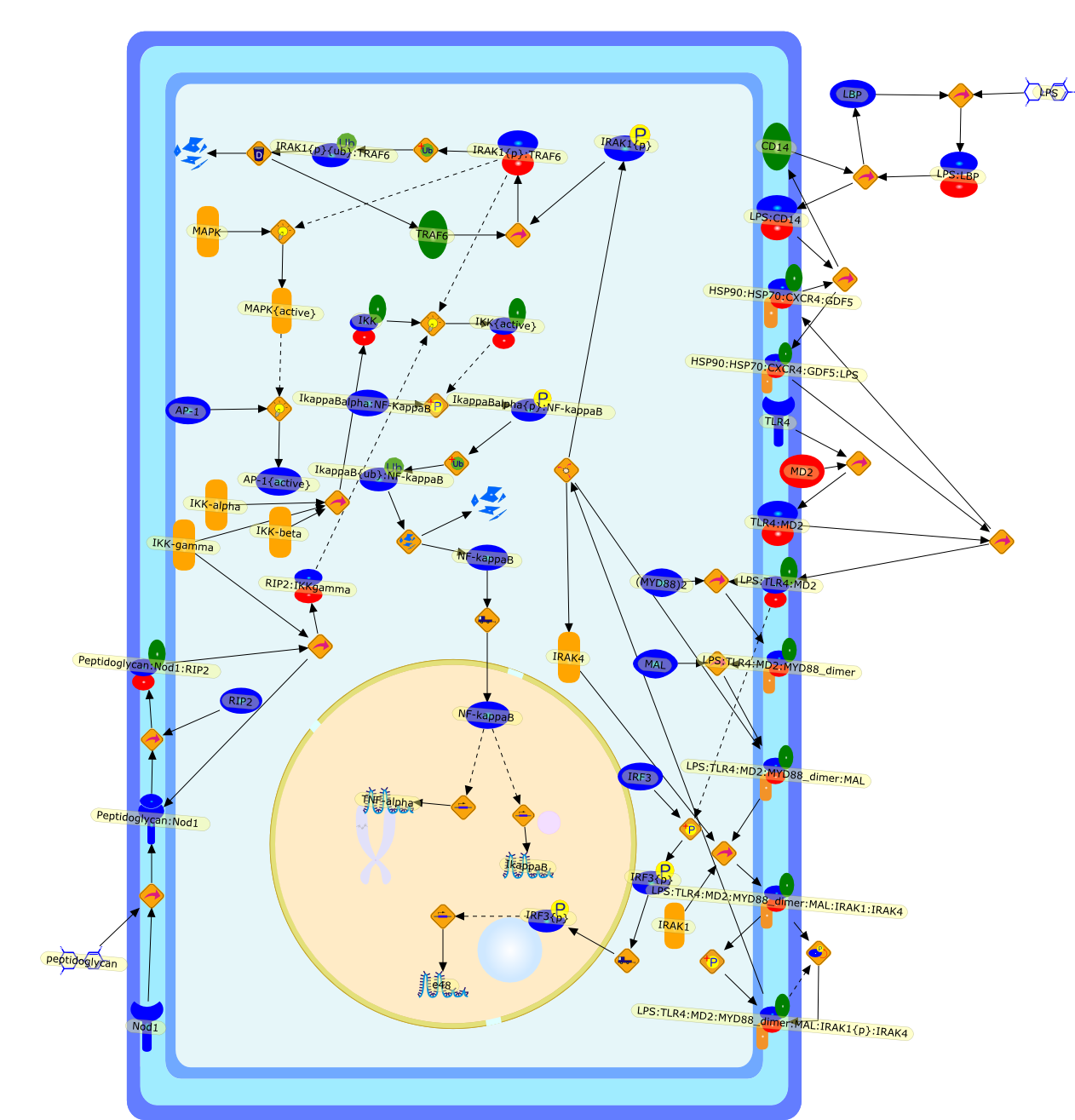
Bacterial lipopolysaccharide (LPS), the major structural component of the outerwall of Gram-negative bacteria, is a potent activator of macrophages. Activatedmacrophages produce a variety of inflammatory cytokines. Excessive production ofcytokines in response to LPS is regarded as the cause of septic shock. On theother hand, macrophages exposed to suboptimal doses of LPS are rendered tolerantto subsequent exposure to LPS and manifest a profoundly altered response to LPS.Increasing evidence suggests that monocytic cells from patients with sepsis andseptic shock survivors have characteristics of LPS tolerance. Thus, anunderstanding of the molecular mechanisms underlying activation and deactivationof macrophages in response to LPS is important for the development oftherapeutics for septic shock and the treatment of septic shock survivors. Overthe past several years, significant progress has been made in identifying andcharacterizing several key molecules and signal pathways involved in theregulation of macrophage functions by LPS. In this paper, we summarize thecurrent findings of the functions of the LPS receptor complex, which is composedof CD14, Toll-like receptor 4 (TLR4), and myeloid differentiation protein-2(MD-2), and the signal pathways of this LPS receptor complex with regard to bothactivation and deactivation of macrophages by LPS. In addition, recenttherapeutic approaches for septic shock targeting the LPS receptor complex aredescribed.
{kind=link}
{kind=link}
(MYD88)2,AP-1,AP-1{active},CD14,HSP90:HSP70:CXCR4:GDF5,HSP90:HSP70:CXCR4:GDF5:LPS,IKK,IKK-alpha,IKK-beta,IKK-gamma,IKK{active},IRAK1,IRAK1{p},IRAK1{p}:TRAF6,IRAK1{p}{ub}:TRAF6,IRAK4,IRF3,IRF3{p},IkappaB,IkappaBalpha:NF-KappaB,IkappaBalpha{p}:NF-kappaB,IkappaB{ub}:NF-kappaB,LBP,LPS,LPS:CD14,LPS:LBP,LPS:TLR4:MD2,LPS:TLR4:MD2:MYD88_dimer,LPS:TLR4:MD2:MYD88_dimer:MAL,LPS:TLR4:MD2:MYD88_dimer:MAL:IRAK1:IRAK4,LPS:TLR4:MD2:MYD88_dimer:MAL:IRAK1{p}:IRAK4,MAL,MAPK,MAPK{active},MD2,NF-kappaB,Nod1,Peptidoglycan:Nod1,Peptidoglycan:Nod1:RIP2,RIP2,RIP2:IKKgamma,TLR4,TLR4:MD2,TNF-alpha,TRAF6,e48,e66,peptidoglycan,protein remnants
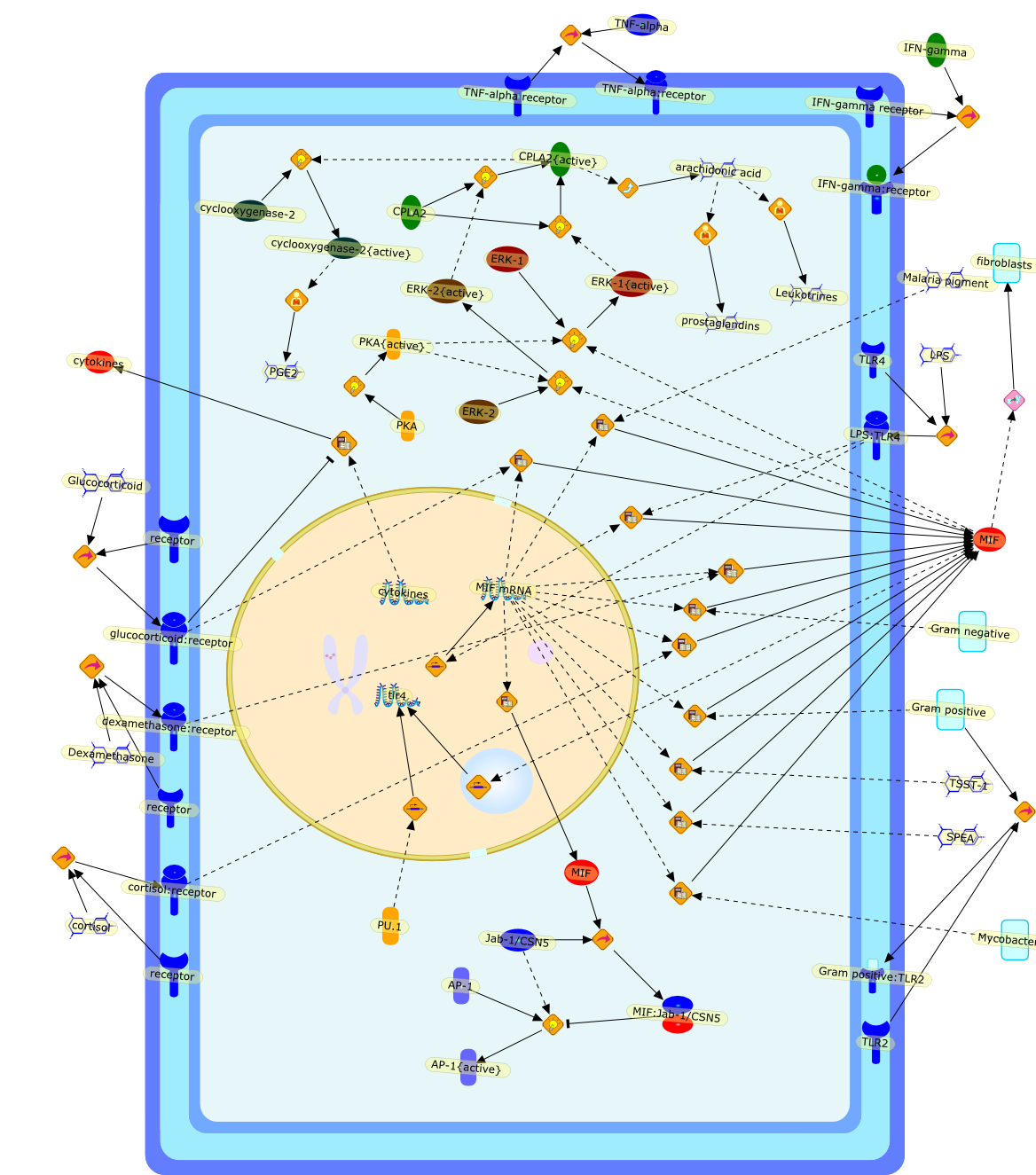
Among innate immune cells, macrophages play an essential role in the sensing andelimination of invasive microorganisms. Binding of microbial products topathogen-recognition receptors stimulates macrophages to release cytokines andother effector molecules that orchestrate the host innate and adaptive immuneresponses. Recently, the protein known as macrophage migration inhibitory factor(MIF) has emerged as a pivotal mediator of innate immunity. First identified asa T-cell cytokine, MIF was rediscovered as a protein released by pituitary cellsafter exposure to endotoxin [lipopolysaccharide (LPS)] or bacteria and inresponse to stress. Monocytes, macrophages and lymphocytes constitutivelyexpress MIF, which is rapidly released after stimulation with bacterialendotoxins and exotoxins, and cytokines. MIF induces powerful proinflammatorybiological responses and has been shown to be an important effector molecule ofseptic shock. High levels of MIF have been detected in the circulation ofpatients with severe sepsis and septic shock. Inhibition of MIF activity withneutralizing anti-MIF antibodies or deletion of the Mif gene led to a markedreduction in cytokine production and protected mice from lethal bacterial sepsisand toxic shock induced by Gram-negative endotoxin or Gram-positive exotoxins.Investigations into the mechanisms whereby MIF modulates innate immune responsesto endotoxin and Gram-negative bacteria have shown that MIF up-regulates theexpression of Toll-like receptor 4 (TLR4), the signal-transducing molecule ofthe LPS receptor complex. Thus, MIF enables cells, such as the macrophage, thatare at the forefront of the host antimicrobial defences, to sense promptly thepresence of invading Gram-negative bacteria and mount an innate immune response.Given that it is a pivotal regulator of innate immune responses to bacterialinfections, MIF appears to be a perfect target for novel therapeuticinterventions in patients with severe sepsis.
{kind=link}
{kind=link}
AP-1,AP-1{active},CPLA2,CPLA2{active},Dexamethasone,ERK-1,ERK-1{active},ERK-2,ERK-2{active},Glucocorticoid,Gram negative,Gram positive,Gram positive:TLR2,IFN-gamma,IFN-gamma receptor,IFN-gamma:receptor,Jab-1/CSN5,LPS,LPS:TLR4,Leukotrines,MIF,MIF mRNA,MIF:Jab-1/CSN5,Malaria pigment,Mycobacteria,PGE2,PKA,PKA{active},PU.1,SPEA,TLR2,TLR4,TNF-alpha,TNF-alpha receptor,TNF-alpha:receptor,TSST-1,arachidonic acid,cortisol,cortisol:receptor,cyclooxygenase-2,cyclooxygenase-2{active},cytokines,dexamethasone:receptor,fibroblasts,glucocorticoid:receptor,prostaglandins,receptor,tlr4
Recent advances in understanding the molecular basis for mammalian host immuneresponses to microbial invasion suggest that the first line of defense againstmicrobes is the recognition of pathogen-associated molecular patterns by a setof germline-encoded receptors: the Toll-like receptors (TLRs). TLRs have beenidentified as being part of a large family of pathogen-recognition receptorsthat play a decisive role in the induction of both innate and adaptive immunity.Indeed, activation of T lymphocytes depends on their interaction with dendriticcells previously stimulated by TLR agonists such as bacterial lipopolysaccharide(LPS), a TLR-4 ligand. A novel PKC epsilon (epsilon) was recently found to be acritical component of TLR-4 signaling pathway and thereby to play a key role inmacrophage and dendritic cell (DC) activation in response to LPS. Thus,controlling the kinase activity of PKC epsilon might represent an efficientstrategy to prevent or treat certain inflammatory disorders of microbial origin.
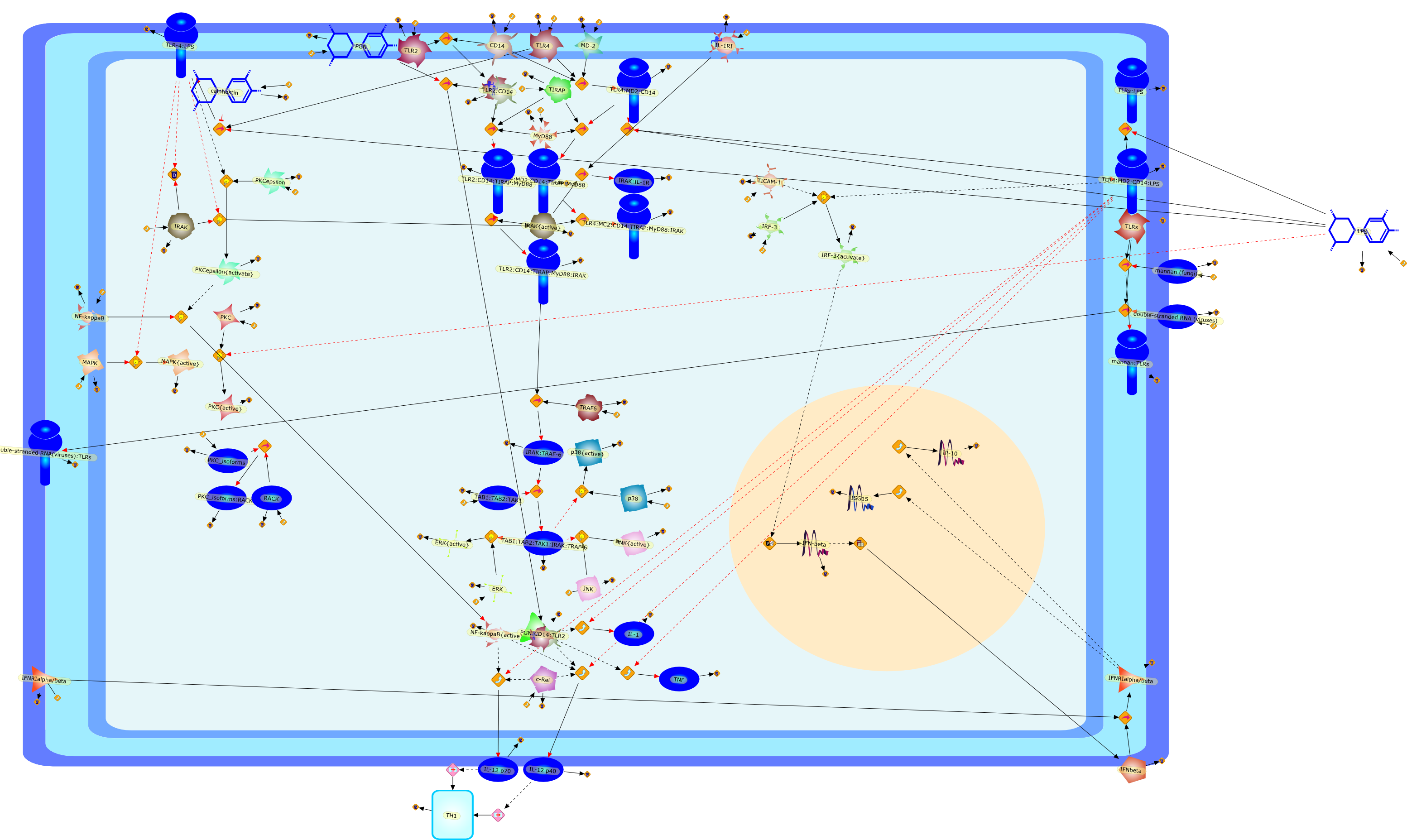{kind=link}
{kind=link}
CD14,ERK,ERK{active},IFN-beta,IFNRIalpha/beta,IFNbeta,IL-1,IL-12 p40,IL-12 p70,IL-1RI,IP-10,IRAK,IRAK:IL-1R,IRAK:TRAF-6,IRAK{active},IRF-3,IRF-3{activate},ISG15,JNK,JNK{active},LPS,MAPK,MAPK{active},MD-2,MyD88,NF-kappaB,NF-kappaB{active},PGN,PGN:CD14:TLR2,PKC,PKC_isoforms,PKC_isoforms:RACK,PKCepsilon,PKCepsilon{activate},PKC{active},RACK,TAB1:TAB2:TAK1,TAB1:TAB2:TAK1:IRAK:TRAF-6,TH1,TICAM-1,TIRAP,TLR-4:LPS,TLR2,TLR2:CD14,TLR2:CD14:TIRAP:MyD88,TLR2:CD14:TIRAP:MyD88:IRAK,TLR4,TLR4:MC2:CD14:TIRAP:MyD88:IRAK,TLR4:MD2:CD14,TLR4:MD2:CD14:LPS,TLR4:MD2:CD14:TIRAP:MyD88,TLRs,TLRs:LPS,TNF,TRAF6,c-Rel,calphostin,double-stranded RNA (viruses),double-stranded RNA(viruses):TLRs,mannan (fungi),mannan:TLRs,p38,p38{active}
Cytokine signaling is dependent on the activation of intracellular molecules,including JAKs (Janus family kinases) and STATs (signal transducers andactivators of transcription). Since the discovery of the SOCS (suppressor ofcytokine signaling) family molecules in 1997, extensive investigation of thesemolecules has given important insights into the as yet unclarified regulatorymechanisms of cytokine signaling. Recent gene targeting analyses of SOCSmolecules have revealed that they do indeed have essential roles in vivo (morespecific than those expected from overexpression studies in vitro) in thenegative regulation of various cytokines. Moreover, consistent with thepathological actions of cytokines in human diseases, recent lines of evidencealso indicate that SOCS molecules are implicated in autoimmunity, allergy andcancers.
{kind=link}
{kind=link}
CIS,CpGDNA,ERK,ERK1,ERK1{active},ERK2,ERK2{active},ERK{active},GH,GH:GHR,GH:GHR:CIS,GH:GHR:SOCS-2,GHR,IFN responsive genes,IGF-1,IGF-1:IGF-1R,IGF-1:IGF-1R:SOCS-2,IGF1,IL-2,IL-2:IL-2R,IL-2:IL-2R:CIS,IL-3:IL-3R,IL-3:IL-3R:CIS,IL-6,IRAK,IRS-1,IRS-1{active},IRS-4,Insulin:Insulin receptor,Insulin:Insulin receptor:SOCS6,JAK,LIgand:EpoR:CIS,LIgand:EpoR{ub}:CIS,LPS,Ligand,Ligand:EpoR,Ligand:EpoR:SOCS-3,Ligand:Prolactin,Ligand:Prolactin:CIS,Ligand:gp130,Ligand:gp130:SOCS-3,SOCS-1,SOCS-1:IRS-1,SOCS-1:IRS-1{ub},SOCS-1:JAK,SOCS-1:JAK{ub},SOCS-1:Vav,SOCS-1:Vav{ub},SOCS-1:iRAK,SOCS-2,SOCS-3,SOCS-3{p},SOCS-5,SOCS1,SOCS5,SOCS6,SOCS6:IRS-4,STAT1,STAT1{active},STAT5,STAT5B,STAT5B{active},STAT5{active},Vav,degradants,insulin,protein kinase B,protein kinase B {active}
Macrophages play diverse roles in host defense and in maintenance ofhomeostasis. Based on their ability to promote inflammatory responses,inappropriate macrophage function also contributes to numerous pathologicalprocesses, including atherosclerosis, rheumatoid arthritis and inflammatorybowel disease. Members of the nuclear receptor superfamily of ligand-dependenttranscriptions factors have emerged as key regulators of inflammation and lipidhomeostasis in macrophages. These include the glucocorticoid receptor (GR),which inhibits inflammatory programs of gene expression in response to naturalcorticosteroids and synthetic anti-inflammatory ligands such as dexamethasone.Also, in response to endogenous eicosanoids and oxysterols, respectively,peroxisome proliferator-activated receptors (PPARs) and liver X receptors (LXRs)regulate transcriptional programs involved in inflammatory responses and lipidhomeostasis. Identification of their mechanisms of action should help guide thedevelopment of new therapeutic agents useful in the treatment of diseases inwhich macrophages play critical pathogenic roles.
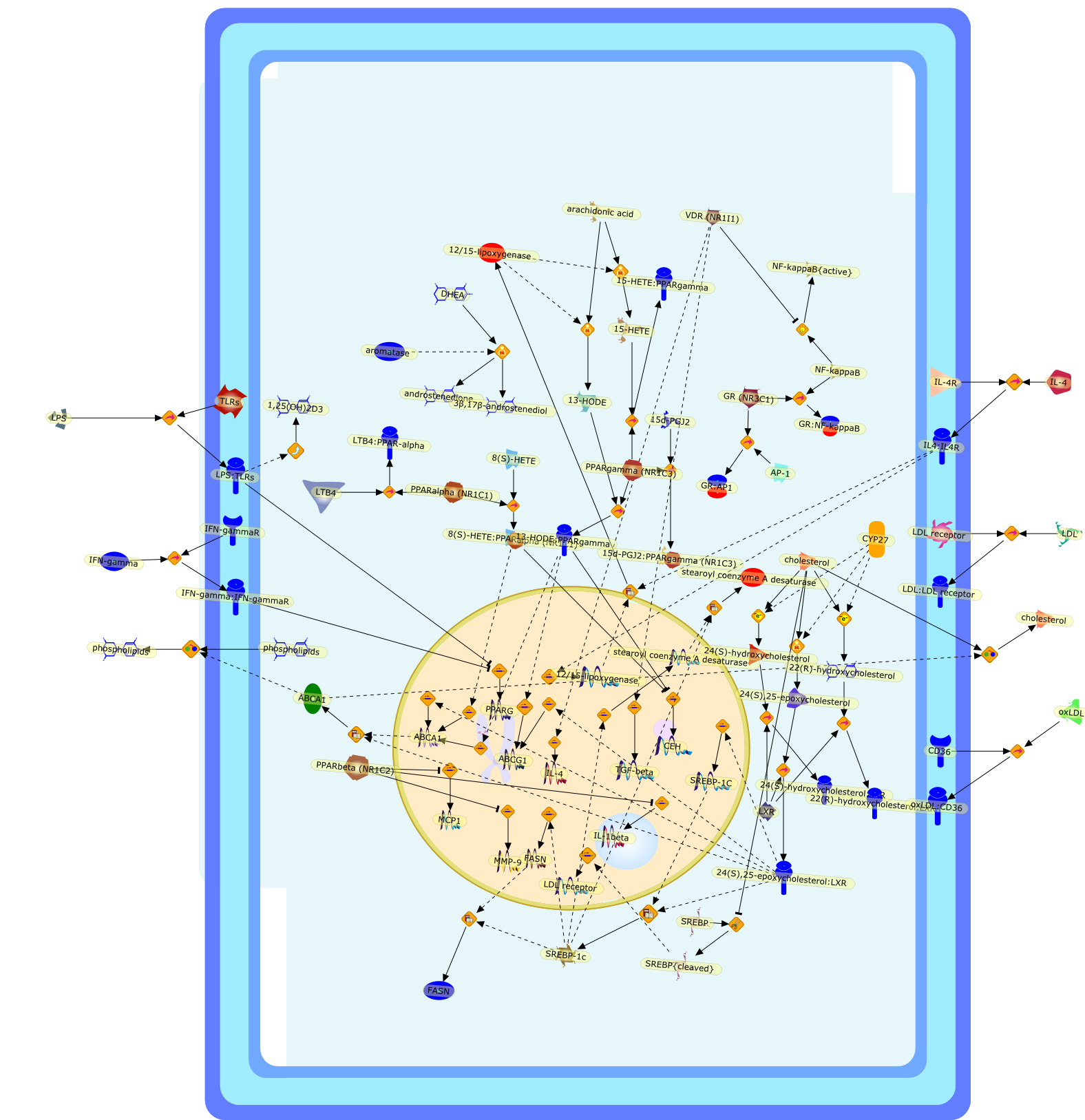{kind=link}
{kind=link}
1,25(OH)2D3,12/15-lipoxygenase,13-HODE,13-HODE:PPARgamma,15-HETE,15-HETE:PPARgamma,15d-PGJ2,15d-PGJ2:PPARgamma (NR1C3),22(R)-hydroxycholesterol,22(R)-hydroxycholesterol:LXR,24(S),25-epoxycholesterol,24(S),25-epoxycholesterol:LXR,24(S)-hydroxycholesterol,24(S)-hydroxycholesterol:LXR,3β,17β-androstenediol,8(S)-HETE,8(S)-HETE:PPARalpha (NR1C1),ABCA1,ABCG1,AP-1,CD36,CEH,CYP27,DHEA,FASN,GR (NR3C1),GR:AP1,GR:NF-kappaB,IFN-gamma,IFN-gamma:IFN-gammaR,IFN-gammaR,IL-1beta,IL-4,IL-4R,IL4:IL4R,LDL,LDL receptor,LDL:LDL receptor,LPS,LPS:TLRs,LTB4,LTB4:PPAR-alpha,LXR,MCP1,MMP-9,NF-kappaB,NF-kappaB{active},PPARG,PPARalpha (NR1C1),PPARbeta (NR1C2),PPARgamma (NR1C3),SREBP,SREBP-1C,SREBP-1c,SREBP{cleaved},TGF-beta,TLRs,VDR (NR1I1),androstenedione,arachidonic acid,aromatase,cholesterol,oxLDL,oxLDL:CD36,phospholipids,stearoyl coenzyme A desaturase
The concept that Toll-like receptors (TLRs) recognize specific molecularpatterns in various pathogens has been established. In signal transduction viaTLRs, MyD88, which harbors a Toll/IL-1 receptor (TIR)-domain and a death domain,has been shown to link between TLRs and MyD88-dependent downstream eventsleading to proinflammatory cytokine production and splenocyte proliferation.However, recent studies using MyD88-deficient mice have revealed that some TLRspossess a MyD88-independent pathway, which is represented by interferon(IFN)-beta production induced by LPS stimulation. This indicates that additionalsignaling molecules other than MyD88 exist in the TLR signaling pathway. Indeed,two additional TIR domain-containing adaptors, TIRAP/Mal and TRIF, have recentlybeen identified. Both define the specific biological responses of each TLR.
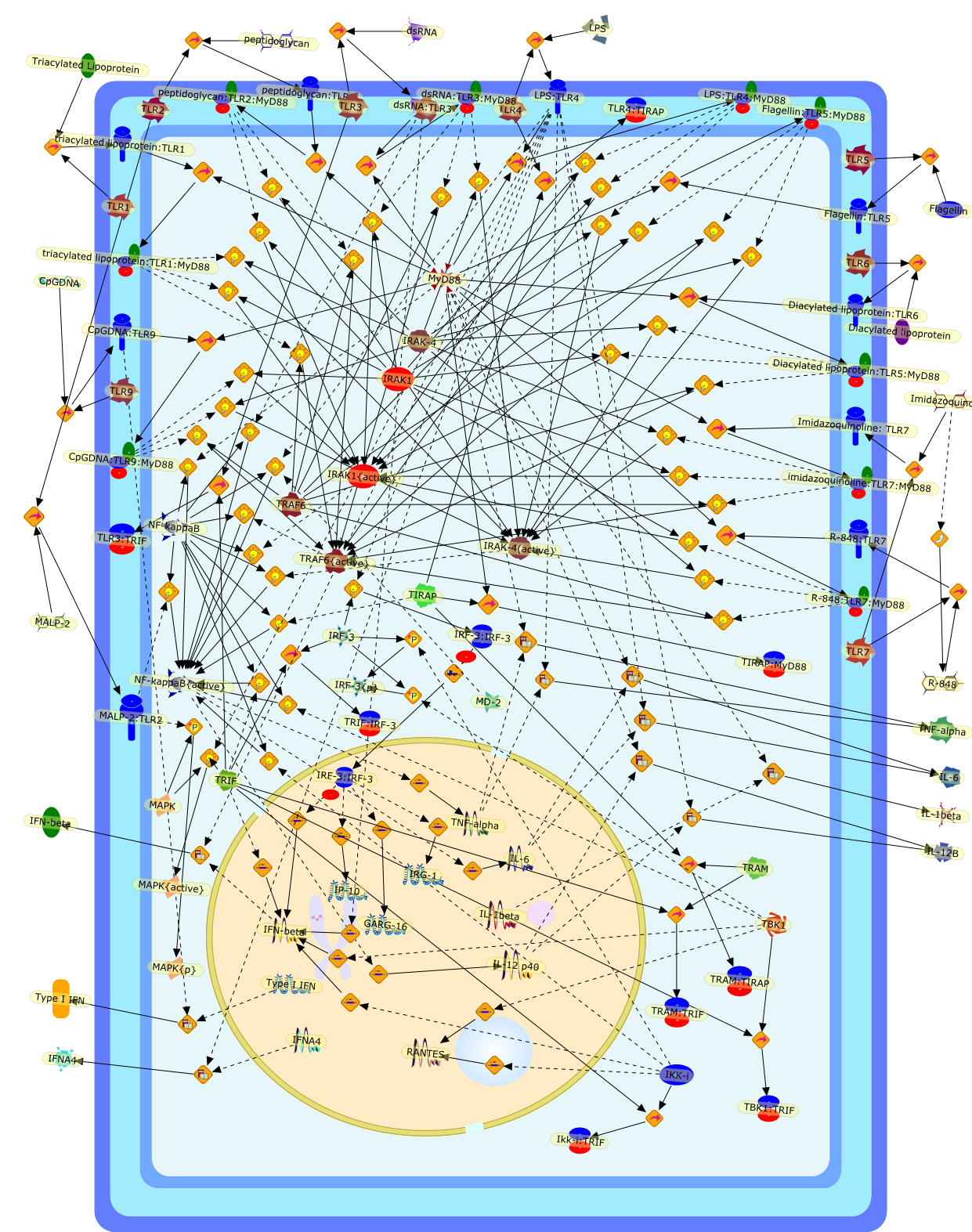{kind=link}
{kind=link}
CpGDNA,CpGDNA:TLR9,CpGDNA:TLR9:MyD88,Diacylated lipoprotein,Diacylated lipoprotein:TLR5:MyD88,Diacylated lipoprotein:TLR6,Flagellin,Flagellin:TLR5,Flagellin:TLR5:MyD88,GARG-16,IFN-beta,IFNA4,IKK-i,IL-12 p40,IL-12B,IL-1beta,IL-6,IP-10,IRAK-4,IRAK-4(active},IRAK1,IRAK1{active},IRF-3,IRF-3:IRF-3,IRF-3{p},IRG-1,Ikk-i:TRIF,Imidazoquinoline,Imidazoquinoline: TLR7,LPS,LPS:TLR4,LPS:TLR4:MyD88,MALP-2,MALP-2:TLR2,MAPK,MAPK{active},MAPK{p},MD-2,MyD88,NF-kappaB,NF-kappaB{active},R-848,R-848:TLR7,R-848:TLR7:MyD88,RANTES,TBK1,TBK1:TRIF,TIRAP,TIRAP:MyD88,TLR1,TLR2,TLR3,TLR3:TRIF,TLR4,TLR4:TIRAP,TLR5,TLR6,TLR7,TLR9,TNF-alpha,TRAF6,TRAF6{active},TRAM,TRAM:TIRAP,TRAM:TRIF,TRIF,TRIF:IRF-3,Triacylated Lipoprotein,Type I IFN,dsRNA,dsRNA:TLR3,dsRNA:TLR3:MyD88,imidazoquinoline:TLR7:MyD88,peptidoglycan,peptidoglycan:TLR2,peptidoglycan:TLR2:MyD88,triacylated lipoprotein:TLR1,triacylated lipoprotein:TLR1:MyD88
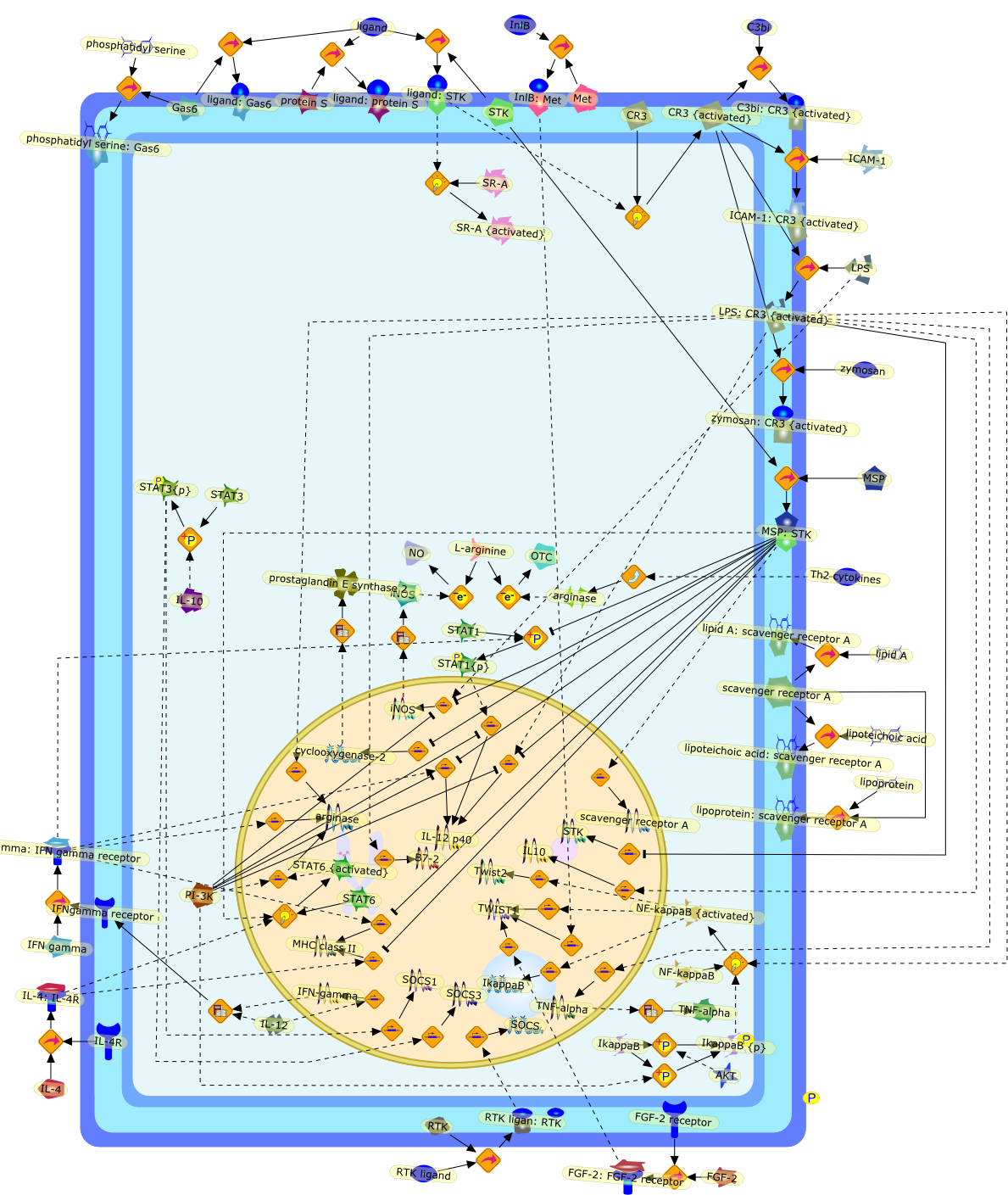{kind=link}
{kind=link}
AKT,B7-2,C3bi,C3bi: CR3 {activated},CR3,CR3 {activated},FGF-2,FGF-2 receptor,FGF-2: FGF-2 receptor,Gas6,ICAM-1,ICAM-1: CR3 {activated},IFN gamma,IFN-gamma,IFNgamma receptor,IFNgamma: IFN gamma receptor,IL-10,IL-12,IL-12 p40,IL-4,IL-4: IL-4R,IL-4R,IL10,IkappaB,IkappaB {p},InlB,InlB: Met,L-arginine,LPS,LPS: CR3 {activated},MHC class II,MSP,MSP: STK,Met,NF-kappaB,NF-kappaB {activated},NO,OTC,PI-3K,RTK,RTK ligan: RTK,RTK ligand,SOCS,SOCS1,SOCS3,SR-A,SR-A {activated},STAT1,STAT1{p},STAT3,STAT3{p},STAT6,STAT6 {activated},STK,TNF-alpha,TWIST1,Th2 cytokines,Twist2,arginase,cyclooxygenase-2,iNOS,ligand,ligand: Gas6,ligand: STK,ligand: protein S,lipid A,lipid A: scavenger receptor A,lipoprotein,lipoprotein: scavenger receptor A,lipoteichoic acid,lipoteichoic acid: scavenger receptor A,phosphatidyl serine,phosphatidyl serine: Gas6,prostaglandin E synthase 2,protein S,scavenger receptor A,zymosan,zymosan: CR3 {activated}
Synthetic oligodeoxynucleotides (ODN) expressing non-methylated "CpG motifs"patterned after those present in bacterial DNA have characteristicimmunomodulatory effects. CpG DNA is recognized as a pathogen-associatedmolecular pattern, and triggers a rapid innate immune response. CpG ODN arebeing harnessed for a variety of therapeutic uses, including as immuneadjuvants, for cancer therapy, as anti-allergens, and as immunoprotectiveagents. The signal transduction pathway mediated by the engagement of CpG DNAwith Toll-like receptor 9 (TLR9) is shared with other members of the TLR family.Recent studies demonstrate that formation and maturation of CpG DNA-containingendosomes are regulated by phosphatidylinositol 3 kinases and the Ras-associatedGTP-binding protein, Rab5, which are essential for the initiation ofTLR9-mediated signaling.
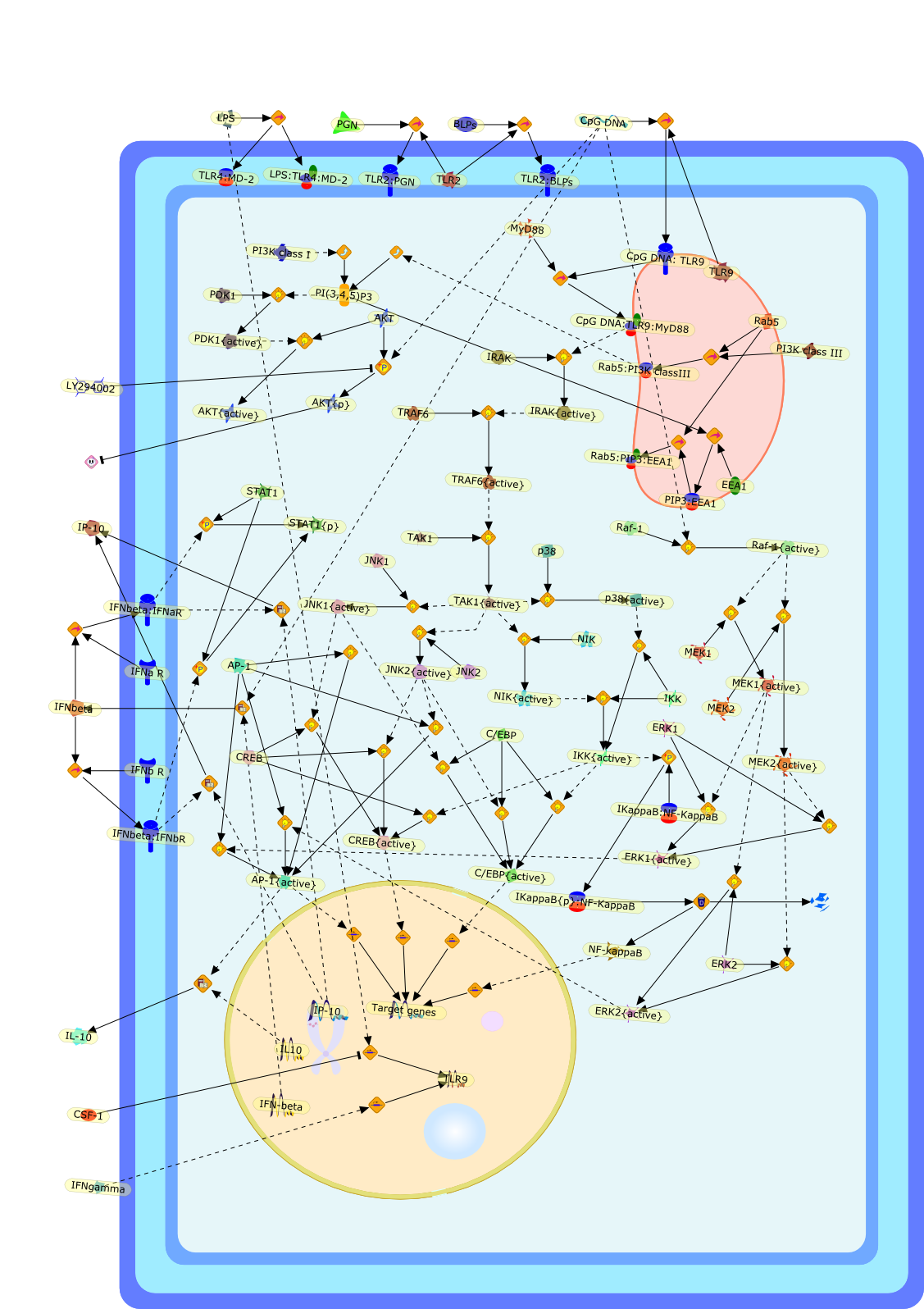{kind=link}
{kind=link}
AKT,AKT{active},AKT{p},AP-1,AP-1{active},BLPs,C/EBP,C/EBP{active},CREB,CREB{active},CSF-1,CpG DNA,CpG DNA: TLR9,CpG DNA:TLR9:MyD88,EEA1,ERK1,ERK1{active},ERK2,ERK2{active},IFN-beta,IFNa R,IFNb R,IFNbeta,IFNbeta:IFNaR,IFNbeta:IFNbR,IFNgamma,IKK,IKK{active},IKappaB:NF-KappaB,IKappaB{p}:NF-KappaB,IL-10,IL10,IP-10,IRAK,IRAK{active},JNK1,JNK1{active},JNK2,JNK2{active},LPS,LPS:TLR4:MD-2,LY294002,MEK1,MEK1{active},MEK2,MEK2{active},MyD88,NF-kappaB,NIK,NIK{active},PDK1,PDK1{active},PGN,PI(3,4,5)P3,PI3K class I,PI3K class III,PIP3:EEA1,Rab5,Rab5:PI3K classIII,Rab5:PIP3:EEA1,Raf-1,Raf-1{active},STAT1,STAT1{p},TAK1,TAK1{active},TLR2,TLR2:BLPs,TLR2:PGN,TLR4:MD-2,TLR9,TRAF6,TRAF6{active},Target genes,degradants,p38,p38{active}
Many of the stromal-derived signals and factors that regulate B lymphopoiesishave been identified. We review recent evidence from our laboratory that showsthat there are at least three phases during B-cell development when cells directtheir own maturation, independent of stromal cells. Following the expression ofthe preB-cell receptor (preBCR), cells acquire the ability to proliferate in lowlevels of interleukin-7 (IL-7), which acts as a self-selecting mechanism toexpand cells that have successfully expressed a preBCR in environments that arenon-permissive to preBCR- cells. Second, the preBCR is required for acontact-mediated event between B-cell progenitors. Disruption at this stageprevents the further maturation of progenitors to the lipopolysaccharide(LPS)-responsive stage. Finally, the transition from IL-7 receptor to matureantigen receptor-based signaling is enhanced by a novel member of the tachykininfamily, hemokinin-1. This series of maturation, survival, and differentiationsignals is generated by B-lineage cells as they progress through developmentalcheckpoints on the way to becoming functionally mature cells.
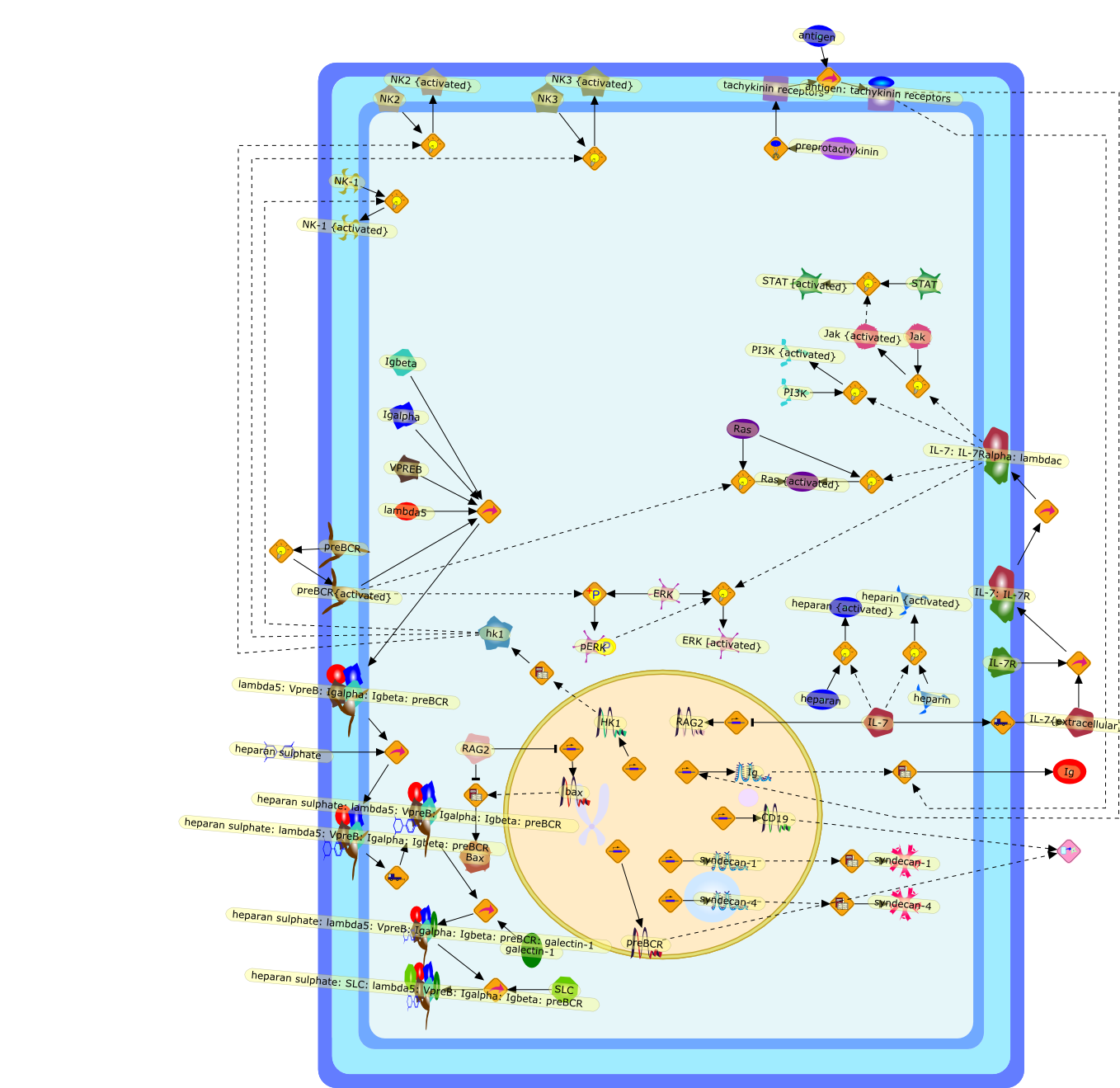{kind=link}
{kind=link}
Bax,CD19,ERK,ERK [activated},HK1,IL-7,IL-7: IL-7R,IL-7: IL-7Ralpha: lambdac,IL-7R,IL-7{extracellular},Ig,Igalpha,Igbeta,Jak,Jak {activated},NK-1,NK-1 {activated},NK2,NK2 {activated},NK3,NK3 {activated},PI3K,PI3K {activated},RAG2,Ras,Ras {activated},SLC,STAT,STAT [activated},VPREB,antigen,antigen: tachykinin receptors,bax,galectin-1,heparan,heparan sulphate,heparan sulphate: SLC: lambda5: VpreB: Igalpha: Igbeta: preBCR,heparan sulphate: lambda5: VpreB: Igalpha: Igbeta: preBCR,heparan sulphate: lambda5: VpreB: Igalpha: Igbeta: preBCR: galectin-1,heparan {activated},heparin,heparin {activated},hk1,lambda5,lambda5: VpreB: Igalpha: Igbeta: preBCR,pERK,preBCR,preBCR{activated},preprotachykinin,syndecan-1,syndecan-4,tachykinin receptors
Production of type I interferon (IFN-alpha/beta) by virus-infected cells is thecentral event in their antiviral immune responses. In mammalian cells,IFN-alpha/beta gene transcription is induced through distinct signaling pathwaysby viral infection or by treatment with double-stranded (ds) RNA, which is anintermediate of virus replication. Toll-like receptor 3 (TLR3) was found torecognize dsRNA and transmit signals to activate NF-kappaB and the IFN-betapromoter. Recent identification of the TLR3-adaptor protein and its downstreamsignaling molecules, which are involved in IFN-alpha/beta production, revealed anovel IFN-inducing pathway for an anti-viral immune response. Here, we summarizethe current knowledge of TLR3-mediated immune responses.
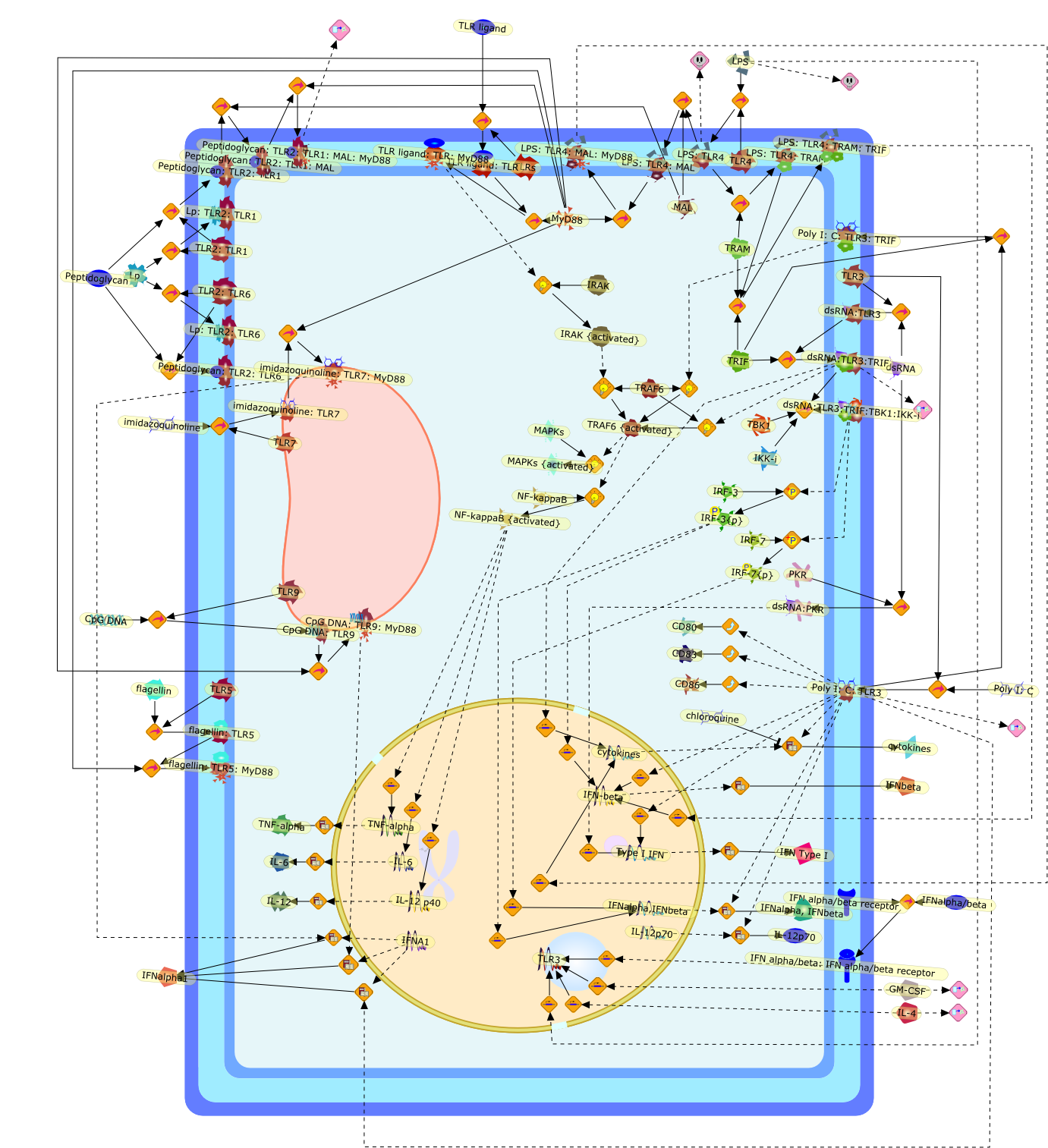{kind=link}
{kind=link}
CD80,CD83,CD86,CpG DNA,CpG DNA: TLR9,CpG DNA: TLR9: MyD88,GM-CSF,IFN Type I,IFN alpha/beta receptor,IFN alpha/beta: IFN alpha/beta receptor,IFN-beta,IFNA1,IFNalpha, IFNbeta,IFNalpha,IFNbeta,IFNalpha/beta,IFNalpha1,IFNbeta,IKK-i,IL-12,IL-12 p40,IL-12p70,IL-4,IL-6,IRAK,IRAK {activated},IRF-3,IRF-3{p},IRF-7,IRF-7{p},LPS,LPS: TLR4,LPS: TLR4: MAL,LPS: TLR4: MAL: MyD88,LPS: TLR4: TRAM,LPS: TLR4: TRAM: TRIF,Lp,Lp: TLR2: TLR1,Lp: TLR2: TLR6,MAL,MAPKs,MAPKs {activated},MyD88,NF-kappaB,NF-kappaB {activated},PKR,Peptidoglycan,Peptidoglycan: TLR2: TLR1,Peptidoglycan: TLR2: TLR1: MAL,Peptidoglycan: TLR2: TLR1: MAL: MyD88,Peptidoglycan: TLR2: TLR6,Poly I: C,Poly I: C: TLR3,Poly I: C: TLR3: TRIF,TBK1,TLR ligand,TLR ligand: TLR,TLR ligand: TLR: MyD88,TLR2: TLR1,TLR2: TLR6,TLR3,TLR4,TLR5,TLR7,TLR9,TLRs,TNF-alpha,TRAF6,TRAF6 {activated},TRAM,TRIF,Type I IFN,chloroquine,cytokines,dsRNA,dsRNA:PKR,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:TBK1:IKK-i,flagellin,flagellin: TLR5,flagellin: TLR5: MyD88,imidazoquinoline,imidazoquinoline: TLR7,imidazoquinoline: TLR7: MyD88
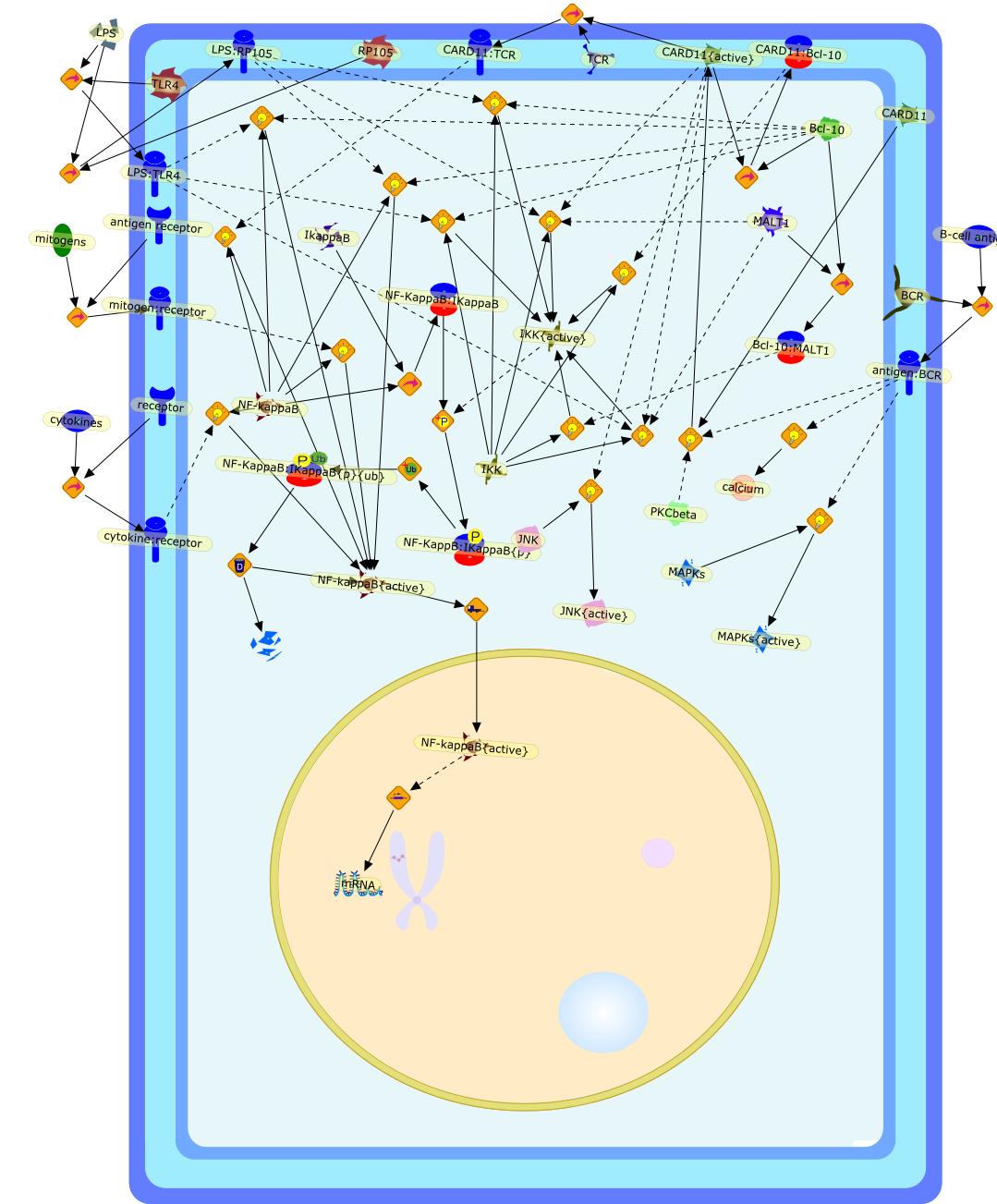{kind=link}
{kind=link}
B-cell antigen,BCR,Bcl-10,Bcl-10:MALT1,CARD11,CARD11:Bcl-10,CARD11:TCR,CARD11{active},IKK,IKK{active},IkappaB,JNK,JNK{active},LPS,LPS:RP105,LPS:TLR4,MALT1,MAPKs,MAPKs{active},NF-KappB:IKappaB{p},NF-KappaB:IKappaB,NF-KappaB:IKappaB{p}{ub},NF-kappaB,NF-kappaB{active},PKCbeta,RP105,TCR,TLR4,antigen receptor,antigen:BCR,calcium,cytokine:receptor,cytokines,mRNA,mitogen:receptor,mitogens,proteosome remnant,receptor
Toll-like receptors (TLRs) are pathogen recognition molecules that activate theimmune system as part of the innate immune response. Microbial recognition byTLRs plays a crucial role in the host immune system's decision to respond or notto a particular microbial infection. Lipopolysaccharide (LPS), a membraneglycolipid of Gram-negative bacteria, exhibits strong immunostimulating activityamong TLR ligands and has been studied in great detail. Recent studies haveshown that cell surface TLR4-MD-2 physically interacts with LPS and triggers therelease of an LPS signal, revealing a host-pathogen interaction mediated by TLR.
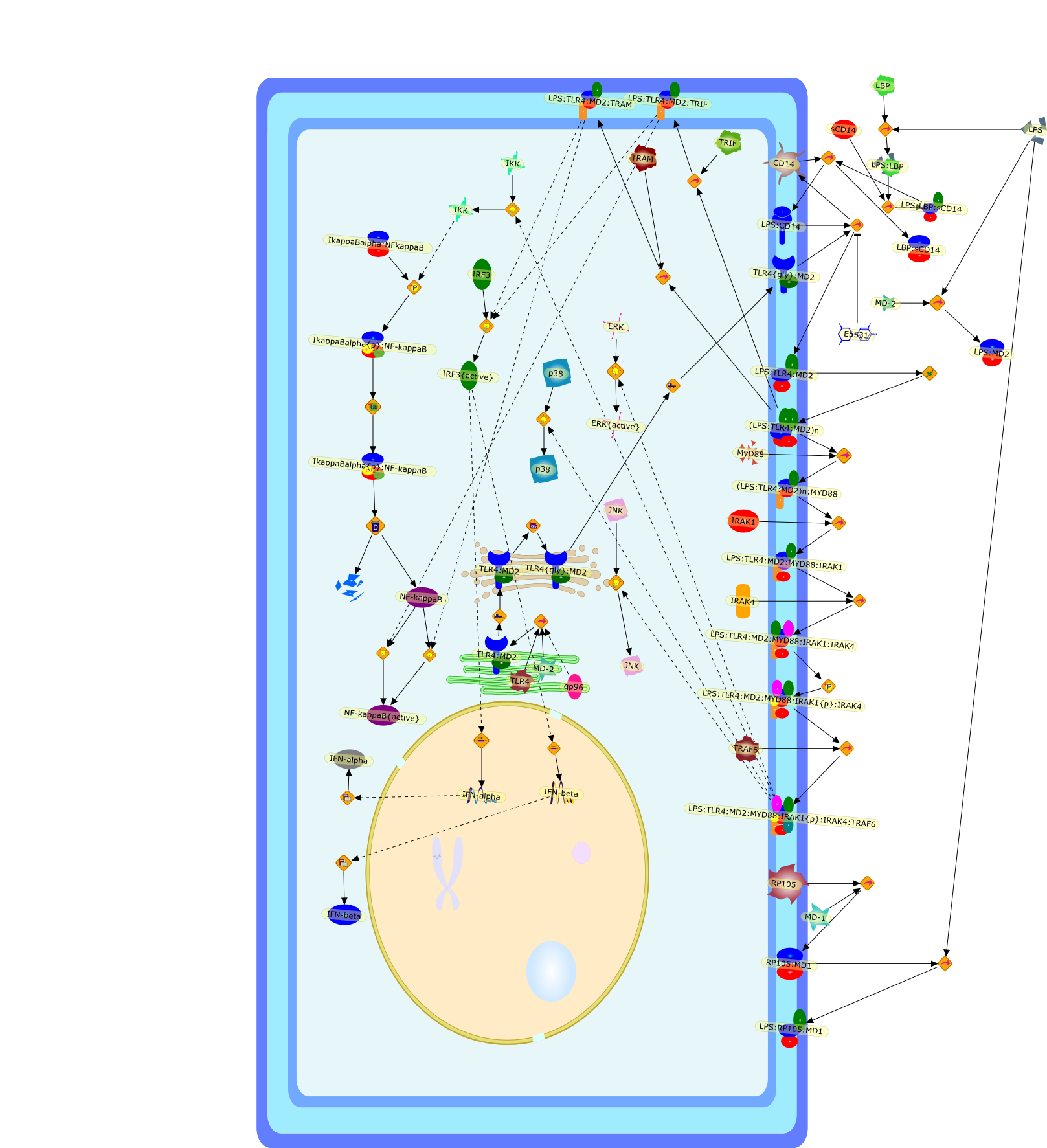{kind=link}
{kind=link}
(LPS:TLR4:MD2)n,(LPS:TLR4:MD2)n:MYD88,CD14,E5531,ERK,ERK{active},IFN-alpha,IFN-beta,IKK,IRAK1,IRAK4,IRF3,IRF3{active},IkappaBalpha:NFkappaB,IkappaBalpha{p}:NF-kappaB,JNK,LBP,LBP:sCD14,LPS,LPS:CD14,LPS:LBP,LPS:LBP:sCD14,LPS:MD2,LPS:RP105:MD1,LPS:TLR4:MD2,LPS:TLR4:MD2:MYD88:IRAK1,LPS:TLR4:MD2:MYD88:IRAK1:IRAK4,LPS:TLR4:MD2:MYD88:IRAK1{p}:IRAK4,LPS:TLR4:MD2:MYD88:IRAK1{p}:IRAK4:TRAF6,LPS:TLR4:MD2:TRAM,LPS:TLR4:MD2:TRIF,MD-1,MD-2,MyD88,NF-kappaB,NF-kappaB{active},RP105,RP105:MD1,TLR4,TLR4:MD2,TLR4{gly}:MD2,TRAF6,TRAM,TRIF,gp96,p38,protein remnants,sCD14
Humans are constantly exposed to a wide variety of microorganisms that can causeinfection. In self-defense, the human host has evolved complex protectivemechanisms, and Toll-like receptors (TLRs) have emerged as a central point indefense. These receptors bind molecular structures that are expressed bymicrobes but are not expressed by the human host, eg, lipopolysaccharides (LPS)or double-stranded RNA (dsRNA). Activation of these receptors initiates aninflammatory cascade that attempts to clear the offending pathogen and set inmotion a specific adaptive immune response. Defects in sensing of pathogens maypredispose the host to recurrent infections. The relative rarity of thesesyndromes of defective innate immunity, however, speaks to the redundancy insensing of pathogens by the innate immune system. More common, polymorphisms inTLR4 are associated with increased predisposition to severe and recurrentinfections but protection against atherosclerotic disease due to diminishedinflammation. Toll-like receptor signaling may also contribute to thepathophysiology of disease and injure the host by activating a deleteriousimmune response such as in sepsis or inflammatory bowel disease (IBD). The focusof this article is to describe the role of TLRs in the innate immune response inhealth and disease.
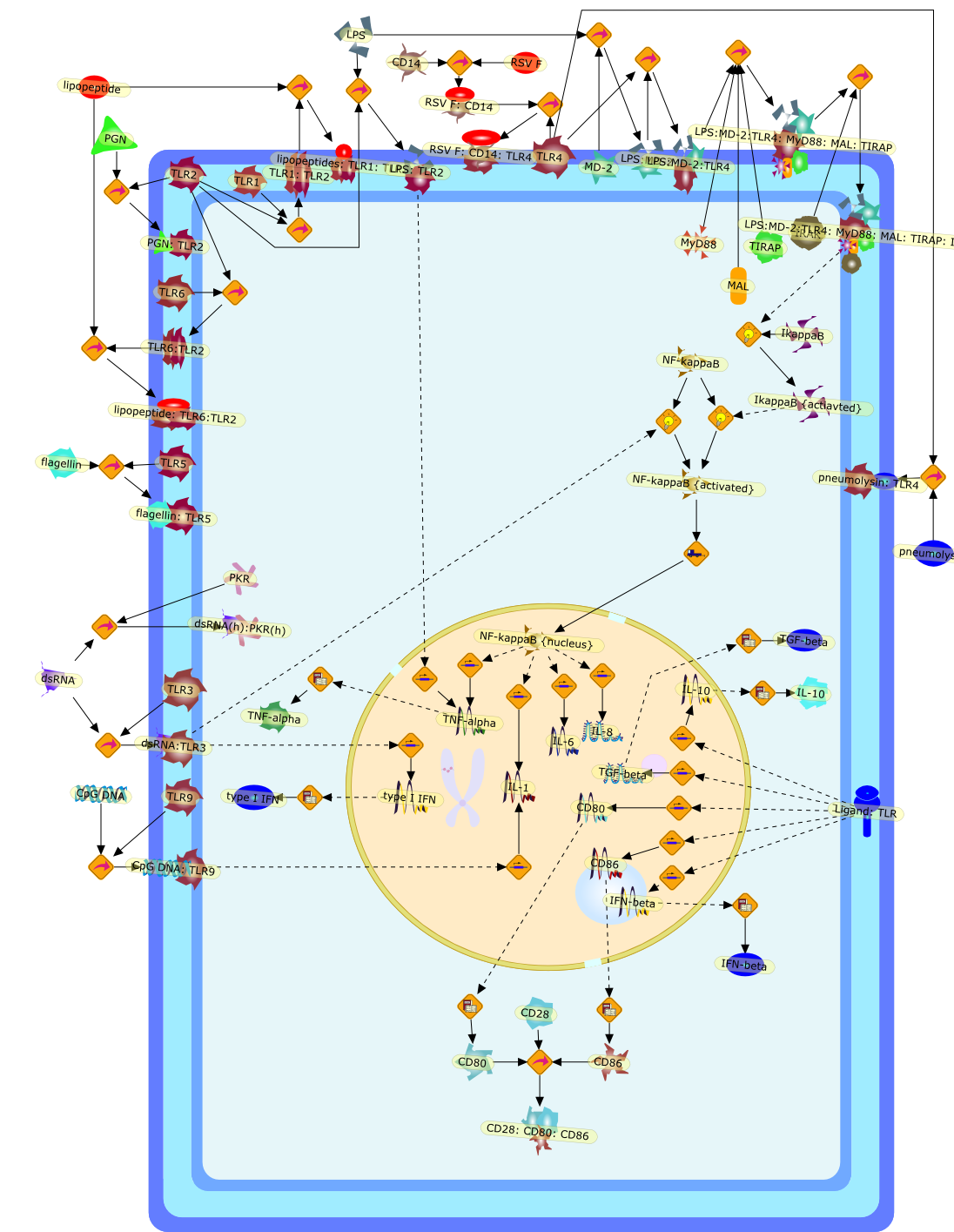{kind=link}
{kind=link}
CD14,CD28,CD28: CD80: CD86,CD80,CD86,CpG DNA,CpG DNA: TLR9,IFN-beta,IL-1,IL-10,IL-6,IL-8,IRAK,IkappaB,IkappaB {actiavted},LPS,LPS: TLR2,LPS:MD-2,LPS:MD-2:TLR4,LPS:MD-2:TLR4: MyD88: MAL: TIRAP,LPS:MD-2:TLR4: MyD88: MAL: TIRAP: IRAK,Ligand: TLR,MAL,MD-2,MyD88,NF-kappaB,NF-kappaB {activated},NF-kappaB {nucleus},PGN,PGN: TLR2,PKR,RSV F,RSV F: CD14,RSV F: CD14: TLR4,TGF-beta,TIRAP,TLR1,TLR1: TLR2,TLR2,TLR3,TLR4,TLR5,TLR6,TLR6:TLR2,TLR9,TNF-alpha,dsRNA,dsRNA(h):PKR(h),dsRNA:TLR3,flagellin,flagellin: TLR5,lipopeptide,lipopeptide: TLR6:TLR2,lipopeptides: TLR1: TLR2,pneumolysin,pneumolysin: TLR4,type I IFN
Post-transcriptional mechanisms play a critical role in regulating theexpression of numerous proteins that promote inflammatory arthritis. The mRNAsencoding a subset of these proteins possess adenine/uridine-rich elements (AREs)in their 3'-untranslated regions that profoundly influence the rate at whichmRNA is degraded and translated into protein. Tristetraprolin (TTP) and T cellintracellular antigen-1 (TIA-1) are ARE-binding proteins that dampen theexpression of this class of proteins by promoting mRNA degradation and proteintranslation, respectively. We have discovered that TIA-1 and TTP function asarthritis-suppressor genes: TIA-1-/- mice develop mild arthritis, TTP-/- micedevelop severe arthritis, and TIA-1-/-TTP-/- mice develop very severe arthritis.Paradoxically, lipopolysaccharide (LPS)-activated macrophages derived fromTIA-1-/-TTP-/- macrophages produce less tumor necrosis factor alpha (TNF-alpha)than TIA-1-/- or TTP-/- macrophages. The bone marrows of these mice exhibitincreased cellularity, reflecting the presence of mature neutrophils thatsecrete TNF-alpha in response to LPS stimulation. We hypothesize thatTIA-1-/-TTP-/- neutrophils are a source of arthritigenic TNF-alpha, whichpromotes severe erosive arthritis in these mice.
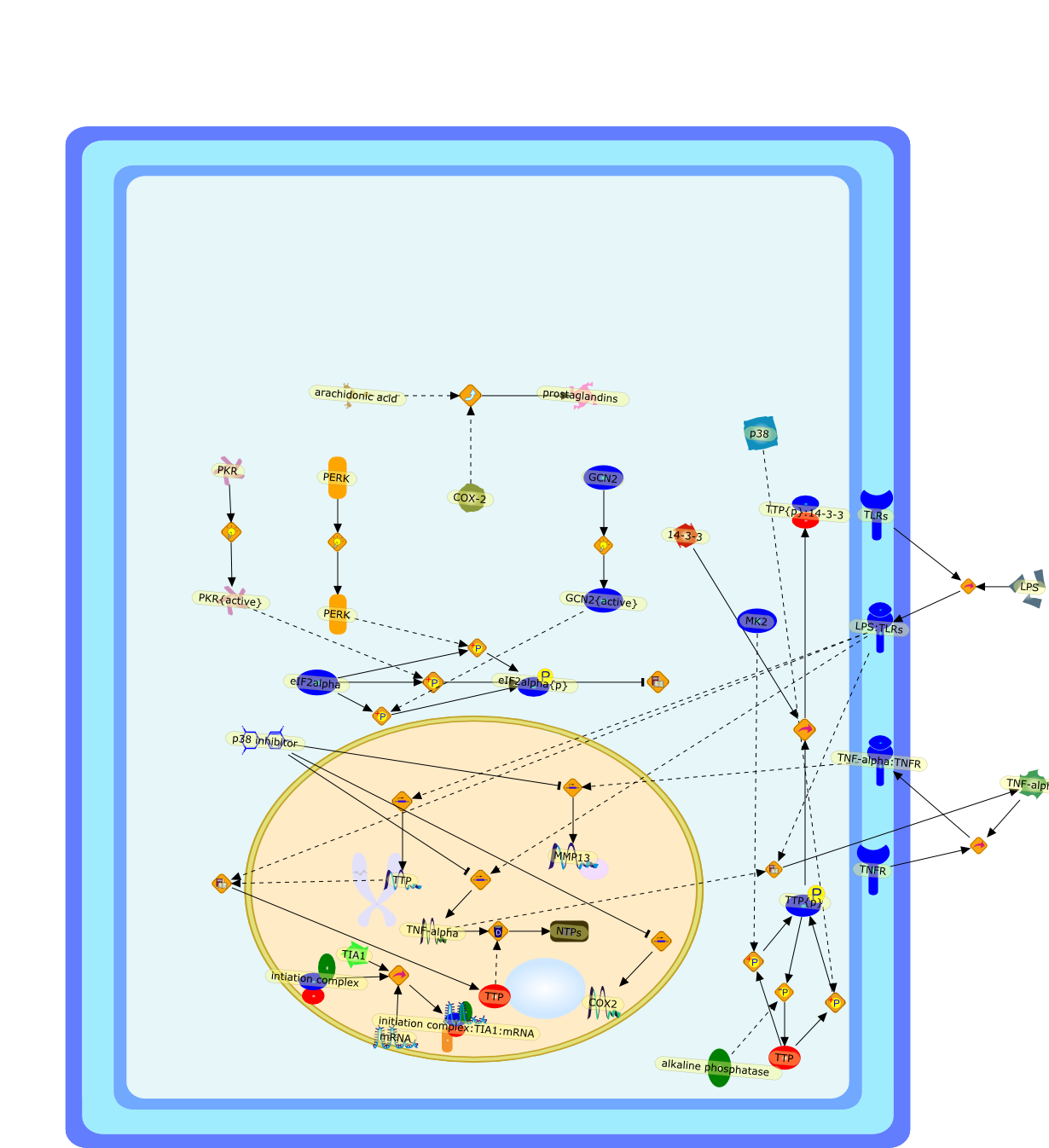{kind=link}
{kind=link}
14-3-3,COX-2,COX2,GCN2,GCN2{active},LPS,LPS:TLRs,MK2,MMP13,NTPs,PERK,PKR,PKR{active},TIA1,TLRs,TNF-alpha,TNF-alpha:TNFR,TNFR,TTP,TTP{p},TTP{p}:14-3-3,alkaline phosphatase,arachidonic acid,eIF2alpha,eIF2alpha{p},initiation complex:TIA1:mRNA,intiation complex,mRNA,p38,p38 inhibitor,prostaglandins
Toll-like receptors (TLRs) have been identified as a major class ofpattern-recognition receptors. Recognition of pathogen-associated molecularpatterns (PAMPs) by TLRs, alone or in heterodimerization with other TLR ornon-TLR receptors, induces signals responsible for the activation of genesimportant for an effective host defense, especially proinflammatory cytokines.Although a certain degree of redundancy exists between signals induced by thevarious TLRs, recent studies have identified intracellular pathways specific forindividual TLRs. This leads to the release of cytokine profiles specific forparticular PAMPs, and thus, TLRs confer a certain degree of specificity to theinnate-immune response. In addition to the activation of the innate-immuneresponse, TLR-mediated recognition represents a link between the innate- andacquired-immune systems, by inducing the maturation of dendritic cells anddirecting the T helper responses. Alternatively, recent data have also suggestedTLR-mediated escape mechanisms used by certain pathogenic microorganisms,especially through TLR2 induction of anti-inflammatory cytokines. Finally, thecrucial role of TLRs for the host defense against infections has beenstrengthened recently by the description of patients partially defective in theTLR-activation pathways.
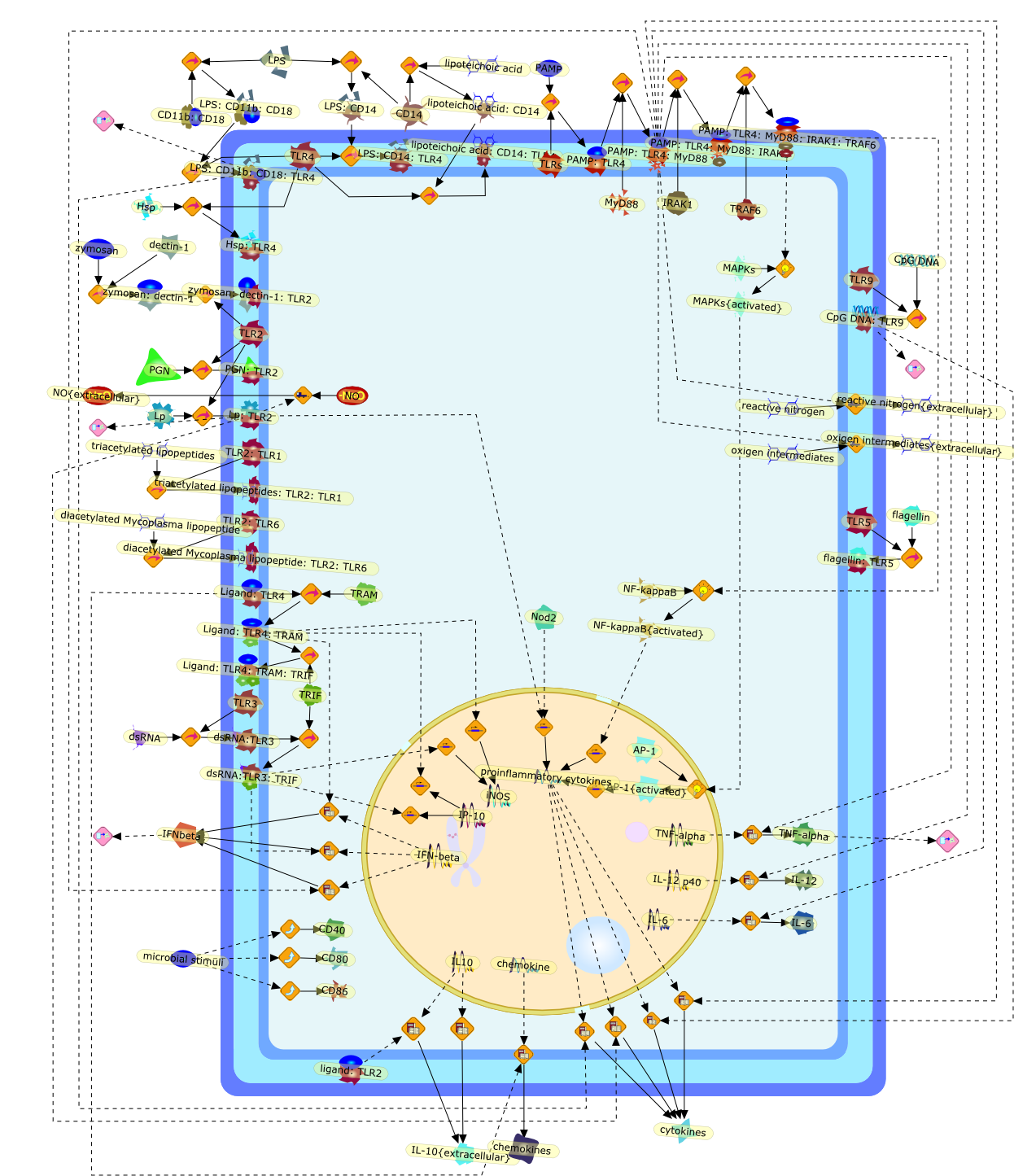{kind=link}
{kind=link}
AP-1,AP-1{activated},CD11b: CD18,CD14,CD40,CD80,CD86,CpG DNA,CpG DNA: TLR9,Hsp,Hsp: TLR4,IFN-beta,IFNbeta,IL-10{extracellular},IL-12,IL-12 p40,IL-6,IL10,IP-10,IRAK1,LPS,LPS: CD11b: CD18,LPS: CD11b: CD18: TLR4,LPS: CD14,LPS: CD14: TLR4,Ligand: TLR4,Ligand: TLR4: TRAM,Ligand: TLR4: TRAM: TRIF,Lp,Lp: TLR2,MAPKs,MAPKs{activated},MyD88,NF-kappaB,NF-kappaB{activated},NO,NO{extracellular},Nod2,PAMP,PAMP: TLR4,PAMP: TLR4: MyD88,PAMP: TLR4: MyD88: IRAK1,PAMP: TLR4: MyD88: IRAK1: TRAF6,PGN,PGN: TLR2,TLR2,TLR2: TLR1,TLR2: TLR6,TLR3,TLR4,TLR5,TLR9,TLRs,TNF-alpha,TRAF6,TRAM,TRIF,chemokine,chemokines,cytokines,dectin-1,diacetylated Mycoplasma lipopeptide,diacetylated Mycoplasma lipopeptide: TLR2: TLR6,dsRNA,dsRNA:TLR3,dsRNA:TLR3: TRIF,flagellin,flagellin: TLR5,iNOS,ligand: TLR2,lipoteichoic acid,lipoteichoic acid: CD14,lipoteichoic acid: CD14: TLR4,microbial stimuli,oxigen intermediates,oxigen intermediates{extracellular},proinflammatory cytokines,reactive nitrogen,reactive nitrogen{extracellular},triacetylated lipopeptides,triacetylated lipopeptides: TLR2: TLR1,zymosan,zymosan: dectin-1,zymosan: dectin-1: TLR2
The discovery of the Toll-like receptors (TLRs) has revolutionised the field ofinnate immunity. One unresolved question regarding LPS signalling is whetherthere is a role for tyrosine kinases downstream of the LPS receptor. Studies inmice deficient in Bruton's tyrosine kinase have previously shown that they aredefective in their responses to LPS. Further investigation into the role of Btkin LPS signalling has directly implicated Btk downstream of TLR4, both withrespect to p38 MAPK activation and activation of the transcription factorNFkappaB. In fact Btk is activated by LPS and has been shown to directly bindTLR4 and the key proximal signalling proteins involved in LPS-induced NFkappaBactivation, MyD88, Mal and IRAK-1. These recent findings point to a direct rolefor Btk in LPS signal transduction and raise interesting questions regarding themode of activation of Btk following LPS stimulation and the precise nature ofthe pathways activated downstream of Btk. A better understanding of how Btkfunctions in LPS signalling will have important implications for inflammatoryand autoimmune disorders and therapies thereof.
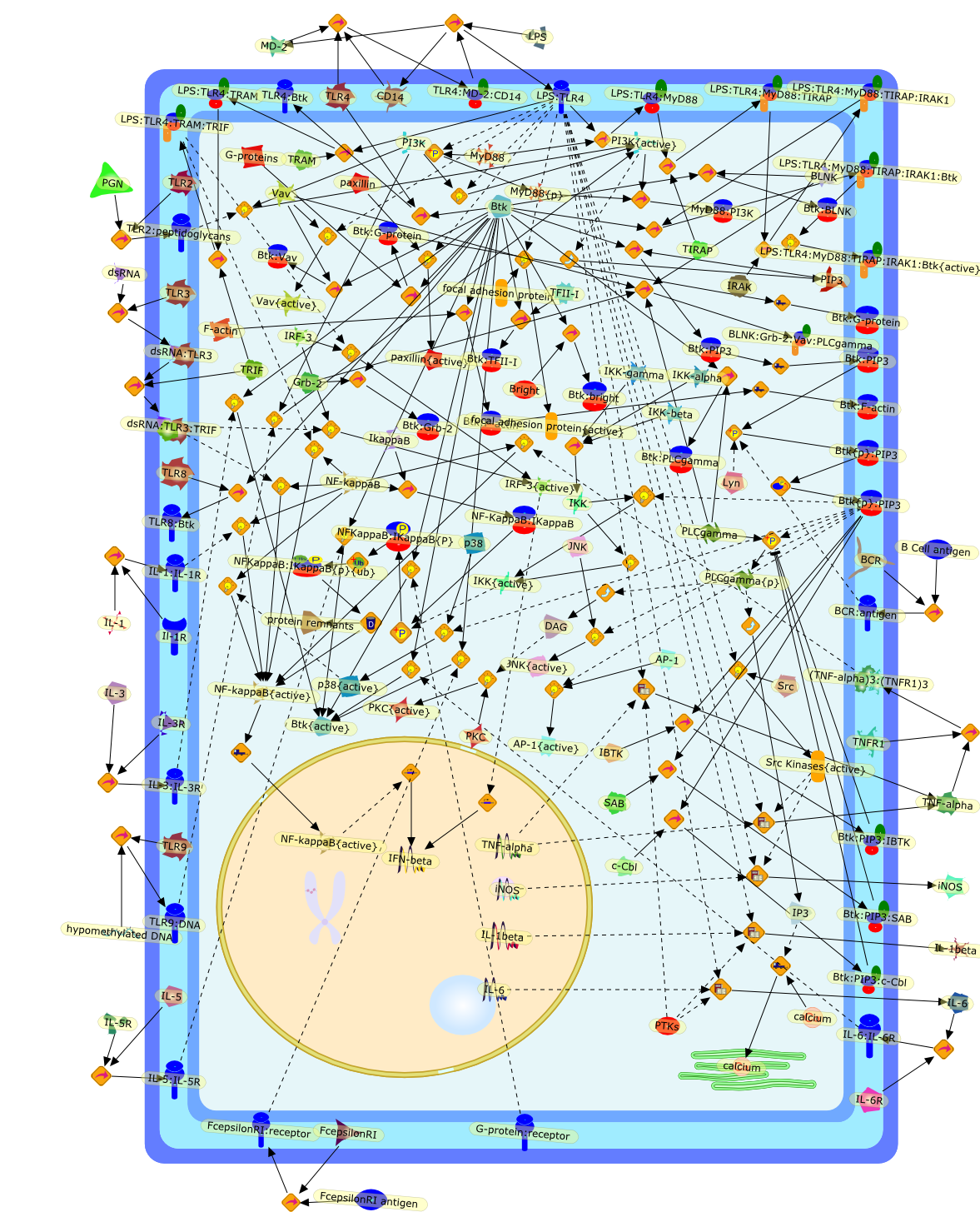{kind=link}
{kind=link}
(TNF-alpha)3:(TNFR1)3,AP-1,AP-1{active},B Cell antigen,BCR,BCR:antigen,BLNK,BLNK:Grb-2:Vav:PLCgamma,Bright,Btk,Btk:BLNK,Btk:F-actin,Btk:G-protein,Btk:Grb-2,Btk:PIP3,Btk:PIP3:IBTK,Btk:PIP3:SAB,Btk:PIP3:c-Cbl,Btk:PLCgamma,Btk:TFII-I,Btk:Vav,Btk:bright,Btk{active},Btk{p}:PIP3,CD14,DAG,F-actin,FcepsilonRI,FcepsilonRI antigen,FcepsilonRI:receptor,G-protein:receptor,G-proteins,Grb-2,IBTK,IFN-beta,IKK,IKK-alpha,IKK-beta,IKK-gamma,IKK{active},IL-1,IL-1:IL-1R,IL-1beta,IL-3,IL-3:IL-3R,IL-3R,IL-5,IL-5:IL-5R,IL-5R,IL-6,IL-6:IL-6R,IL-6R,IP3,IRAK,IRF-3,IRF-3{active},IkappaB,Il-1R,JNK,JNK{active},LPS,LPS:TLR4,LPS:TLR4:MyD88,LPS:TLR4:MyD88:TIRAP,LPS:TLR4:MyD88:TIRAP:IRAK1,LPS:TLR4:MyD88:TIRAP:IRAK1:Btk,LPS:TLR4:MyD88:TIRAP:IRAK1:Btk{active},LPS:TLR4:TRAM,LPS:TLR4:TRAM:TRIF,Lyn,MD-2,MyD88,MyD88:PI3K,MyD88{p},NF-KappaB:IKappaB,NF-kappaB,NF-kappaB{active},NFKappaB:IKappaB{P},NFKappaB:IKappaB{p}{ub},PGN,PI3K,PI3K{active},PIP3,PKC,PKC{active},PLCgamma,PLCgamma{p},PTKs,SAB,Src,Src Kinases{active},TFII-I,TIRAP,TLR2,TLR2:peptidoglycans,TLR3,TLR4,TLR4:Btk,TLR4:MD-2:CD14,TLR8,TLR8:Btk,TLR9,TLR9:DNA,TNF-alpha,TNFR1,TRAM,TRIF,Vav,Vav{active},c-Cbl,calcium,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,focal adhesion protein,focal adhesion protein{active},hypomethylated DNA,iNOS,p38,p38{active},paxillin,paxillin{active},protein remnants
Extensive work has suggested that a number of endogenous molecules such as heatshock proteins (hsp) may be potent activators of the innate immune systemcapable of inducing proinflammatory cytokine production by themonocyte-macrophage system and the activation and maturation of dendritic cells.The cytokine-like effects of these endogenous molecules are mediated via theToll-like receptor (TLR) signal-transduction pathways in a manner similar tolipopolysaccharide (LPS; via TLR4) and bacterial lipoproteins (via TLR2).However, recent evidence suggests that the reported cytokine effects of hsp maybe a result of the contaminating LPS and LPS-associated molecules. The reasonsfor previous failure to recognize the contaminant(s) being responsible for theputative TLR ligands of hsp include failure to use highly purified hsp free ofLPS contamination; failure to recognize the heat sensitivity of LPS; and failureto consider contaminant(s) other than LPS. Whether other reported putativeendogenous ligands of TLR2 and TLR4 are a result of contamination ofpathogen-associated molecular patterns is not clear. It is essential thatefforts should be directed to conclusively determine whether the reportedputative endogenous ligands of TLRs are a result of the endogenous molecules orof contaminant(s), before exploring further the implication and therapeuticpotential of these putative TLR ligands.
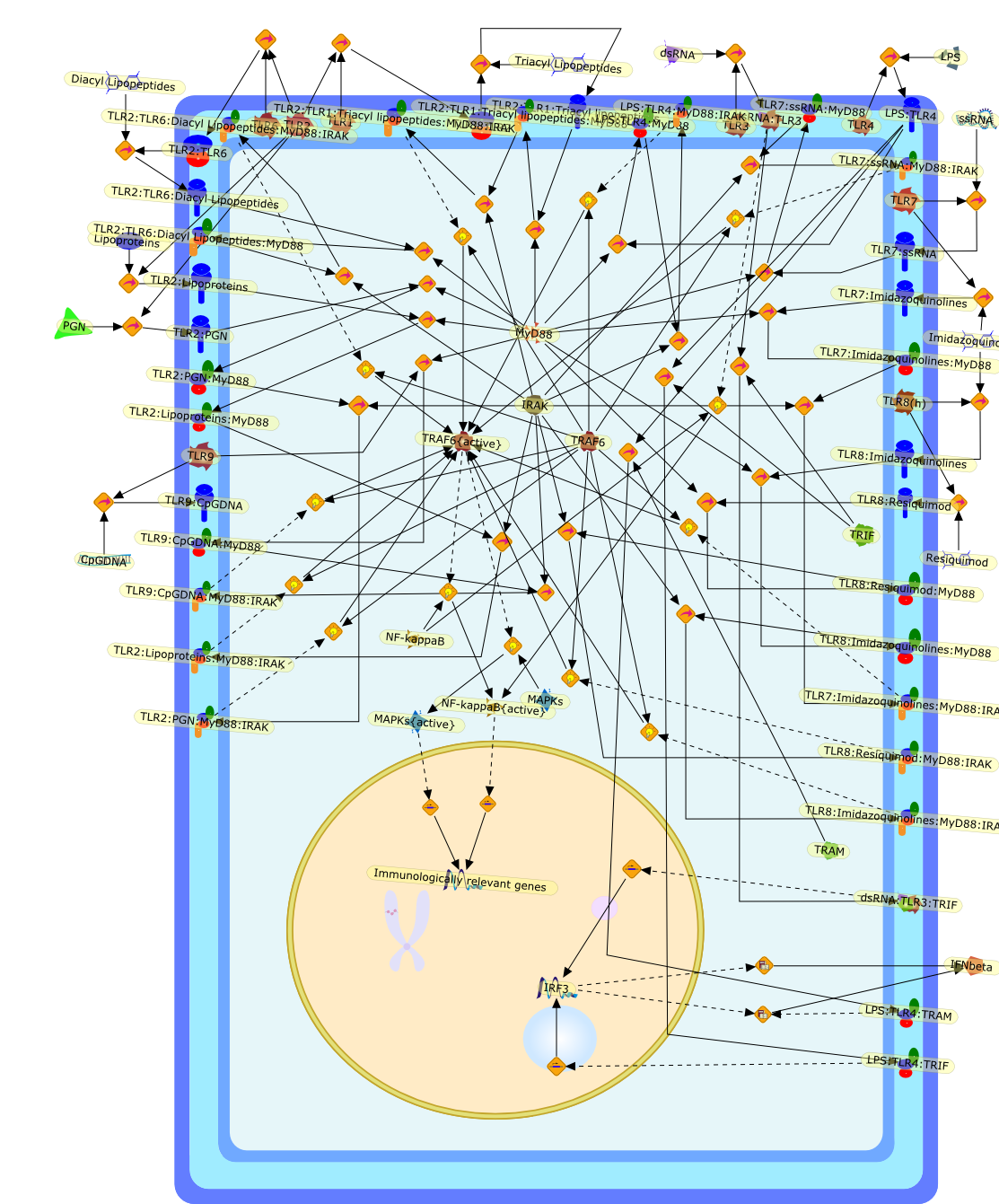{kind=link}
{kind=link}
CpGDNA,Diacyl Lipopeptides,IFNbeta,IRAK,IRF3,Imidazoquinolines,Immunologically relevant genes,LPS,LPS:TLR4,LPS:TLR4:MyD88,LPS:TLR4:MyD88:IRAK,LPS:TLR4:TRAM,LPS:TLR4:TRIF,Lipoproteins,MAPKs,MAPKs{active},MyD88,NF-kappaB,NF-kappaB{active},PGN,Resiquimod,TLR1,TLR1:TLR2,TLR2,TLR2:Lipoproteins,TLR2:Lipoproteins:MyD88,TLR2:Lipoproteins:MyD88:IRAK,TLR2:PGN,TLR2:PGN:MyD88,TLR2:PGN:MyD88:IRAK,TLR2:TLR1:Triacyl lipopeptides,TLR2:TLR1:Triacyl lipopeptides:MyD88,TLR2:TLR1:Triacyl lipopeptides:MyD88:IRAK,TLR2:TLR6,TLR2:TLR6:Diacyl Lipopeptides,TLR2:TLR6:Diacyl Lipopeptides:MyD88,TLR2:TLR6:Diacyl Lipopeptides:MyD88:IRAK,TLR3,TLR4,TLR6,TLR7,TLR7:Imidazoquinolines,TLR7:Imidazoquinolines:MyD88,TLR7:Imidazoquinolines:MyD88:IRAK,TLR7:ssRNA,TLR7:ssRNA:MyD88,TLR7:ssRNA:MyD88:IRAK,TLR8(h),TLR8:Imidazoquinolines,TLR8:Imidazoquinolines:MyD88,TLR8:Imidazoquinolines:MyD88:IRAK,TLR8:Resiquimod,TLR8:Resiquimod:MyD88,TLR8:Resiquimod:MyD88:IRAK,TLR9,TLR9:CpGDNA,TLR9:CpGDNA:MyD88,TLR9:CpGDNA:MyD88:IRAK,TRAF6,TRAF6{active},TRAM,TRIF,Triacyl Lipopeptides,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,ssRNA
The macrophage is an important cell type in the pathophysiology of humanimmunodeficiency virus type 1 (HIV-1) infection. Macrophages both support viralreplication and are capable of attracting and activating lymphocytes, thusrendering CD4+ T lymphocytes highly permissive for infection. The viral Tatprotein, whose function is mediated by the cellular cyclin T1 protein complexedwith CDK9, is required for efficient transcription of the integrated HIV-1provirus by RNA polymerase II. Cyclin T1 expression is highly regulated duringmacrophage differentiation, and this has important implications for HIV-1replication. In monocytes isolated from healthy blood donors, cyclin T1 proteinexpression is low and is induced to high levels within the first few days ofdifferentiation by a post-transcriptional mechanism. After 1-2 weeks ofmacrophage differentiation, however, cyclin T1 expression is shut off. Treatmentof macrophages with lipopolysaccharide (LPS) can re-induce cyclin T1, indicatingthat the activation status of macrophages can regulate cyclin T1 expression.Recent results indicate that HIV-1 infection is able to induce cyclin T1expression in macrophages. Future studies of cyclin T1 regulation in macrophagesmay suggest means of manipulating expression of this crucial cellular co-factorfor therapeutic benefit in HIV-1 infected individuals. Copyright 2004 ElsevierB.V.
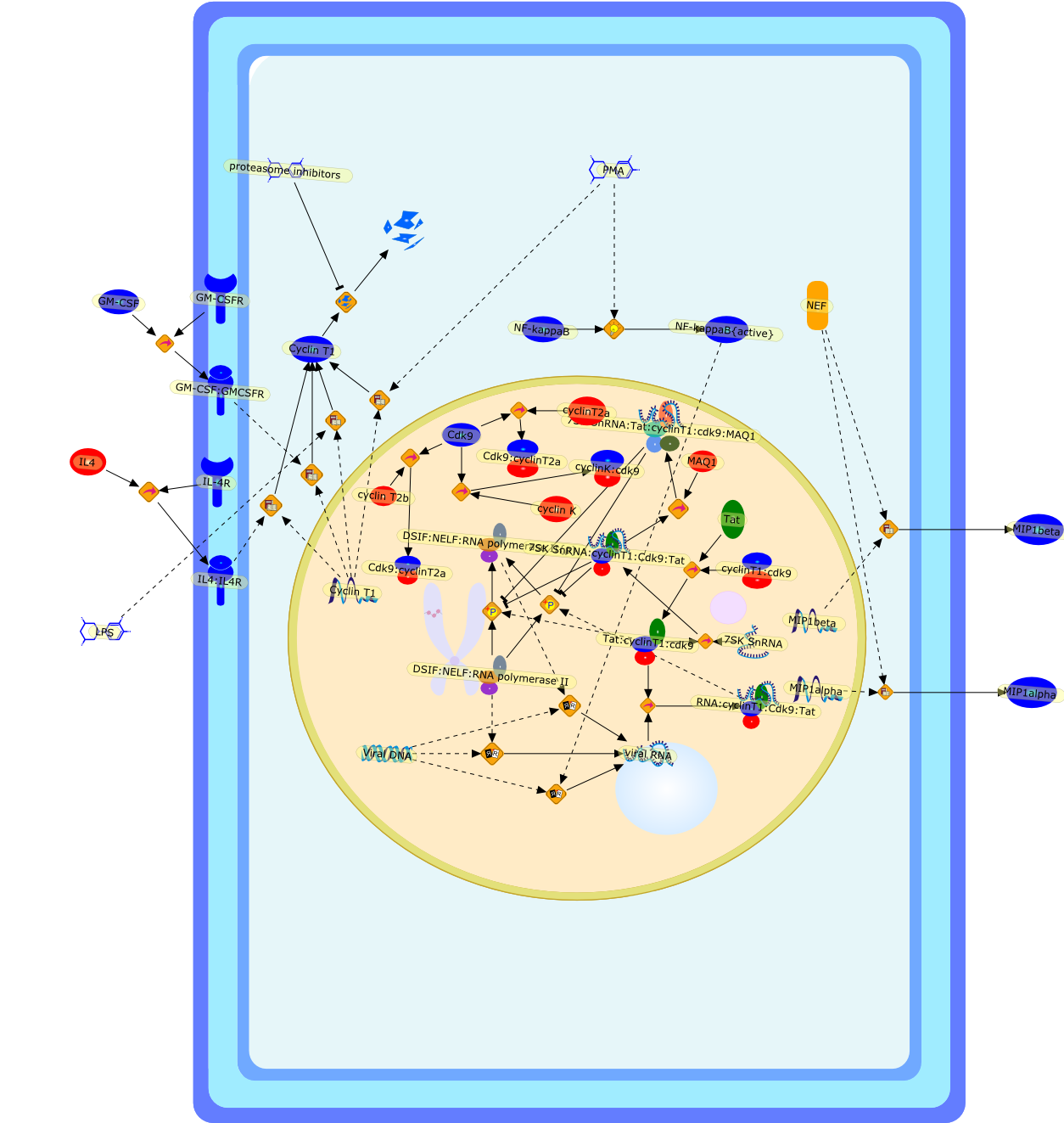{kind=link}
{kind=link}
7SK SnRNA,7SK SnRNA:Tat:cyclinT1:cdk9:MAQ1,7SK SnRNA:cyclinT1:Cdk9:Tat,Cdk9,Cdk9:cyclinT2a,Cyclin T1,DSIF:NELF:RNA polymerase II,DSIF:NELF:RNA polymerase II{p},GM-CSF,GM-CSF:GMCSFR,GM-CSFR,IL-4R,IL4,IL4:IL4R,LPS,MAQ1,MIP1alpha,MIP1beta,NEF,NF-kappaB,NF-kappaB{active},PMA,RNA:cyclinT1:Cdk9:Tat,Tat,Tat:cyclinT1:cdk9,Viral DNA,cyclin K,cyclin T2b,cyclinK:cdk9,cyclinT1:cdk9,cyclinT2a,proteasome inhibitors,protein remnants,viral RNA
Lysophospholipids (LPs), such as lysophosphatidic acid and sphingosine1-phosphate, are membrane-derived bioactive lipid mediators. LPs can affectfundamental cellular functions, which include proliferation, differentiation,survival, migration, adhesion, invasion, and morphogenesis. These functionsinfluence many biological processes that include neurogenesis, angiogenesis,wound healing, immunity, and carcinogenesis. In recent years, identification ofmultiple cognate G protein-coupled receptors has provided a mechanisticframework for understanding how LPs play such diverse roles. Generation of LPreceptor-null animals has allowed rigorous examination of receptor-mediatedphysiological functions in vivo and has identified new functions for LP receptorsignaling. Efforts to develop LP receptor subtype-specific agonists/antagonistsare in progress and raise expectations for a growing collection of chemicaltools and potential therapeutic compounds. The rapidly expanding literature onthe LP receptors is herein reviewed.
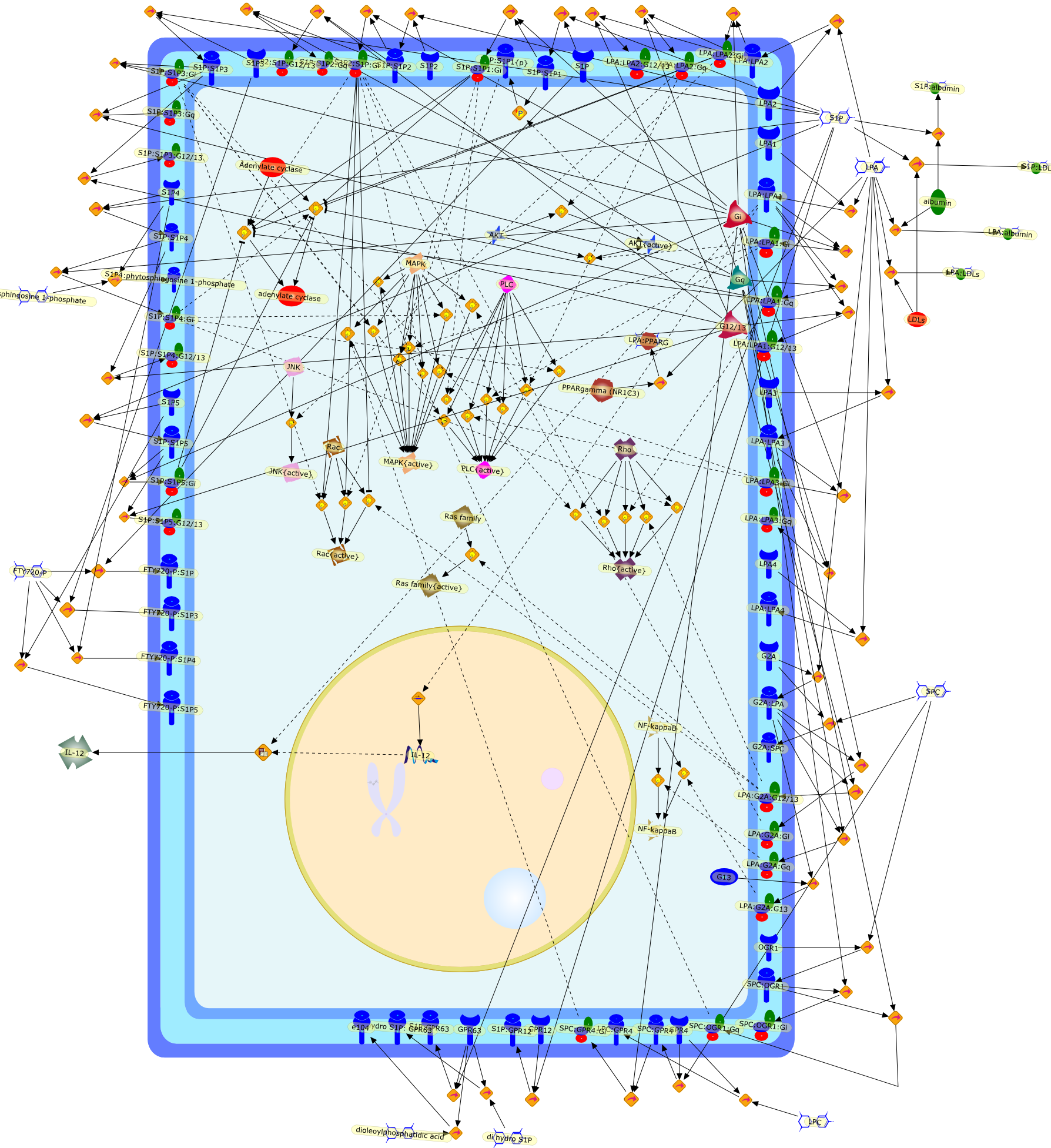{kind=link}
{kind=link}
AKT,AKT{active},Adenylate cyclase,FTY720-P,FTY720-P:S1P,FTY720-P:S1P3,FTY720-P:S1P4,FTY720-P:S1P5,G12/13,G13,G2A,G2A:LPA,G2A:SPC,GPR12,GPR4,GPR63,Gi,Gq,IL-12,JNK,JNK{active},LDLs,LPA,LPA1,LPA2,LPA3,LPA4,LPA:G2A:G12/13,LPA:G2A:G13,LPA:G2A:Gi,LPA:G2A:Gq,LPA:LDLs,LPA:LPA1,LPA:LPA1:G12/13,LPA:LPA1:Gi,LPA:LPA1:Gq,LPA:LPA2,LPA:LPA2:G12/13,LPA:LPA2:Gi,LPA:LPA2:Gq,LPA:LPA3,LPA:LPA3:Gi,LPA:LPA3:Gq,LPA:LPA4,LPA:PPARG,LPA:albumin,LPC,LPC:GPR4,MAPK,MAPK{active},NF-kappaB,OGR1,PLC,PLC{active},PPARgamma (NR1C3),Rac,Rac{active},Ras family,Ras family{active},Rho,Rho{active},S1P,S1P2,S1P2:S1P:G12/13,S1P2:S1P:Gi,S1P3,S1P4,S1P4:phytosphingosine 1-phosphate,S1P5,S1P:GPR12,S1P:GPR63,S1P:LDLs,S1P:S1P1,S1P:S1P1:Gi,S1P:S1P1{p},S1P:S1P2,S1P:S1P2:Gq,S1P:S1P3,S1P:S1P3:G12/13,S1P:S1P3:Gi,S1P:S1P3:Gq,S1P:S1P4,S1P:S1P4:G12/13,S1P:S1P4:Gi,S1P:S1P5,S1P:S1P5:G12/13,S1P:S1P5:Gi,S1P:albumin,SPC,SPC:GPR4,SPC:GPR4:Gi,SPC:OGR1,SPC:OGR1:Gi,SPC:OGR1:Gq,adenylate cyclase,albumin,di hydro S1P,dihydro S1P: GPR63,dioleoylphosphatidic acid,e104,phytosphingosine 1-phosphate
Non-self-recognition of invading microbes relies on the pattern-recognition ofpathogen-associated molecular patterns (PAMPs) derived from microbial cell-wallcomponents. Insects and mammals conserve a signaling pathway of the innateimmune system through cell-surface receptors called Tolls and Toll-likereceptors (TLRs). Bacterial lipopolysaccharides (LPSs) are an important triggerof the horseshoe crab's innate immunity to infectious microorganisms. Horseshoecrabs' granular hemocytes respond specifically to LPS stimulation, inducing thesecretion of various defense molecules from the granular hemocytes. Here, weshow a cDNA which we named tToll, coding for a TLR identified from hemocytes ofthe horseshoe crab Tachypleus tridentatus. tToll is most closely related toDrosophila Toll in both domain architecture and overall length. Human TLRs havebeen suggested to contain numerous PAMP-binding insertions located in theleucine-rich repeats (LRRs) of their ectodomains. However, the LRRs of tTollcontained no obvious PAMP-binding insertions. Furthermore, tToll wasnon-specifically expressed in horseshoe crab tissues. These observations suggestthat tToll does not function as an LPS receptor on granular hemocytes.
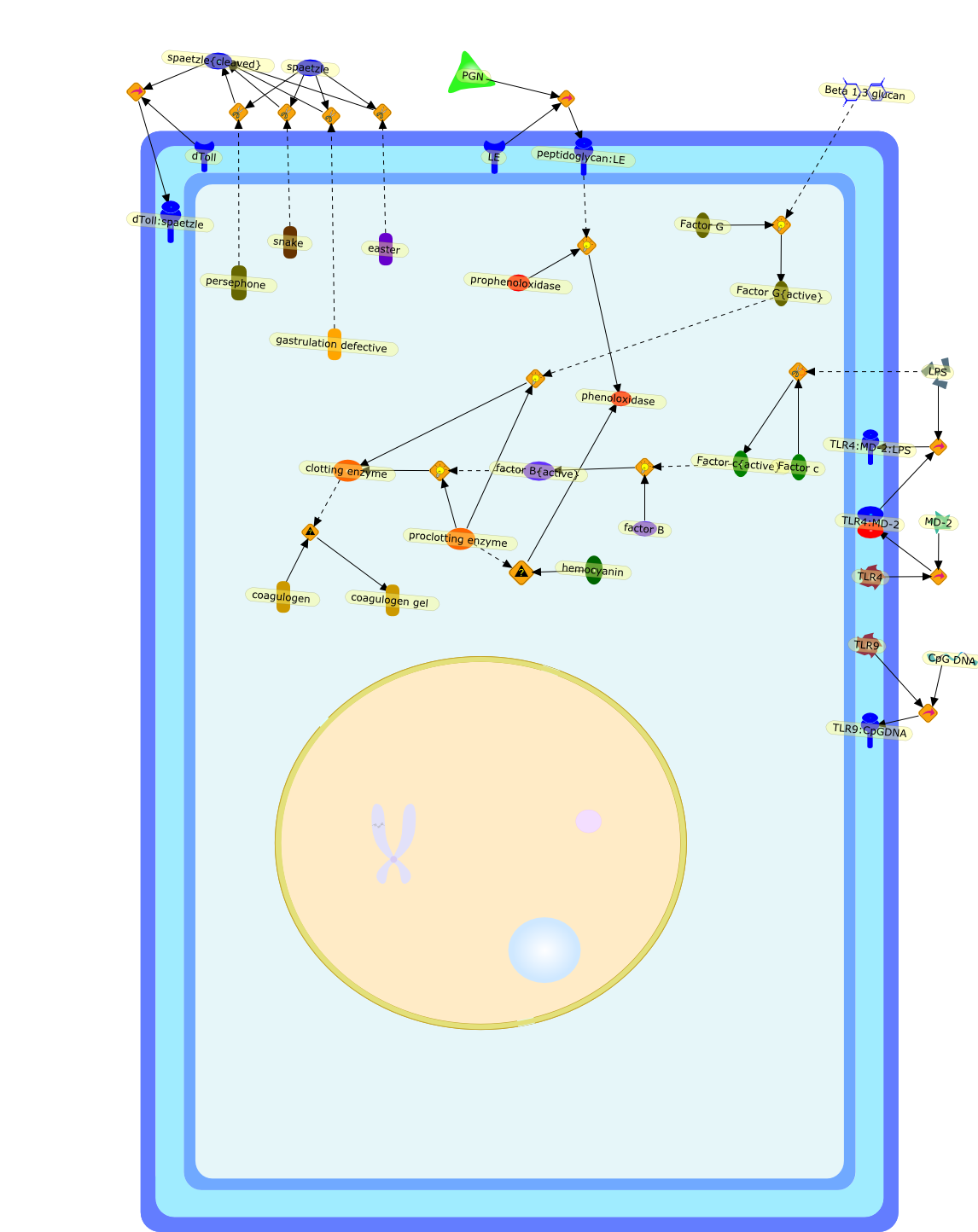{kind=link}
{kind=link}
Beta 1,3 glucan,CpG DNA,Factor G,Factor G{active},Factor c,Factor c{active},LE,LPS,MD-2,PGN,TLR4,TLR4:MD-2,TLR4:MD-2:LPS,TLR9,TLR9:CpGDNA,clotting enzyme,coagulogen,coagulogen gel,dToll,dToll:spaetzle,easter,factor B,factor B{active},gastrulation defective,hemocyanin,peptidoglycan:LE,persephone,phenoloxidase,proclotting enzyme,prophenoloxidase,snake,spaetzle,spaetzle{cleaved}
Lipopolysaccharide (LPS), a major component of the outer membrane ofGram-negative bacteria, can be beneficial to the host by activating the innateimmune system, or harmful, by inducing inflammation, disseminated intravascularcoagulation, multiple organ failure, shock and often death. On the bacteria, andin host biological fluids and cells, LPS is never free but constantly attachedto cognate-binding proteins. Understanding how LPS is transported and furtherrecognized by sensors able to deliver a signal, or by inactivating moleculesable to neutralize its biological effects, is an important goal. This reviewdescribes the large panel of peptides and proteins reported to associate withLPS, and provides information on their origin, their structure and the locationof amino acid residues involved in their interaction with LPS. A betterunderstanding of the mode of recognition of LPS by cognate proteins promptedmany laboratories to design on a rational basis synthetic molecules which can beused to detect low amounts of endotoxin, or to act as efficient blockers of invitro and in vivo responses to LPS.
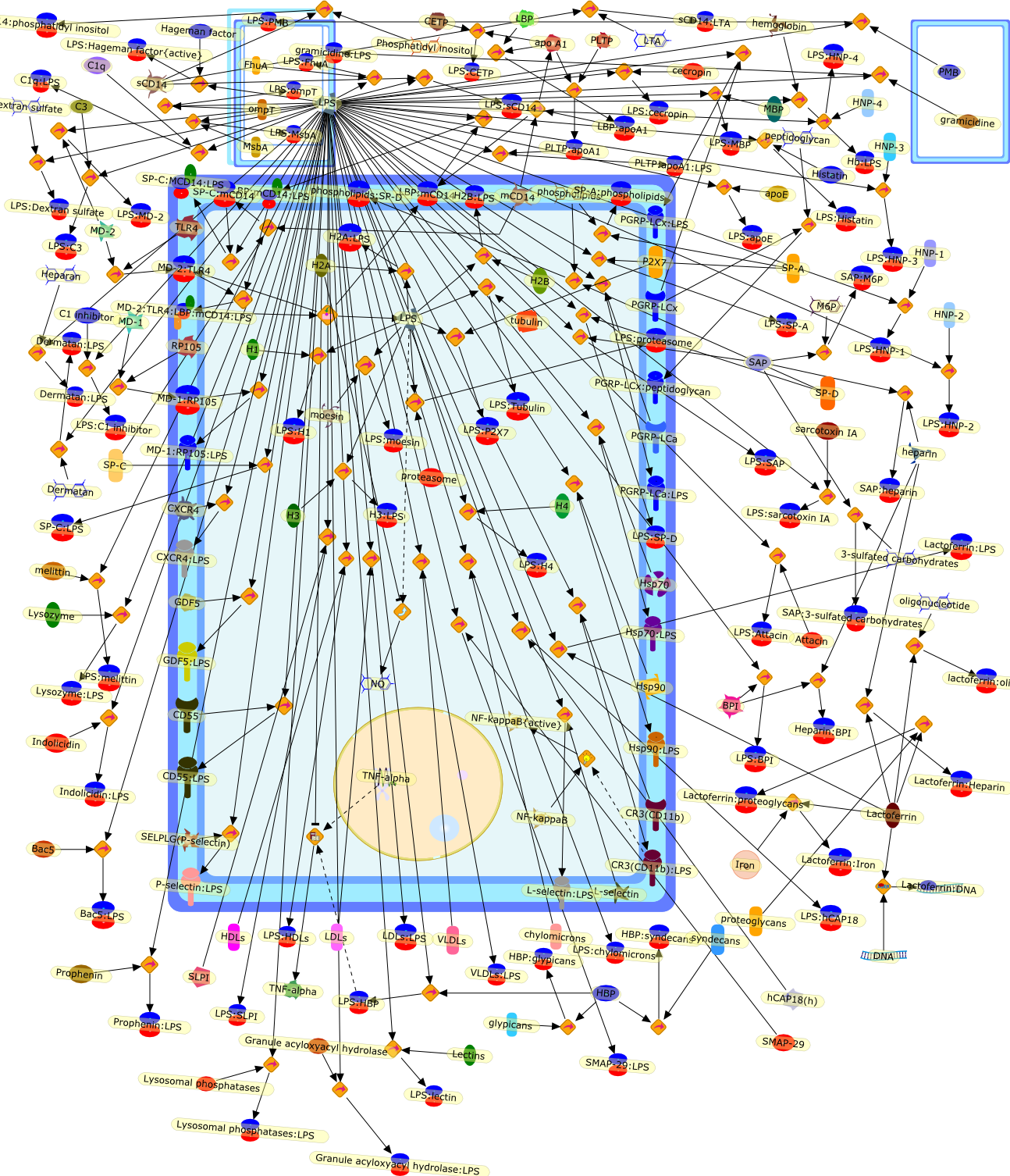{kind=link}
{kind=link}
3-sulfated carbohydrates,Attacin,BPI,Bac5,Bac5:LPS,C1 inhibitor,C1q,C1q:LPS,C3,CD55,CD55:LPS,CETP,CR3(CD11b),CR3(CD11b):LPS,CXCR4,CXCR4:LPS,DNA,Dermatan,Dermatan:LPS,Dextran sulfate,FhuA,GDF5,GDF5:LPS,Granule acyloxyacyl hydrolase,Granule acyloxyacyl hydrolase:LPS,H1,H2A,H2A:LPS,H2B,H2B:LPS,H3,H3:LPS,H4,HBP,HBP:glypicans,HBP:syndecans,HDLs,HNP-1,HNP-2,HNP-3,HNP-4,Hageman factor,Hb:LPS,Heparan,Heparin:BPI,Histatin,Hsp70,Hsp70:LPS,Hsp90,Hsp90:LPS,Indolicidin,Indolicidin:LPS,Iron,L-selectin,L-selectin:LPS,LBP,LBP:apoA1,LBP:mCD14,LBP:mCD14:LPS,LDLs,LDLs:LPS,LPS,LPS:Attacin,LPS:BPI,LPS:C1 inhibitor,LPS:C3,LPS:CETP,LPS:Dextran sulfate,LPS:FhuA,LPS:H1,LPS:H4,LPS:HBP,LPS:HDLs,LPS:HNP-1,LPS:HNP-2,LPS:HNP-3,LPS:HNP-4,LPS:Hageman factor{active},LPS:Histatin,LPS:MBP,LPS:MD-2,LPS:MsbA,LPS:P2X7,LPS:PMB,LPS:SAP,LPS:SLPI,LPS:SP-A,LPS:SP-D,LPS:Tubulin,LPS:apoE,LPS:cecropin,LPS:chylomicrons,LPS:hCAP18,LPS:lectin,LPS:melittin,LPS:moesin,LPS:ompT,LPS:proteasome,LPS:sCD14,LPS:sarcotoxin IA,LTA,Lactoferrin,Lactoferrin:DNA,Lactoferrin:Heparin,Lactoferrin:Iron,Lactoferrin:LPS,Lactoferrin:proteoglycans,Lectins,Lysosomal phosphatases,Lysosomal phosphatases:LPS,Lysozyme,Lysozyme:LPS,M6P,MBP,MD-1,MD-1:RP105,MD-1:RP105:LPS,MD-2,MD-2:TLR4,MD-2:TLR4:LBP:mCD14:LPS,MsbA,NF-kappaB,NF-kappaB{active},NO,P-selectin:LPS,P2X7,PGRP-LCa,PGRP-LCa:LPS,PGRP-LCx,PGRP-LCx:LPS,PGRP-LCx:peptidoglycan,PLTP,PLTP:apoA1,PLTP:apoA1:LPS,PMB,Phosphatidyl inositol,Prophenin,Prophenin:LPS,RP105,SAP,SAP:3-sulfated carbohydrates,SAP:M6P,SAP:heparin,SELPLG(P-selectin),SLPI,SMAP-29,SMAP-29:LPS,SP-A,SP-A:phospholipids,SP-C,SP-C:LPS,SP-C:MCD14:LPS,SP-C:mCD14,SP-D,TLR4,TNF-alpha,VLDLs,VLDLs:LPS,apo A1,apoE,cecropin,chylomicrons,glypicans,gramicidine,gramicidine:LPS,hCAP18(h),hemoglobin,heparin,lactoferrin:oligo,mCD14,melittin,moesin,oligonucleotide,ompT,peptidoglycan,phospholipids,phospholipids:SP-D,proteasome,proteoglycans,sCD14,sCD14:LTA,sCD14:phosphatidyl inositol,sarcotoxin IA,syndecans,tubulin
Despite advances in treatment strategies for hepatitis C virus (HCV), asignificant proportion of patients fail to achieve viral clearance followingtreatment with pegylated interferon (IFN)-alpha plus ribavirin. Many of theseindividuals show elevated levels of tumor necrosis factor (TNF)-alpha comparedwith normal controls, and recent data have implicated this cytokine in thenegative regulation of IFN-alpha. Although a therapeutic opportunity forTNF-alpha antagonists might exist for reducing inflammation in chronic HCVdisease, further exploration is required to identify the key mediators ofresponsiveness to IFN-alpha. In particular, the interplay should be clarifiedbetween host response factors [e.g. IFN-alpha, IFN-gamma, suppressor of cytokinesignaling (SOCS), TNF-alpha and others] and pathogen-associated molecularpatterns [PAMPs, e.g. lipopolysaccharide (LPS) and CpG DNA] in HCV disease; thisinformation might guide future therapies aimed at improving IFN-alpharesponsiveness.
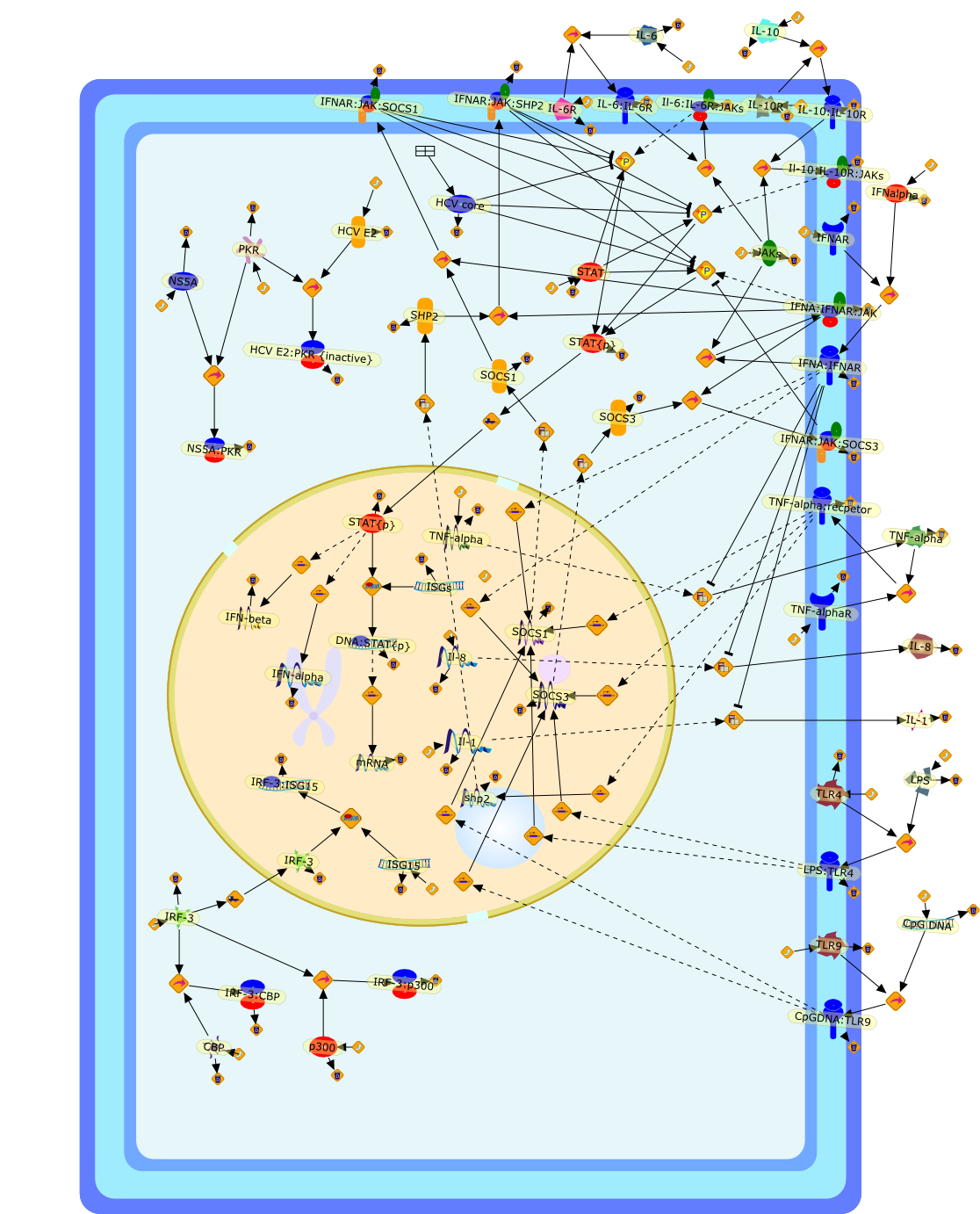{kind=link}
{kind=link}
CBP,CpG DNA,CpGDNA:TLR9,DNA:STAT{p},HCV E2,HCV E2:PKR {inactive},HCV core,IFN-alpha,IFN-beta,IFNA:IFNAR,IFNA:IFNAR:JAK,IFNAR,IFNAR:JAK:SHP2,IFNAR:JAK:SOCS1,IFNAR:JAK:SOCS3,IFNalpha,IL-1,IL-10,IL-10:IL-10R,IL-10R,IL-6,IL-6:IL-6R,IL-6R,IL-8,IRF-3,IRF-3:CBP,IRF-3:ISG15,IRF-3:p300,ISG15,ISGs,Il-1,Il-10:IL-10R:JAKs,Il-6:IL-6R:JAKs,Il-8,JAKs,LPS,LPS:TLR4,NS5A,NS5A:PKR,PKR,SHP2,SOCS1,SOCS3,STAT,STAT{p},TLR4,TLR9,TNF-alpha,TNF-alpha:recpetor,TNF-alphaR,mRNA,p300,shp2
Many physiological functions of the body change during the aging process.Dysregulated immune and inflammatory responses have been well documented in bothhumans and animals. The investigation into the cellular and molecular mechanismunderlying these disorders has provided compelling evidence that up-regulatedcyclooxygenase (COX)-2 and its product, particularly prostaglandin (PG)E2, playa critical role in the age-associated dysregulation of the immune andinflammatory responses. In particular, several studies have shown that increasedPGE2 production in old macrophages (Mphi) contributes to the suppression of Tcell function with aging. Furthermore, interventions targeted at decreasing PGE2production have been shown to enhance T cell-mediated function. COX-2 and itscatalytic products are also suggested to play a key role in age-relatedneurodegenerative diseases such as Alzheimer's and Parkinson's disease.Administration of anti-inflammatory drugs which inhibit COX activity has beenshown, by some investigators, to be beneficial in preventing and treating thesediseases. It is, thus, important to understand the underlying mechanisms ofage-related COX-2 up-regulation and to delineate the factors, which contributeto this age-related change. This review focuses on the regulation of PGE2production in murine Mphi; the age-associated changes in COX-2 expression; andits implication for certain disorders observed in the aged immune system andbrain. Increased PGE2 production has been shown to be mainly due to an increasein COX activity, which is, in turn, due to an increase in COX-2 protein and mRNAexpression. Elevated COX-2 mRNA represents a higher transcription rate ratherthan an altered stability of COX-2 mRNA. Upon stimulation, Mphi from old micegenerate more ceramide, a sphingolipid, than those from young mice. Ceramide hasbeen shown to induce, by itself, and also augment, LPS-stimulated COX-2expression and PGE2 production. Several lines of evidence indicate that thehigher ceramide levels in old Mphi are an important contributor to theage-associated up-regulation of COX-2 in Mphi. Ceramide up-regulates COX-2transcription by increasing activation of transcription factor NF-kappaB.Further understanding of molecular mechanisms involved in COX-2 up-regulationwill help in delineating fundamental age-related changes, which lead to thedevelopment of immune and neurological disorders in the aged.
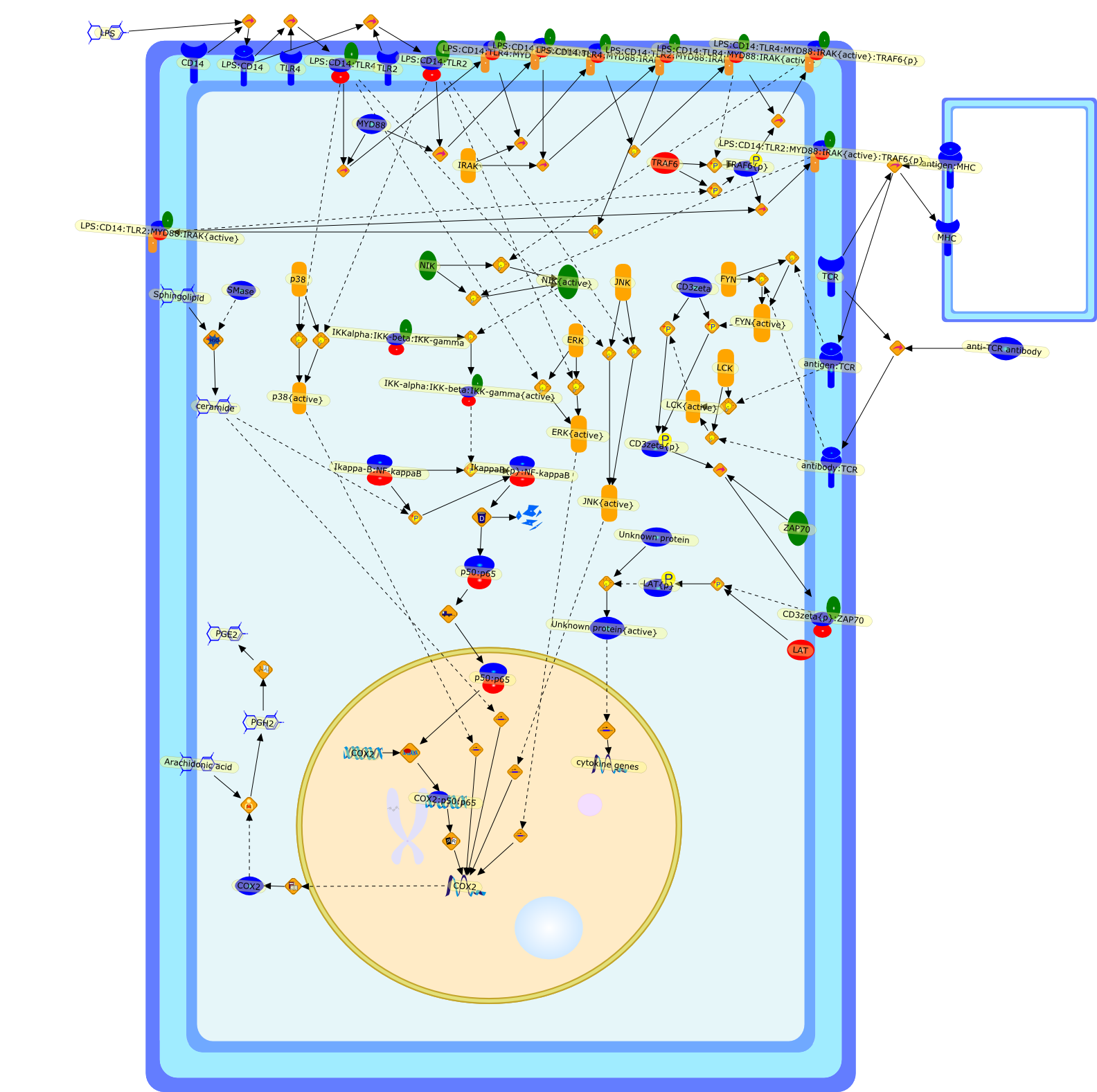{kind=link}
{kind=link}
Arachidonic acid,CD14,CD3zeta,CD3zeta{p},CD3zeta{p}:ZAP70,COX2,COX2:p50:p65,ERK,ERK{active},FYN,FYN{active},IKK-alpha:IKK-beta:IKK-gamma{active},IKKalpha:IKK-beta:IKK-gamma,IRAK,Ikappa-B:NF-kappaB,IkappaB{p}:NF-kappaB,JNK,JNK{active},LAT,LAT{p},LCK,LCK{active},LPS,LPS:CD14,LPS:CD14:TLR2,LPS:CD14:TLR2:MYD88,LPS:CD14:TLR2:MYD88:IRAK,LPS:CD14:TLR2:MYD88:IRAK{active},LPS:CD14:TLR2:MYD88:IRAK{active}:TRAF6{p},LPS:CD14:TLR4,LPS:CD14:TLR4:MYD88,LPS:CD14:TLR4:MYD88:IRAK,LPS:CD14:TLR4:MYD88:IRAK{active},LPS:CD14:TLR4:MYD88:IRAK{active}:TRAF6{p},MHC,MYD88,NIK,NIK{active},PGE2,PGH2,SMase,Sphingolipid,TCR,TLR2,TLR4,TRAF6,TRAF6{p},Unknown protein,Unknown protein{active},ZAP70,anti-TCR antibody,antibody:TCR,antigen:MHC,antigen:TCR,ceramide,cytokine genes,p38,p38{active},p50:p65,protein remnants
Atherosclerosis is a slowly evolutive age-linked disease of large arteries,characterized by a local lipid deposition associated with a chronic inflammatoryresponse, leading potentially to acute plaque rupture, thrombosis and ischemicheart disease. Atherogenesis is a complex sequence of events associating firstexpression of adhesion molecules, recruitment of mononuclear cells to theendothelium, local activation of leukocytes and inflammation, lipid accumulationand foam cell formation. Low density lipoproteins (LDLs) become atherogenicafter undergoing oxidation by vascular cells, that transform them into highlybioreactive oxidized LDL (oxidized LDLs). Oxidized LDLs are involved in foamcell formation, and trigger proatherogenic events such as overexpression ofadhesion molecules, chemoattractant agents growth factors and cytokines involvedin the inflammatory process, cell proliferation and apoptosis. Moreover, thistoxic effect of oxidized LDLs plays probably a role in plaque erosion/ruptureand subsequent atherothrombosis. Several biological effects of oxidized LDLs aremediated through changes in the activity of transcription factors andsubsequently in gene expression. Oxidized LDLs exert a biphasic effect on theredox-sensitive transcription factor NF-kappaB, which can be activated therebyup-regulating proinflammatory gene expression, such as adhesion molecules,tissue factor, scavenger receptor LOX-1. On the other hand, higherconcentrations of oxidized LDLs may inhibit NF-kappaB activation triggered byinflammatory agents such as LPS, and may thereby exert an immunosuppressiveeffect. This review is an attempt to clarify the mechanism by which oxidizedLDLs may up- or down-regulate NF-kappaB, the role of NF-kappaB activation (orinhibition), and the consequences of the oxidized LDLs-mediated NF-kappaBdysregulation and their potential involvement in atherosclerosis.
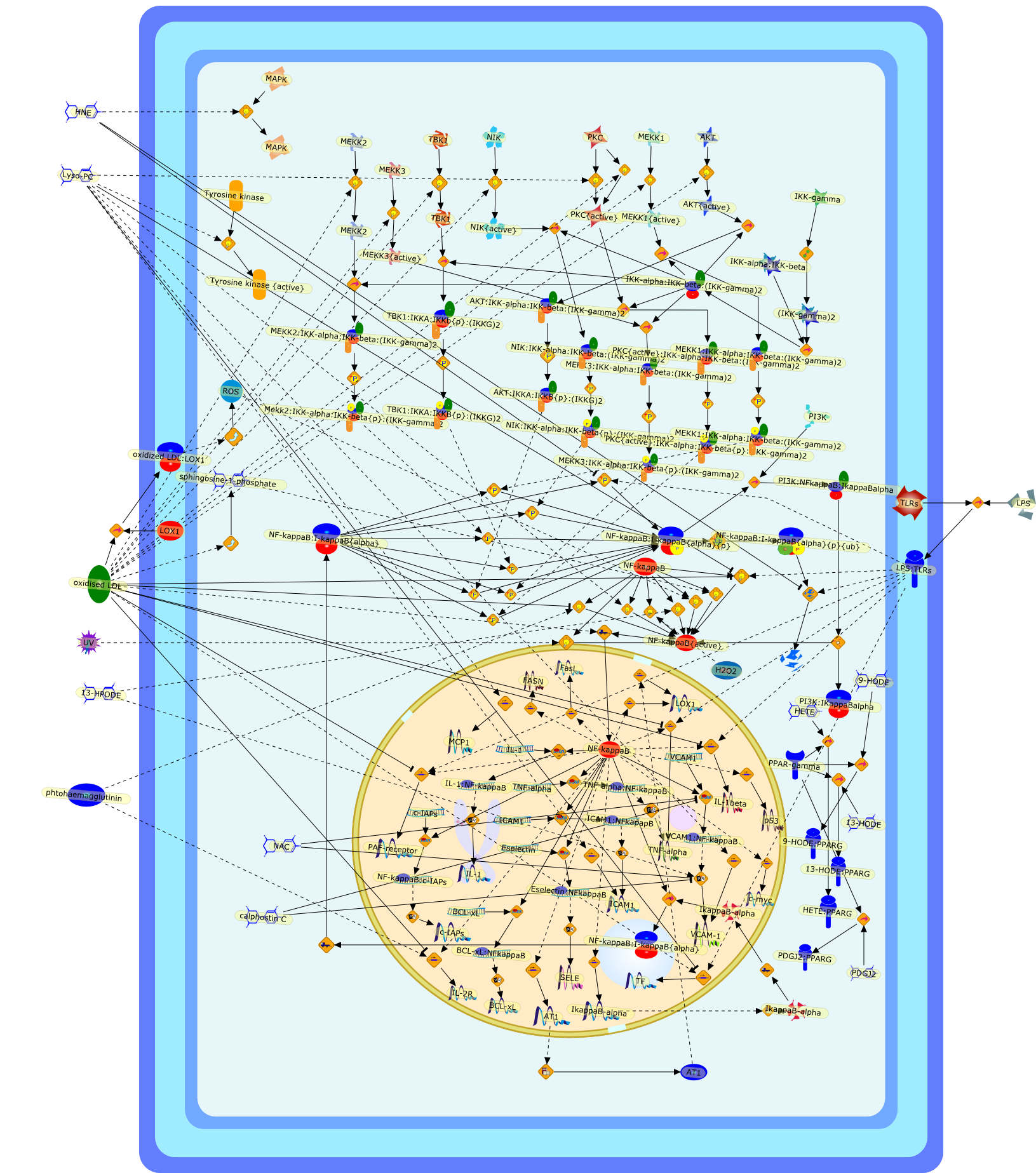{kind=link}
{kind=link}
(IKK-gamma)2,13-HODE,13-HODE:PPARG,13-HPODE,9-HODE,9-HODE:PPARG,AKT,AKT:IKK-alpha:IKK-beta:(IKK-gamma)2,AKT:IKKA:IKKB{p}:(IKKG)2,AKT{active},AT1,BCL-xL,BCL-xL:NFkappaB,Eselectin,Eselectin:NFkappaB,FASN,FasL,H2O2,HETE,HETE:PPARG,HNE,ICAM1,ICAM1:NFkapapB,IKK-alpha:IKK-beta,IKK-alpha:IKK-beta:(IKK-gamma)2,IKK-gamma,IL-1,IL-1:NF-kappaB,IL-1beta,IL-2R,IkappaB-alpha,LOX1,LPS,LPS:TLRs,Lyso-PC,MAPK,MCP1,MEKK1,MEKK1:IKK-alpha:IKK-beta:(IKK-gamma)2,MEKK1{active},MEKK2,MEKK2:IKK-alpha:IKK-beta:(IKK-gamma)2,MEKK3,MEKK3:IKK-alpha:IKK-beta:(IKK-gamma)2,MEKK3:IKK-alpha:IKK-beta{p}:(IKK-gamma)2,MEKK3{active},Mekk2:IKK-alpha:IKK-beta{p}:(IKK-gamma)2,NAC,NF-kappaB,NF-kappaB:I-kappaB{alpha},NF-kappaB:I-kappaB{alpha}{p},NF-kappaB:I-kappaB{alpha}{p}{ub},NF-kappaB:c-IAPs,NF-kappaB{active},NIK,NIK:IKK-alpha:IKK-beta:(IKK-gamma)2,NIK:IKK-alpha:IKK-beta{p}:(IKK-gamma)2,NIK{active},PAF-receptor,PDGJ2,PDGJ2:PPARG,PI3K,PI3K:IKappaBalpha,PI3K:NFkappaB:IkappaBalpha,PKC,PKC{active},PKC{active}:IKK-alpha:IKK-beta:(IKK-gamma)2,PKC{active}:IKK-alpha:IKK-beta{p}:(IKK-gamma)2,PPAR-gamma,ROS,SELE,TBK1,TBK1:IKKA:IKKB{p}:(IKKG)2,TBK1:IKKA:IKKb{p}:(IKKG)2,TF,TLRs,TNF-alpha,TNF-alpha:NF-kappaB,Tyrosine kinase,Tyrosine kinase {active},UV,VCAM-1,VCAM1,VCAM1:NF-kappaB,c-IAPs,c-myc,calphostin C,oxidised LDL,oxidized LDL:LOX1,p53,phtohaemagglutinin,protein remnants,sphingosine-1-phosphate
The intracellular protozoan Toxoplasma gondii exerts profound effects on nuclearfactor-kappaB (NF-kappaB)- and mitogen-activated protein kinase (MAPK)-signalingcascades in macrophages. During early infection, nuclear translocation ofNF-kappaB is blocked, and later, the cells display defects in lipopolysaccharide(LPS)-induced MAPK phosphorylation after undergoing initial activation inresponse to Toxoplasma itself. Infected macrophages that are subjected totriggering through Toll-like receptor 4 (TLR4) with LPS display defectiveproduction of tumor necrosis factor-alpha and IL-12 (IL-12) that likely reflectsinterference with NF-kappaB- and MAPK-signaling cascades. Nevertheless, T.gondii possesses molecules that themselves induce eventual proinflammatorycytokine synthesis. For interleukin-12, this occurs through both myeloiddifferentiation factor 88-dependent and chemokine receptor CCR5-dependentpathways. The balance between activation and interference with proinflammatorysignaling is likely to reflect the need to achieve an appropriate level ofimmunity that allows the host and parasite to maintain a stable interaction.
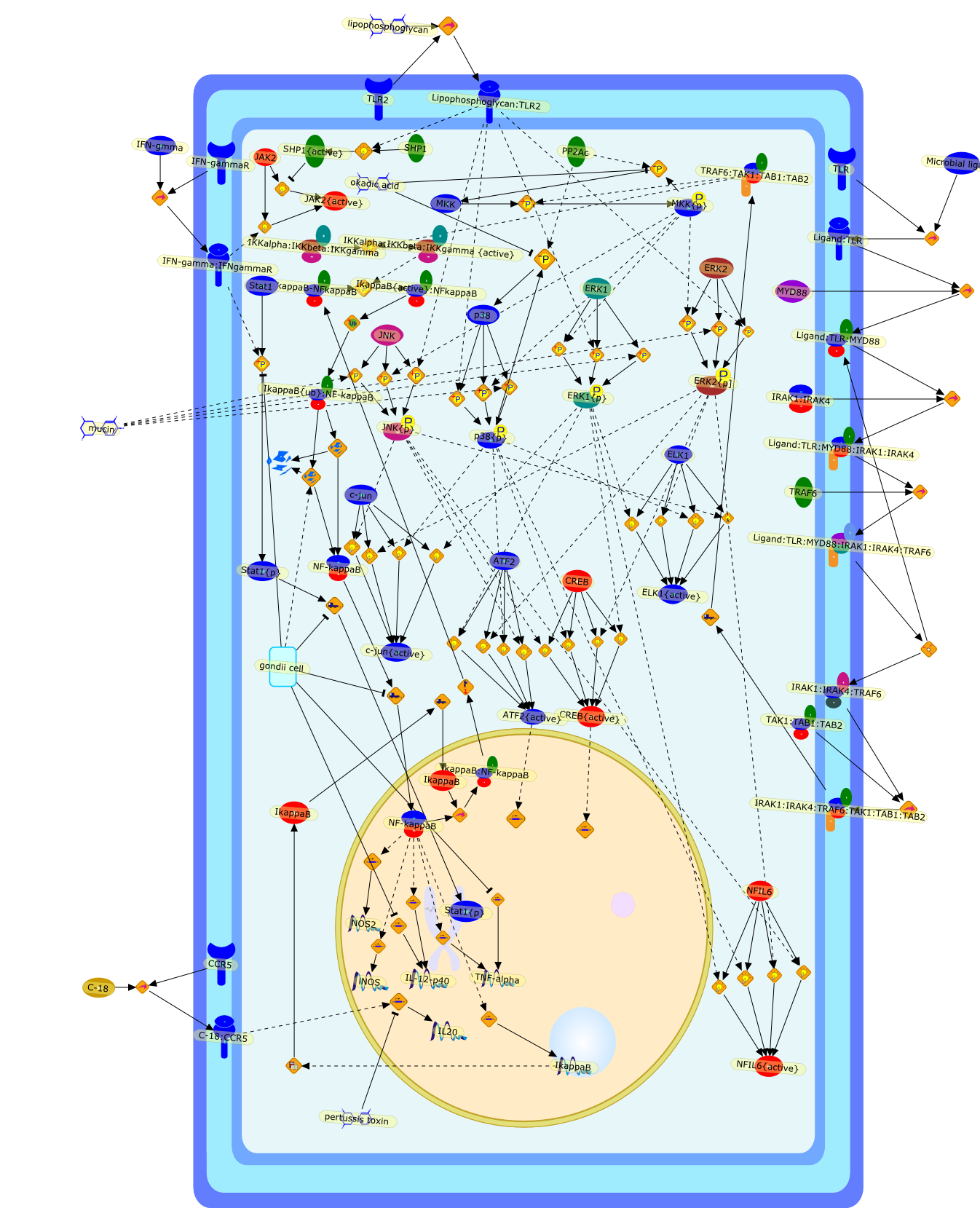{kind=link}
{kind=link}
ATF2,ATF2{active},C-18,C-18:CCR5,CCR5,CREB,CREB{active},ELK1,ELK1{active},ERK1,ERK1{p},ERK2,ERK2{p],IFN-gamma:IFNgammaR,IFN-gammaR,IFN-gmma,IKKalpha:IKKbeta:IKKgamma,IKKalpha:IKKbeta:IKKgamma {active},IL-12-p40,IL20,IRAK1:IRAK4,IRAK1:IRAK4:TRAF6,IRAK1:IRAK4:TRAF6:TAK1:TAB1:TAB2,IkappaB,IkappaB-NFkappaB,IkappaB:NF-kappaB,IkappaB{active}:NFkappaB,IkappaB{ub}:NF-kappaB,JAK2,JAK2{active},JNK,JNK{p},Ligand:TLR,Ligand:TLR:MYD88,Ligand:TLR:MYD88:IRAK1:IRAK4,Ligand:TLR:MYD88:IRAK1:IRAK4:TRAF6,Lipophosphoglycan:TLR2,MKK,MKK{p},MYD88,Microbial ligand,NF-kappaB,NFIL6,NFIL6{active},NOS2,PP2Ac,SHP1,SHP1{active},Stat1,Stat1{p},TAK1:TAB1:TAB2,TLR,TLR2,TNF-alpha,TRAF6,TRAF6:TAK1:TAB1:TAB2,c-jun,c-jun{active},gondii cell,iNOS,lipophosphoglycan,mucin,okadic acid,p38,p38{p},pertussis toxin,protein remnants
An understanding of lipopolysaccharide (LPS) signal transduction is a key goalin the effort to provide a molecular basis for the lethal effect of LPS duringseptic shock and point the way to novel therapies. Rapid progress in this fieldduring the last 6 years has resulted in the discovery of not only the receptorfor LPS - Toll-like receptor 4 (TLR4) - but also in a better appreciation of thecomplexity of the signalling pathways activated by LPS. Soon after the discoveryof TLR4, the formation of a receptor complex in response to LPS, consisting ofdimerized TLR4 and MD-2, was described. Intracellular events following theformation of this receptor complex depend on different sets of adapters. Anearly response, which is dependent on MyD88 and MyD88-like adapter (Mal), leadsto the activation of nuclear factor-kappaB (NF-kappaB). A later response to LPSmakes use of TIR-domain-containing adapter-inducing interferon-beta (TRIF) andTRIF-related adapter molecule (TRAM), and leads to the late activation ofNF-kappaB and IRF3, and to the induction of cytokines, chemokines, and othertranscription factors. As LPS signal transduction is an area of intense researchand rapid progress, this review is intended to sum up our present understandingof the events following LPS binding to TLR4, and we also attempt to create amodel of the signalling pathways activated by LPS.
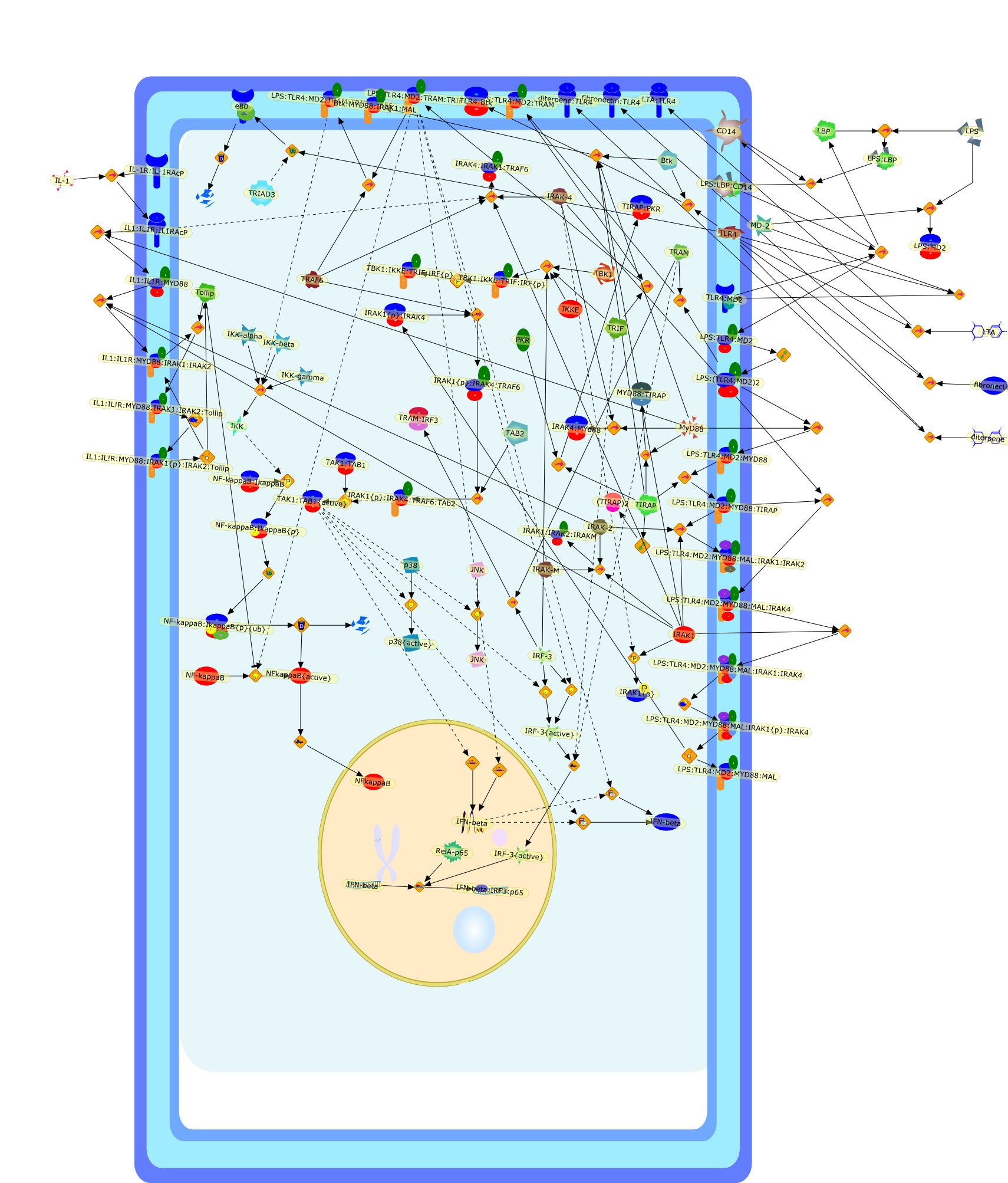{kind=link}
{kind=link}
(TIRAP)2,Btk,Btk:MYD88:IRAK1:MAL,CD14,IFN-beta,IFN-beta:IRF3:p65,IKK,IKK-alpha,IKK-beta,IKK-gamma,IKKE,IL-1,IL-1R:IL-1RAcP,IL1:IL!R:MYD88:IRAK1:IRAK2:Tollip,IL1:IL!R:MYD88:IRAK1{p}:IRAK2:Tollip,IL1:IL1R:IL1RAcP,IL1:IL1R:MYD88,IL1:IL1R:MYD88:IRAK1:IRAK2,IRAK-2,IRAK-4,IRAK-M,IRAK1,IRAK1:IRAK2:IRAKM,IRAK1{p},IRAK1{p}:IRAK4,IRAK1{p}:IRAK4:TRAF6,IRAK1{p}:IRAK4:TRAF6:TAb2,IRAK4:IRAK1:TRAF6,IRAK4:MYd88,IRF-3,IRF-3{active},JNK,LBP,LPS,LPS:(TLR4:MD2)2,LPS:LBP,LPS:LBP:CD14,LPS:MD2,LPS:TLR4:MD2,LPS:TLR4:MD2:MYD88,LPS:TLR4:MD2:MYD88:MAL,LPS:TLR4:MD2:MYD88:MAL:IRAK1:IRAK2,LPS:TLR4:MD2:MYD88:MAL:IRAK1:IRAK4,LPS:TLR4:MD2:MYD88:MAL:IRAK1{p}:IRAK4,LPS:TLR4:MD2:MYD88:MAL:IRAK4,LPS:TLR4:MD2:MYD88:TIRAP,LPS:TLR4:MD2:TRAM,LPS:TLR4:MD2:TRAM:TRIF,LPS:TLR4:MD2:TRAM:TRIF:TRAF6,LTA,LTA:TLR4,MD-2,MYD88:TIRAP,MyD88,NF-kappaB,NF-kappaB:IkappaB,NF-kappaB:IkappaB{p},NF-kappaB:IkappaB{p}{ub},NFkappaB,NFkappaB{active},PKR,RelA-p65,TAB2,TAK1:TAB1,TAK1:TAB1{active},TBK1,TBK1:IKKE:TRIF:IRF{p},TIRAP,TIRAP:PKR,TLR4,TLR4:Btk,TLR4:MD2,TRAF6,TRAM,TRAM:IRF3,TRIAD3,TRIF,Tollip,diterpene,diterpene:TLR4,e80,fibronectin,fibronectin:TLR4,p38,p38{active},protein remnants
The vertebrate innate immune system recognizes infectious non-self by employinga set of germline-encoded receptors such as nucleotide-binding oligomerisationdomain proteins (NODs) or Toll-like receptors (TLRs). These proteins areinvolved in the recognition of various microbial-derived molecules, includinglipopolysaccharide (LPS), peptidoglycan (PGN) and beta1,3-glucan. DrosophilaToll receptors are not directly dedicated to non-self recognition and insect NODorthologues have not yet been identified. Studies started more than 20 years agoand conducted on different insect models have identified other receptors onwhich invertebrate innate systems rely to sense invading microorganisms.
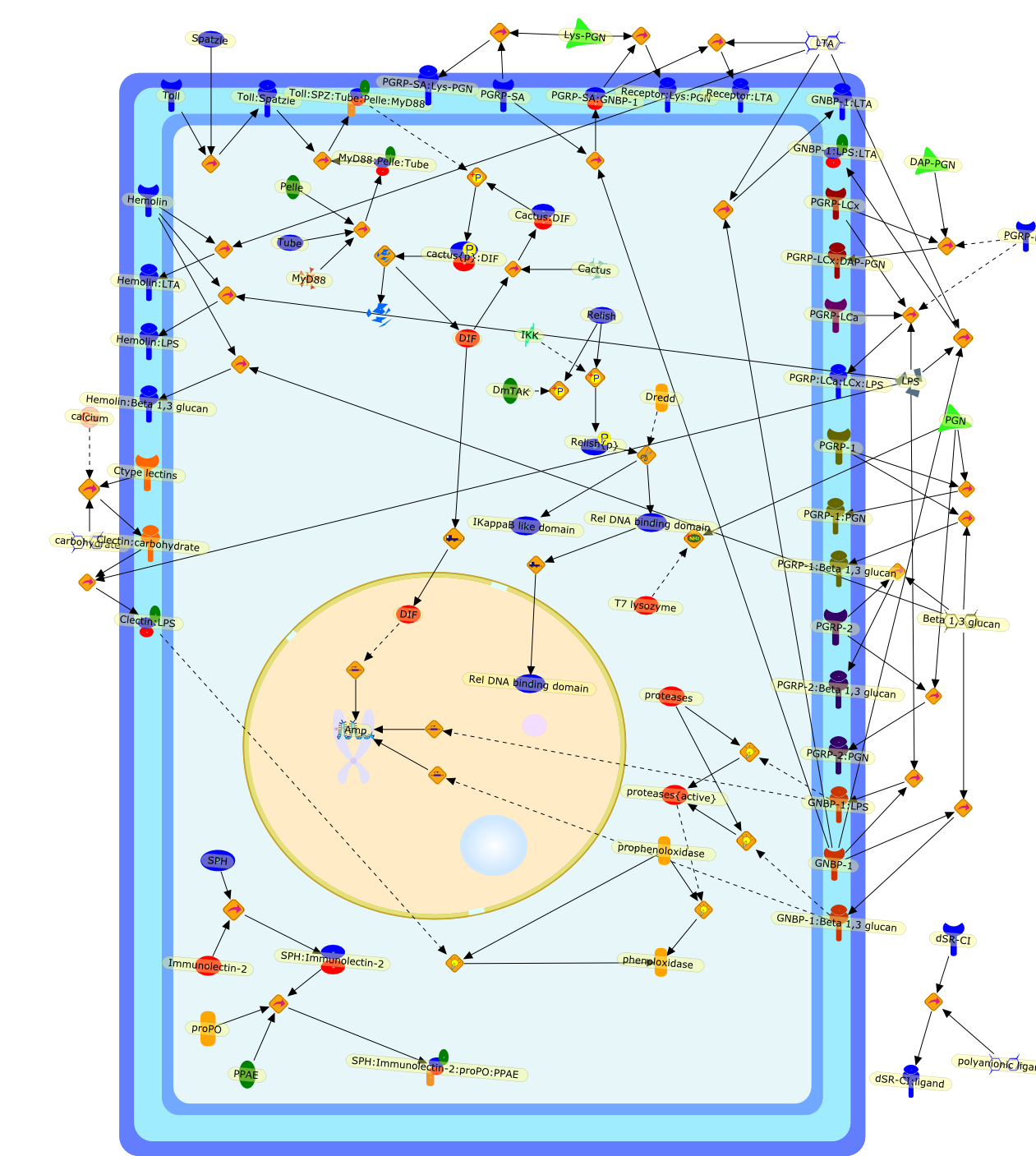{kind=link}
{kind=link}
Amp,Beta 1,3 glucan,Cactus,Cactus:DIF,Clectin:LPS,Clectin:carbohydrate,Ctype lectins,DAP-PGN,DIF,DmTAK,Dredd,GNBP-1,GNBP-1:Beta 1,3 glucan,GNBP-1:LPS,GNBP-1:LPS:LTA,GNBP-1:LTA,Hemolin,Hemolin:Beta 1,3 glucan,Hemolin:LPS,Hemolin:LTA,IKK,IKappaB like domain,Immunolectin-2,LPS,LTA,Lys-PGN,MyD88,MyD88:Pelle:Tube,PGN,PGRP-1,PGRP-1:Beta 1,3 glucan,PGRP-1:PGN,PGRP-2,PGRP-2:Beta 1,3 glucan,PGRP-2:PGN,PGRP-LCa,PGRP-LCx,PGRP-LCx:DAP-PGN,PGRP-LE,PGRP-SA,PGRP-SA:GNBP-1,PGRP-SA:Lys-PGN,PGRP:LCa:LCx:LPS,PPAE,Pelle,Receptor:LTA,Receptor:Lys:PGN,Rel DNA binding domain,Relish,Relish{p},SPH,SPH:Immunolectin-2,SPH:Immunolectin-2:proPO:PPAE,Spatzle,T7 lysozyme,Toll,Toll:SPZ:Tube:Pelle:MyD88,Toll:Spatzle,Tube,cactus{p}:DIF,calcium,carbohydrate,dSR-CI,dSR-CI:ligand,phenoloxidase,polyanionic ligands,proPO,prophenoloxidase,proteases,proteases{active},proteosome remnants
Our understanding of innate immunity in mammals has greatly expanded followingthe discovery of the family of membrane-bound receptors, called the Toll-likereceptors (TLRs). More recently, the nucleotide-binding oligomerisation domain(Nod) molecules, Nod1 and Nod2, which are cytoplasmic surveillance proteins,have also been shown to be involved in the innate immune response. These twoclasses of detection molecules, classified as "pattern recognition receptors"(PRRs), detect microbial ligands in order to initiate a defense response tofight infectious disease. These microbial ligands or "pathogen-associatedmolecular patterns" (PAMPs), detected by TLRs and Nods are often structuralcomponents of the microorganism that are not subject to much variation. Theseinclude such factors as lipopolysaccharide (LPS) and peptidoglycan from the cellwalls of bacteria. In order to understand the role of TLRs and Nod proteins ininfectious disease in vivo it is important to define the site of interactionbetween PRRs and PAMPS. Additionally, the challenge of mice deficient in thevarious PRRs in natural infection models will help to decipher the contributionof these molecules not only in the innate immune response against pathogeninfection but also how these proteins may instruct the adaptive immune responsein order to have a tailored immune response against a particular microbe.
{kind=link}
{kind=link}
C1Q,CD14,DEFB,DEFB: TLR4,GM-triDAP,GM-triDAP: Nod1,HSP: TLR4,Hsp,IFN-gamma,IFNgamma,IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma {activated},IL-12,IL-12: IL-12R,IL-12R,IRAK,IRAK [activated},IkappaB-alpha,IkappaB-alpha {ub},IkappaB-alpha{p},LBP,LPG,LPG: TLR2,LPS,LPS: CD14: MD-2: TLR4,LPS: CD14: MD-2: TLR4: MyD88,LPS:LBP,LPS:LBP:CD14,LTA,LTA: TLR2,Lp,Lp: TLR2,MD-2,MD-2: TLR4,MDP,MDP: Nod2,MyD88,NF-kappaB,NF-kappaB {nucleus},Nod,Nod {oligomer},Nod1,Nod2,PGN,PGN: TLR2,Ptx3,Ptx3: C1q,RIP2,RIP2 {activated}: NOD {oligomer},RIP2 {activated}: NOD {oligomer}: IKK-alpha:IKK-beta:IKK-gamma,RIP2 {activated}: NOD {oligomer}: IKK-alpha:IKK-beta:IKK-gamma (n),RIP2: NOD {oligomer},S. flexneri,TLR2,TLR3,TLR4,TLR5,TRAF6,TRAF6: IRAK {activated},dsRNA,dsRNA:TLR3,flagellin,flagellin: TLR5,lipoarabinomannan,lipoarabinomannan: TLR2,pro-inflammatory response genes,protein remnants,zymosan,zymosan: TLR2
Toll-like receptors (TLR) mediate recognition of several microbial products.Accumulating evidence indicates that TLR are capable of inducing distinctresponses in dendritic cells and other antigen-presenting cells, and can directT-helper cell differentiation in opposing directions. The generation of suchvaried responses is achieved through the selective utilization of adaptormolecules that link TLR to distinct signal transduction pathways. The ability ofTLR to activate and guide innate and adaptive immunity has the potential to beexploited for practical application that may lead to the development of moresuccessful immunotherapies and vaccination strategies. A review of recentliterature, unpublished observations, and future challenges is presented here.
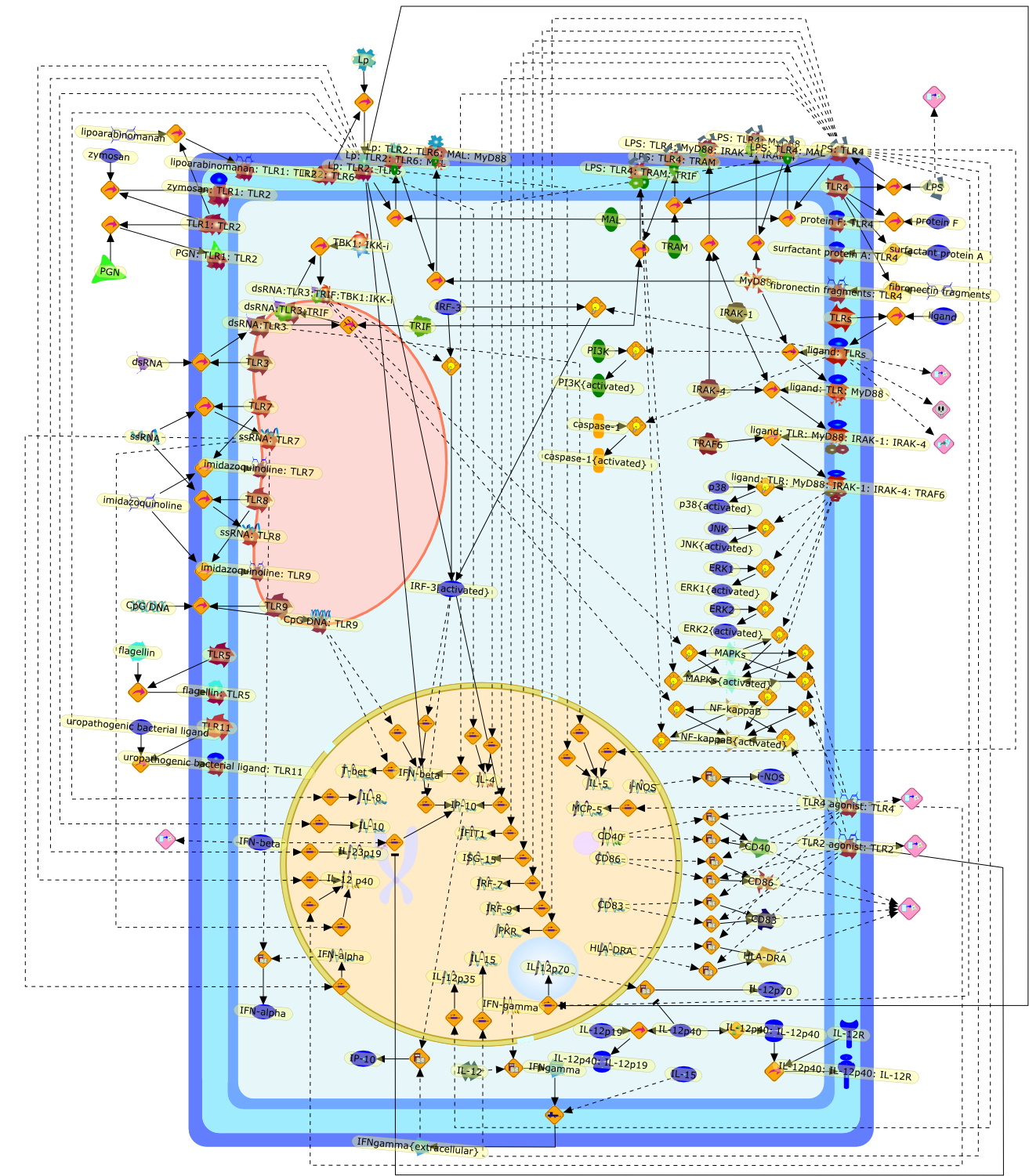{kind=link}
{kind=link}
CD40,CD83,CD86,CpG DNA,CpG DNA: TLR9,ERK1,ERK1{activated},ERK2,ERK2{activated},HLA-DRA,IFIT1,IFN-alpha,IFN-beta,IFN-gamma,IFNgamma,IFNgamma{extracellular},IL-10,IL-12,IL-12 p40,IL-12R,IL-12p19,IL-12p35,IL-12p40,IL-12p40: IL-12p19,IL-12p40: IL-12p40,IL-12p40: IL-12p40: IL-12R,IL-12p70,IL-15,IL-23p19,IL-4,IL-5,IL-8,IP-10,IRAK-1,IRAK-4,IRF-3,IRF-3[activated},IRF-7,IRF-9,ISG-15,JNK,JNK{activated},LPS,LPS: TLR4,LPS: TLR4: MAL,LPS: TLR4: MyD88,LPS: TLR4: MyD88: IRAK-1: IRAK-4,LPS: TLR4: TRAM,LPS: TLR4: TRAM: TRIF,Lp,Lp: TLR2: TLR6,Lp: TLR2: TLR6: MAL,Lp: TLR2: TLR6: MAL: MyD88,MAL,MAPKs,MAPKs{activated},MCP-5,MyD88,NF-kappaB,NF-kappaB{activated},PGN,PGN: TLR1: TLR2,PI3K,PI3K{activated},PKR,T-bet,TBK1: IKK-i,TLR11,TLR1: TLR2,TLR2 agonist: TLR2,TLR2: TLR6,TLR3,TLR4,TLR4 agonist: TLR4,TLR5,TLR7,TLR8,TLR9,TLRs,TRAF6,TRAM,TRIF,caspase-1,caspase-1{activated},dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:TBK1:IKK-i,fibronectin fragments,fibronectin fragments: TLR4,flagellin,flagellin: TLR5,i-NOS,imidazoquinoline,imidazoquinoline: TLR7,imidazoquinoline: TLR9,ligand,ligand: TLR: MyD88,ligand: TLR: MyD88: IRAK-1: IRAK-4,ligand: TLR: MyD88: IRAK-1: IRAK-4: TRAF6,ligand: TLRs,lipoarabinomanan,lipoarabinomanan: TLR1: TLR2,p38,p38{activated},protein F,protein F: TLR4,ssRNA,ssRNA: TLR7,ssRNA: TLR8,surfactant protein A,surfactant protein A: TLR4,uropathogenic bacterial ligand,uropathogenic bacterial ligand: TLR11,zymosan,zymosan: TLR1: TLR2
PI3K (phosphoinositide 3-kinase) I(A) family members contain a regulatorysubunit and a catalytic subunit. The p110delta catalytic subunit is expressedpredominantly in haematopoietic cells. There, among other functions, itregulates antigen receptor-mediated responses. Using mice deficient in thep110delta subunit of PI3K, we investigated the role of this subunit in LPS(lipopolysaccharide)-induced B cell responses, which are mediated by Toll-likereceptor 4 and RP105. After injection of DNP-LPS (where DNP stands for2,4-dinitrophenol), p110delta(-/-) mice produced reduced levels of DNP-specificIgM and IgG when compared with wild-type mice. In vitro, the proliferation andup-regulation of surface activation markers such as CD86 and CD25 induced by LPSand an antibody against RP105 were decreased. We analysed the activation stateof key components of the LPS pathway in B cells to determine whether there was adefect in signalling in p110delta(-/-) B cells. They showed normalextracellular-signal-regulated kinase phosphorylation, but anti-RP105-inducedprotein kinase B, IkappaB (inhibitor of nuclear factor kappaB) and c-JunN-terminal kinase activation was severely reduced. This demonstrates that thep110delta subunit of PI3K is involved in the LPS response in B cells and mayrepresent a link between the innate and the adaptive immune system.
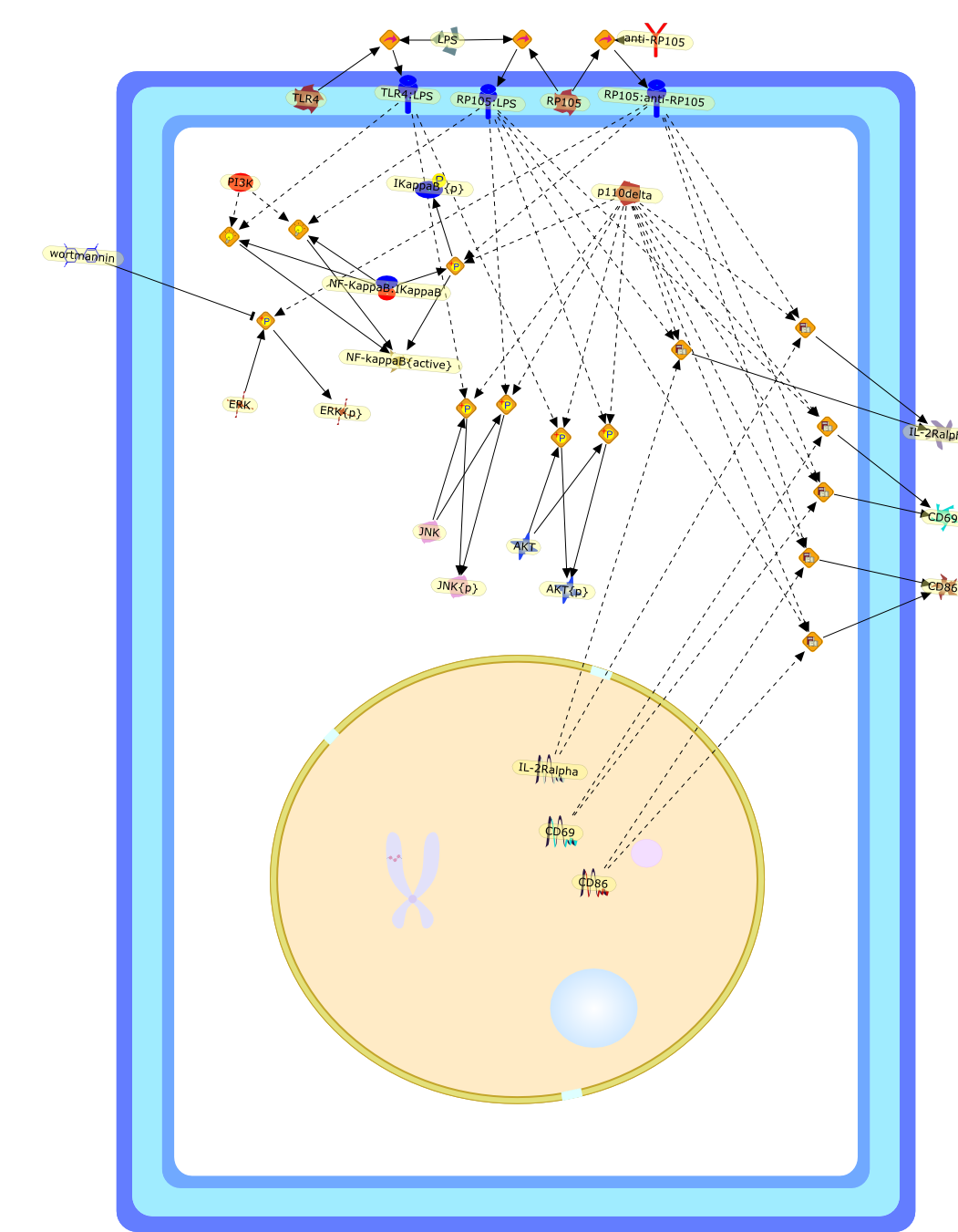{kind=link}
{kind=link}
AKT,AKT{p},CD69,CD86,ERK,ERK{p},IKappaB {p},IL-2Ralpha,JNK,JNK{p},LPS,NF-KappaB:IKappaB,NF-kappaB{active},PI3K,RP105,RP105:LPS,RP105:anti-RP105,TLR4,TLR4:LPS,anti-RP105,p110delta,wortmannin
SAP and EAT-2 define a new class of adaptor proteins composed almost exclusivelyof a Src homology 2 (SH2) domain. By way of their SH2 domain, SAP-like adaptorsinteract with tyrosine-based motifs in the cytoplasmic region of SLAM-relatedreceptors, a family of immune cell-specific molecules involved inimmunoregulation. Recent findings indicate that SAP is required for thefunctions of SLAM family receptors, as a consequence of its ability to promoterecruitment of Src-related protein tyrosine kinase FynT and allow SLAM-relatedreceptors to transduce tyrosine phosphorylation signals. SAP is mutated inX-linked lymphoproliferative (XLP) syndrome, a rare inherited human diseasecharacterized by an deregulated immune response to Epstein-Barr virus infection.Several lines of evidence indicate that defects in the activities ofSLAM-related receptors caused by SAP deficiency account for the immunedysfunctions associated with XLP.
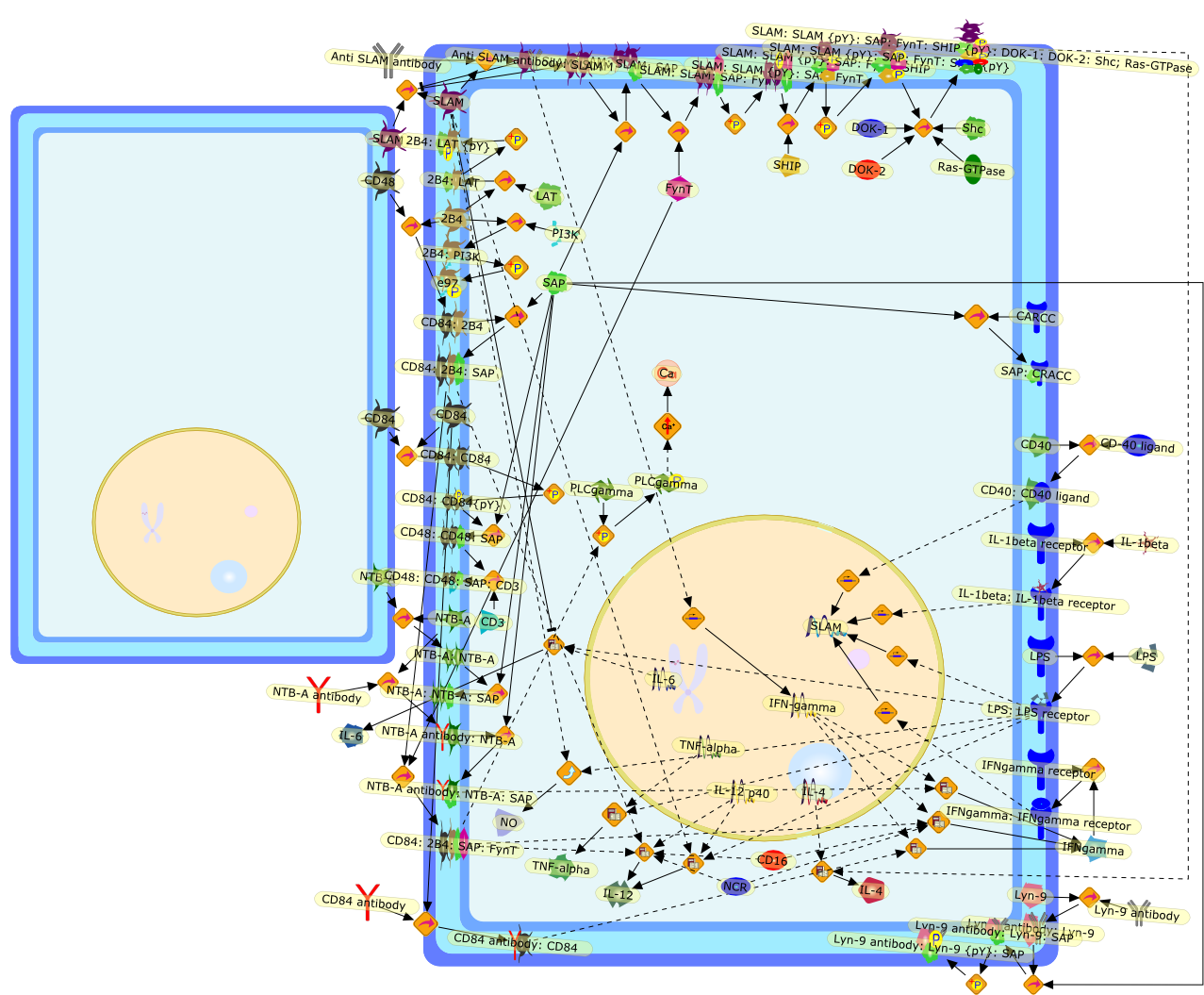{kind=link}
{kind=link}
2B4,2B4: LAT,2B4: LAT {pY},2B4: PI3K,Anti SLAM antibody,Anti SLAM antibody: SLAM,CARCC,CD-40 ligand,CD16,CD3,CD40,CD40: CD40 ligand,CD48,CD48: CD48: SAP,CD48: CD48: SAP: CD3,CD84,CD84 antibody,CD84 antibody: CD84,CD84: 2B4,CD84: 2B4: SAP,CD84: 2B4: SAP: FynT,CD84: CD84,CD84: CD84{pY},Ca,DOK-1,DOK-2,FynT,IFN-gamma,IFNgamma,IFNgamma receptor,IFNgamma: IFNgamma receptor,IL-12,IL-12 p40,IL-1beta,IL-1beta receptor,IL-1beta: IL-1beta receptor,IL-4,IL-6,LAT,LPS,LPS: LPS receptor,Lyn-9,Lyn-9 antibody,Lyn-9 antibody: Lyn-9,Lyn-9 antibody: Lyn-9 {pY}: SAP,Lyn-9 antibody: Lyn-9: SAP,NCR,NO,NTB-A,NTB-A antibody,NTB-A antibody: NTB-A,NTB-A antibody: NTB-A: SAP,NTB-A: NTB-A,NTB-A: NTB-A: SAP,PI3K,PLCgamma,Ras-GTPase,SAP,SAP: CRACC,SHIP,SLAM,SLAM: SLAM,SLAM: SLAM {pY}: SAP: FynT,SLAM: SLAM {pY}: SAP: FynT: SHIP,SLAM: SLAM {pY}: SAP: FynT: SHIP {pY},SLAM: SLAM {pY}: SAP: FynT: SHIP {pY}: DOK-1: DOK-2: Shc; Ras-GTPase,SLAM: SLAM: SAP,SLAM: SLAM: SAP: FynT,Shc,TNF-alpha,e97
Cerebral ischemia triggers acute inflammation, which exacerbates primary braindamage. Activation of the innate immune system is an important component of thisinflammatory response. Inflammation occurs through the action of proinflammatorycytokines, such as TNF, IL-1 beta and IL-6, that alter blood flow and increasevascular permeability, thus leading to secondary ischemia and accumulation ofimmune cells in the brain. Production of these cytokines is initiated bysignaling through Toll-like receptors (TLRs) that recognize host-derivedmolecules released from injured tissues and cells. Recently, great strides havebeen made in understanding the regulation of the innate immune system,particularly the signaling mechanisms of TLRs. Negative feedback inhibitors ofTLRs and inflammatory cytokines have now been identified and characterized. Itis also evident that lipid rafts exist in membranes and play a role inreceptor-mediated inflammatory signaling events. In the present review, usingthis newly available large body of knowledge, we take a fresh look at studies ofischemic tolerance. Based on this analysis, we recognize a striking similaritybetween ischemic tolerance and endotoxin tolerance, an immune suppressive statecharacterized by hyporesponsiveness to lipopolysaccharide (LPS). In view of thisanalogy, and considering recent discoveries related to molecular mechanisms ofendotoxin tolerance, we postulate that inhibition of TLR and proinflammatorycytokine signaling contributes critically to ischemic tolerance in the brain andother organs. Ischemic tolerance is a protective mechanism induced by a varietyof preconditioning stimuli. Tolerance can be established with two temporalprofiles: (i) a rapid form in which the trigger induces tolerance to ischemiawithin minutes and (ii) a delayed form in which development of protection takesseveral hours or days and requires de-novo protein synthesis. The rapid form oftolerance is achieved by direct interference with membrane fluidity, causingdisruption of lipid rafts leading to inhibition of TLR/cytokine signalingpathways. In the delayed form of tolerance, the preconditioning stimulus firsttriggers the TLR/cytokine inflammatory pathways, leading not only toinflammation but also to simultaneous upregulation of feedback inhibitors ofinflammation. These inhibitors, which include signaling inhibitors, decoyreceptors, and anti-inflammatory cytokines, reduce the inflammatory response toa subsequent episode of ischemia. This novel interpretation of the molecularmechanism of ischemic tolerance highlights new avenues for future investigationinto the prevention and treatment of stroke and related diseases.
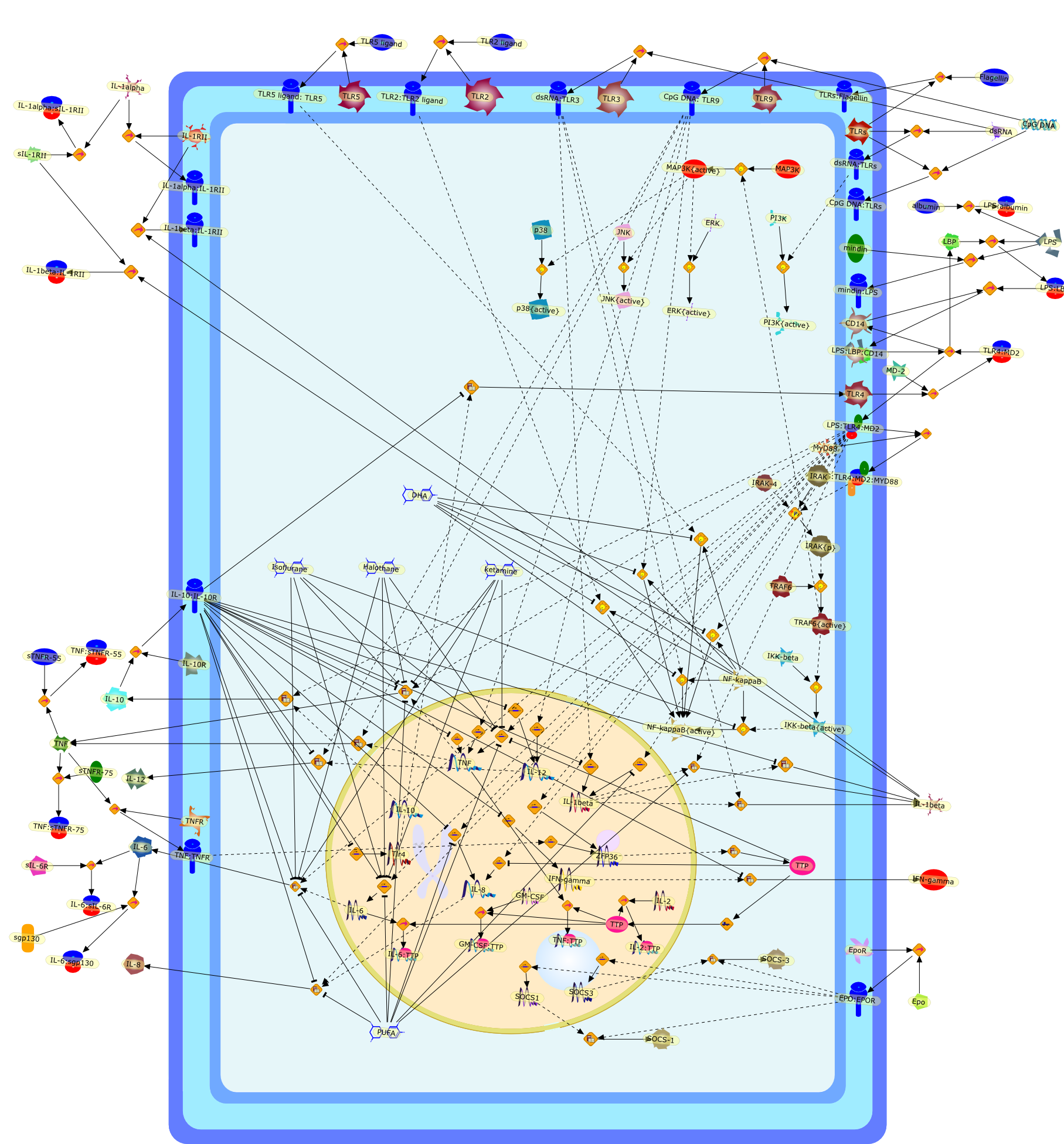{kind=link}
{kind=link}
CD14,CpG DNA,CpG DNA: TLR9,CpG DNA:TLRs,DHA,EPO:EPOR,ERK,ERK{active},Epo,EpoR,Flagellin,GM-CSF,GM-CSF:TTP,Halothane,IFN-gamma,IKK-beta,IKK-beta{active},IL-10,IL-10:IL-10R,IL-10R,IL-12,IL-1RII,IL-1alpha,IL-1alpha:IL-1RII,IL-1alpha:sIL-1RII,IL-1beta,IL-1beta:IL-1RII,IL-2,IL-2:TTP,IL-6,IL-6:TTP,IL-6:sIL-6R,IL-6:sgp130,IL-8,IRAK,IRAK-4,IRAK{p},Isoflurane,JNK,JNK{active},LBP,LPS,LPS:LBP,LPS:LBP:CD14,LPS:TLR4:MD2,LPS:TLR4:MD2:MYD88,LPS:albumin,MAP3K,MAP3K{active},MD-2,MyD88,NF-kappaB,NF-kappaB{active},PI3K,PI3K{active},PUFA,SOCS-1,SOCS-3,SOCS1,SOCS3,TLR2,TLR2 ligand,TLR2:TLR2 ligand,TLR3,TLR4,TLR4:MD2,TLR5,TLR5 ligand,TLR5 ligand: TLR5,TLR9,TLRs,TLRs:Flagellin,TNF,TNF:TNFR,TNF:TTP,TNF:sTNFR-55,TNF:sTNFR-75,TNFR,TRAF6,TRAF6{active},TTP,Tlr4,ZFP36,albumin,dsRNA,dsRNA:TLR3,dsRNA:TLRs,ketamine,mindin,mindin:LPS,p38,p38{active},sIL-1RII,sIL-6R,sTNFR-55,sTNFR-75,sgp130
Produced in response to a variety of pathogenic organisms, interleukin (IL)-12and IL-23 are key immunoregulatory cytokines that coordinate innate and adaptiveimmune responses. These dimeric cytokines share a subunit, designated p40, andbind to a common receptor chain, IL-12R beta 1. The receptor for IL-12 iscomposed of IL-12R beta 1 and IL-12R beta 2, whereas IL-23 binds to a receptorcomposed of IL-12R beta 1 and IL-23R. Both cytokines activate the Janus kinasesTyk2 and Jak2, the transcription factor signal transducer and activator oftranscription 4 (STAT4), as well as other STATs. A major action of IL-12 is topromote the differentiation of naive CD4+ T cells into T-helper (Th) 1 cells,which produce interferon (IFN)-gamma, and deficiency of IL-12, IL-12R subunitsor STAT4 is similar in many respects. In contrast, IL-23 promotes end-stageinflammation. Targeting IL-12, IL-23, and their downstream signaling elementswould therefore be logical strategies for the treatment of immune-mediateddiseases.
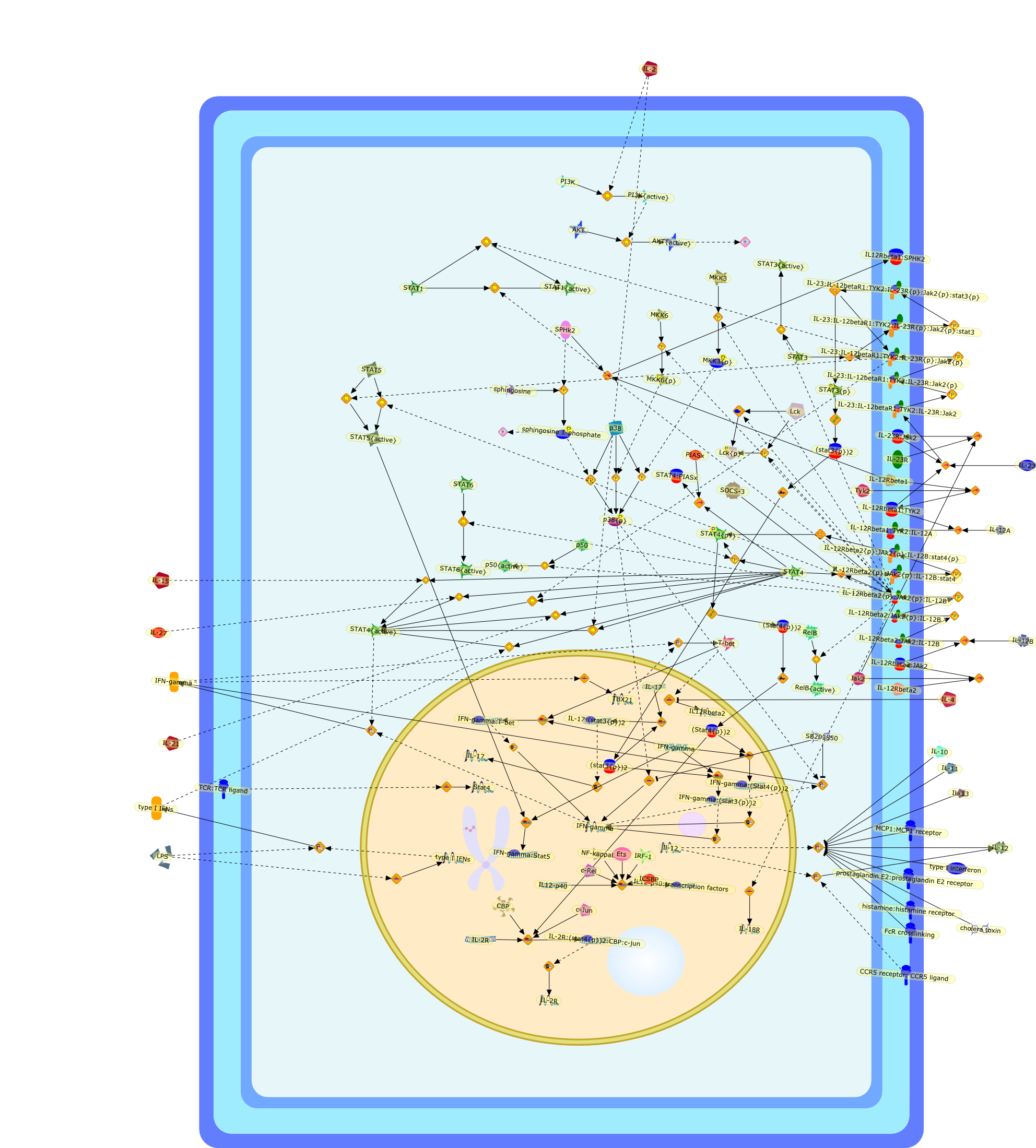{kind=link}
{kind=link}
(Stat4{p})2,(stat3{p})2,AKT,AKT{active},CBP,CCR5 receptor: CCR5 ligand,Ets,FcR crosslinking,ICSBP,IFN-gamma,IFN-gamma:(Stat4{p})2,IFN-gamma:(stat3{p})2,IFN-gamma:Stat5,IFN-gamma:T-bet,IL-10,IL-11,IL-12,IL-12A,IL-12B,IL-12Rbeta1,IL-12Rbeta1:TYK2,IL-12Rbeta1:TYK2:IL-12A,IL-12Rbeta2,IL-12Rbeta2:JAk2,IL-12Rbeta2:JAk2:IL-12B,IL-12Rbeta2:JAk2{p}:IL-12B,IL-12Rbeta2{p}:JAk2{p}:IL-12B,IL-12Rbeta2{p}:JAk2{p}:IL-12B:stat4,IL-12Rbeta2{p}:JAk2{p}:IL-12B:stat4{p},IL-13,IL-15,IL-17,IL-17:(stat3{p})2,IL-18R,IL-2,IL-21,IL-23,IL-23:IL-12betaR1:TYK2:IL-23R:Jak2,IL-23:IL-12betaR1:TYK2:IL-23R:Jak2{p},IL-23:IL-12betaR1:TYK2:IL-23R{p}:Jak2{p},IL-23:IL-12betaR1:TYK2:IL-23R{p}:Jak2{p}:stat3,IL-23:IL-12betaR1:TYK2:IL-23R{p}:Jak2{p}:stat3{p},IL-23R,IL-23R:Jak2,IL-27,IL-2R,IL-2R:(stat4{p})2:CBP:c-jun,IL-4,IL12-p40,IL12-p40:transcription factors,IL12Rbeta1:SPHK2,IL12Rbeta2,IRF-1,Il-12,Jak2,LPS,Lck,Lck{p},MCP1:MCP1 receptor,MKK3,MKK3{p},MKK6,MKK6{p},NF-kappaB,PI3K,PI3K{active},PIASx,RelB,RelB{active},SB203850,SOCS-3,SPHk2,STAT1,STAT1{active},STAT3,STAT3{active},STAT3{p},STAT4,STAT4:PIASx,STAT4{active},STAT4{pY},STAT5,STAT5{active},STAT6,STAT6{active},Stat4,T-bet,TBX21,TCR:TCR ligand,Tyk2,c-Jun,c-Rel,cholera toxin,histamine:histamine receptor,p38,p38{p},p50,p50{active},prostaglandin E2:prostaglandin E2 receptor,sphingosine,sphingosine 1-phosphate,type I IFNs,type I interferon
The innate immune system senses bacteria in the environment and defends againstinfection. Here we will discuss two types of sensor protein families. The plasmamembrane receptors that comprise the Toll-like receptor (TLRs) family and theintracellular proteins termed NOD1 and NOD2. These proteins directly bindbacterial products such as lipopolysaccharides (LPS), peptidoglycan fragments,bacterial DNA, and receptor binding leads to intracellular signaling and geneexpression. TLR signaling involves members of the MyD88 family of adaptorproteins. In contrast NOD1 or NOD2 utilize pathways that do not depend on theMyD88 family members.
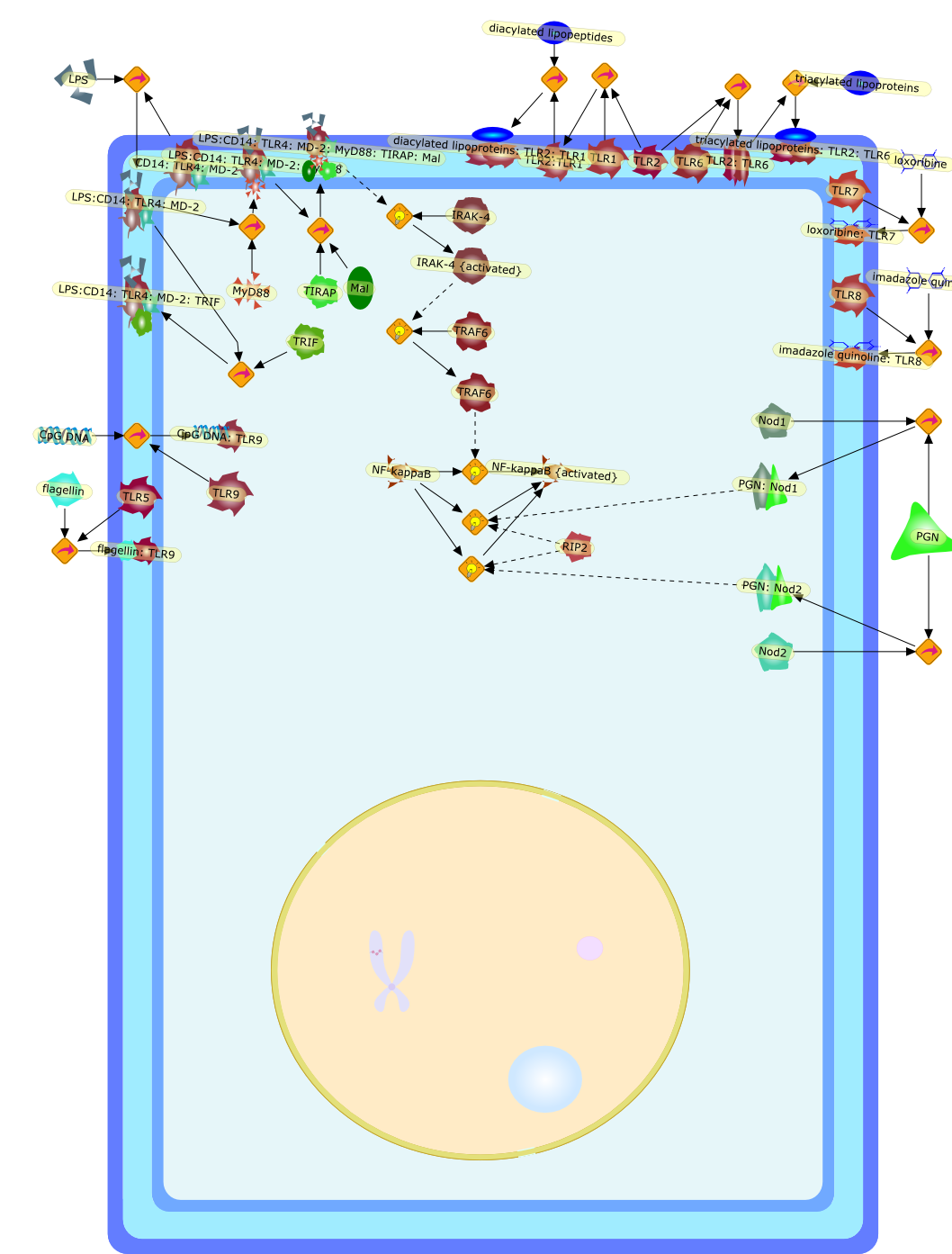{kind=link}
{kind=link}
CD14: TLR4: MD-2,CpG DNA,CpG DNA: TLR9,IRAK-4,IRAK-4 {activated},LPS,LPS:CD14: TLR4: MD-2,LPS:CD14: TLR4: MD-2: MyD88,LPS:CD14: TLR4: MD-2: MyD88: TIRAP: Mal,LPS:CD14: TLR4: MD-2: TRIF,Mal,MyD88,NF-kappaB,NF-kappaB {activated},Nod1,Nod2,PGN,PGN: Nod1,PGN: Nod2,RIP2,TIRAP,TLR1,TLR2,TLR2: TLR6,TLR2:TLR1,TLR5,TLR6,TLR7,TLR8,TLR9,TRAF6,TRIF,diacylated lipopeptides,diacylated lipoproteins: TLR2: TLR1,flagellin,flagellin: TLR9,imadazole quinoline,imadazole quinoline: TLR8,loxoribine,loxoribine: TLR7,triacylated lipoproteins,triacylated lipoproteins: TLR2: TLR6
The Toll-IL-1 receptor (TIR) superfamily, defined by the presence of anintracellular TIR domain, initiates innate immunity via NF-kappaB activation,leading to production of proinflammatory cytokines. ST2 is a member of the TIRfamily that does not activate NF-kappaB and has been suggested as an importanteffector molecule of type 2 T helper cell responses. We have recentlydemonstrated that the membrane bound form of ST2 (ST2L) negatively regulatedIL-1RI and TLR4 but not TLR3 signaling by sequestrating the adaptors MyD88 andMal. In contrast to wild-type mice, ST2 deficient mice failed to developendotoxin tolerance. Thus, ST2 suppresses IL-1R and TLR4 signaling via MyD88-and Mal-dependent pathways and modulates innate immunity. The results provide amolecular explanation for the role of ST2 in T(H)2 responses since inhibition ofTLRs will promote a T(H)2 response and also identify ST2 as a key regulator ofendotoxin tolerance.
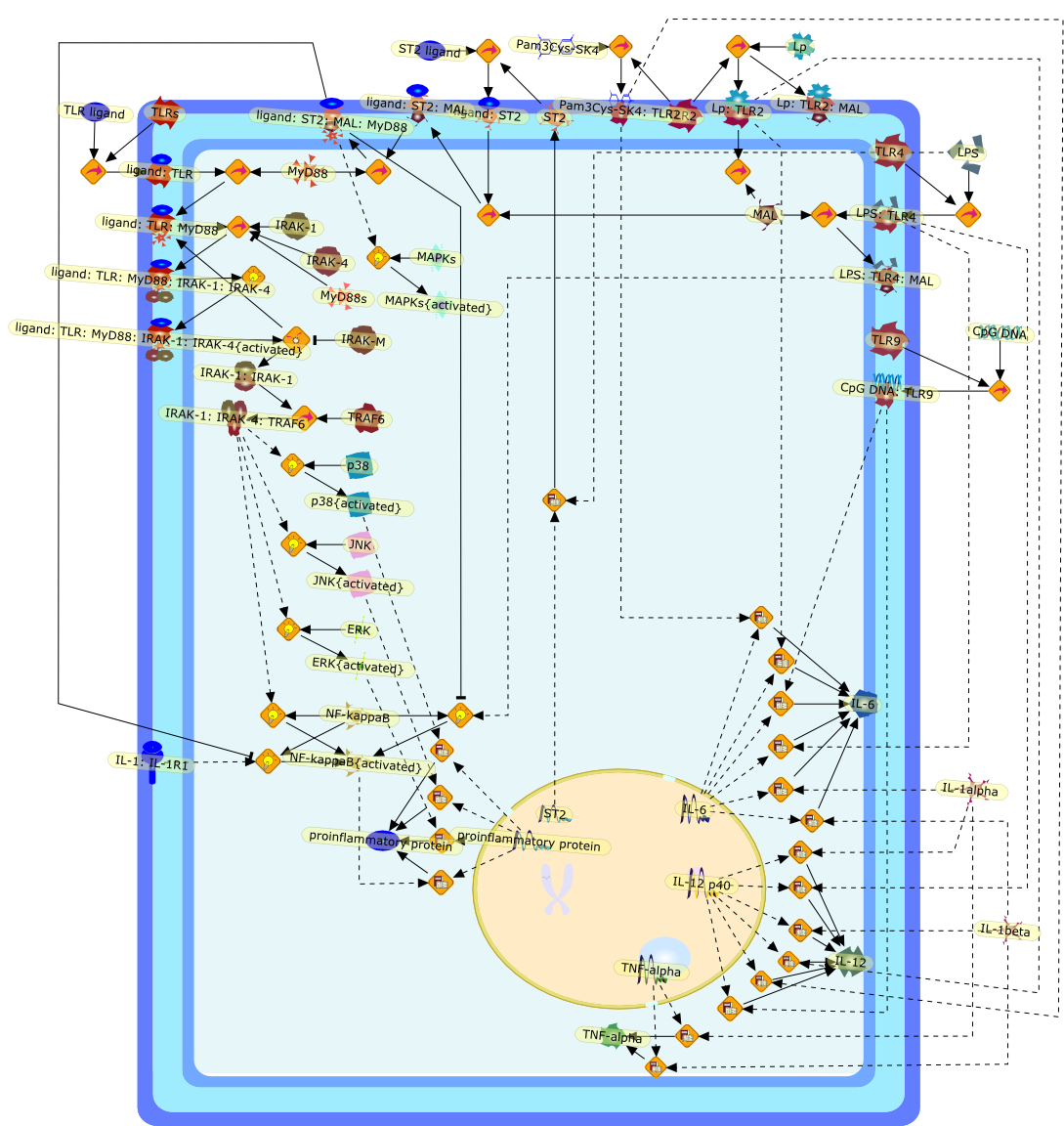{kind=link}
{kind=link}
CpG DNA,CpG DNA: TLR9,ERK,ERK{activated},IL-12,IL-12 p40,IL-1: IL-1R1,IL-1alpha,IL-1beta,IL-6,IRAK-1,IRAK-1: IRAK-1,IRAK-1: IRAK-4: TRAF6,IRAK-4,IRAK-M,JNK,JNK{activated},LPS,LPS: TLR4,LPS: TLR4: MAL,Lp,Lp: TLR2,Lp: TLR2: MAL,MAL,MAPKs,MAPKs{activated},MyD88,MyD88s,NF-kappaB,NF-kappaB{activated},Pam3Cys-SK4,Pam3Cys-SK4: TLR2,ST2,ST2 ligand,TLR ligand,TLR2,TLR4,TLR9,TLRs,TNF-alpha,TRAF6,ligand: ST2,ligand: ST2: MAL,ligand: ST2: MAL: MyD88,ligand: TLR,ligand: TLR: MyD88,ligand: TLR: MyD88: IRAK-1: IRAK-4,ligand: TLR: MyD88: IRAK-1: IRAK-4{activated},p38,p38{activated},proinflammatory protein
Innate immune receptors recognize microorganism-specific motifs. One suchreceptor-ligand complex is formed between the mammalian Toll-like receptor 4(TLR4)-MD2-CD14 complex and bacterial lipopolysaccharide (LPS). Recent researchindicates that there is significant phylogenetic and individual diversity inTLR4-mediated responses. In addition, the diversity of LPS structures and thedifferential recognition of these structures by TLR4 have been associated withseveral bacterial diseases. This review will examine the hypothesis that thevariability of bacterial ligands such as LPS and their innate immune receptorsis an important factor in determining the outcome of infectious disease.
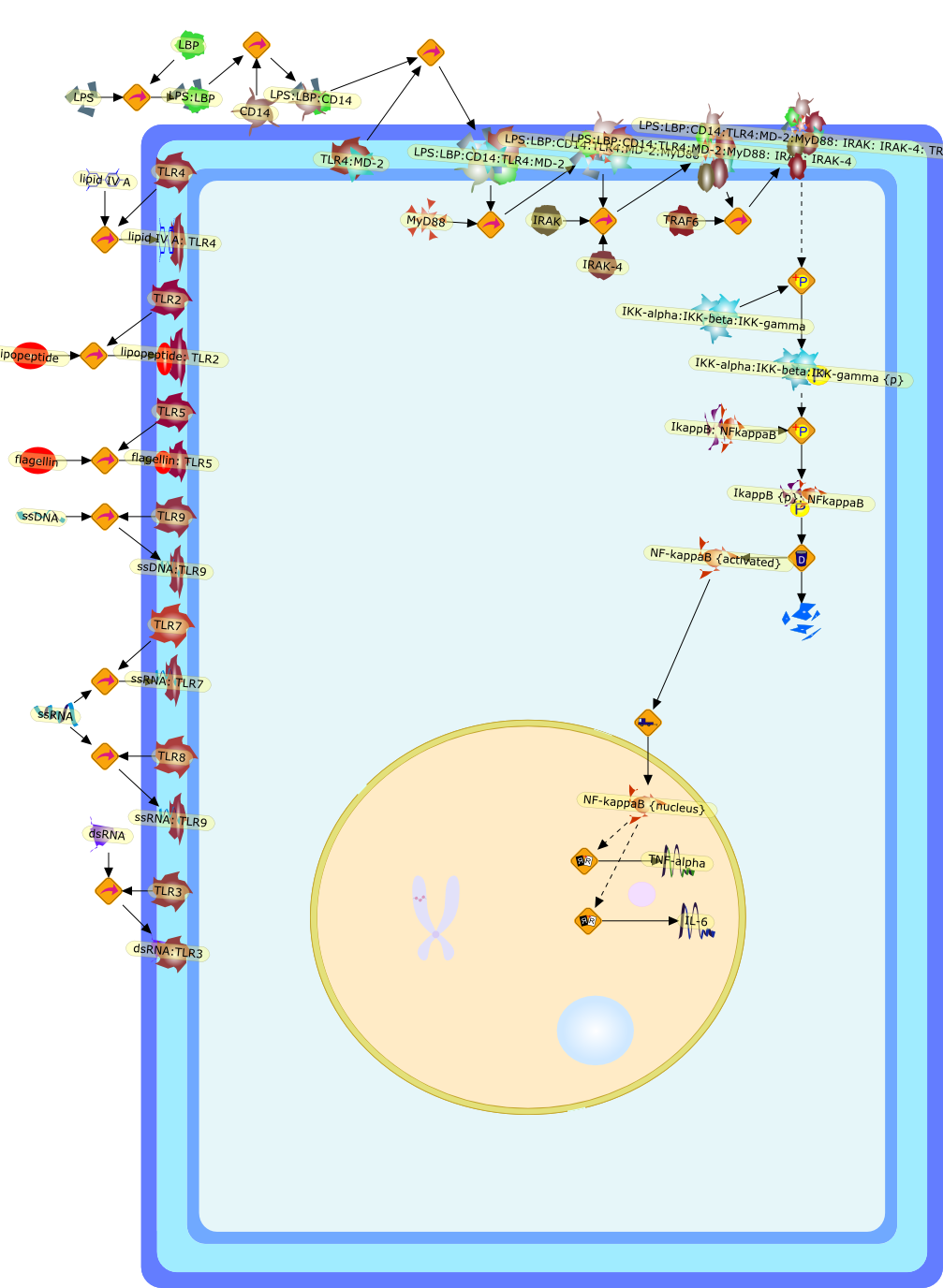{kind=link}
{kind=link}
CD14,IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma {p},IL-6,IRAK,IRAK-4,IkappB {p}: NFkappaB,IkappB: NFkappaB,IkappaB {degraded},LBP,LPS,LPS:LBP,LPS:LBP:CD14,LPS:LBP:CD14:TLR4:MD-2,LPS:LBP:CD14:TLR4:MD-2:MyD88,LPS:LBP:CD14:TLR4:MD-2:MyD88: IRAK: IRAK-4,LPS:LBP:CD14:TLR4:MD-2:MyD88: IRAK: IRAK-4: TRAF6,MyD88,NF-kappaB {activated},NF-kappaB {nucleus},TLR2,TLR3,TLR4,TLR4:MD-2,TLR5,TLR7,TLR8,TLR9,TNF-alpha,TRAF6,dsRNA,dsRNA:TLR3,flagellin,flagellin: TLR5,lipid IV A,lipid IV A: TLR4,lipopeptide,lipopeptide: TLR2,ssDNA,ssDNA:TLR9,ssRNA,ssRNA: TLR7,ssRNA: TLR9
Toll-like receptor (TLR) 3 and 4 mediate the expression of many genes, includingNF-kappaB- and interferon-regulatory factor (IRF)-3/interferon (IFN)-induciblegenes, in macrophages and dendritic cells (DCs) in response to their ligandstimuli, polyI:C and lipopolysaccharide (LPS). Toll-IL-1 receptor homologydomain (TIR)-containing adapter molecule 1 (TICAM-1) facilitates expression ofIFN-inducible genes via TLR3. Although MyD88 and Mal/TIRAP adapters functiondownstream of TLR4, they barely induce IFN-beta. In addition, DC maturation aswell as IFN-beta induction are largely independent of MyD88 and Mal/TIRAP.TICAM-1 is the functional adapter for both TLR3 and TLR4 that induces type 1 IFNand MyD88-independent DC maturation. In LPS-mediated TLR4 activation, a complexof TICAM-1 and an additional TLR4-binding adapter serves as the adapter. Wenamed this TLR4-TICAM-1-bridging adapter TICAM-2. Our results reveal the detailsof MyD88-independent pathways which separately recruit the distinct adaptersdownstream of TLR3 and TLR4 and variations of the TLR output are in partregulated by the two additional adapters in DCs.
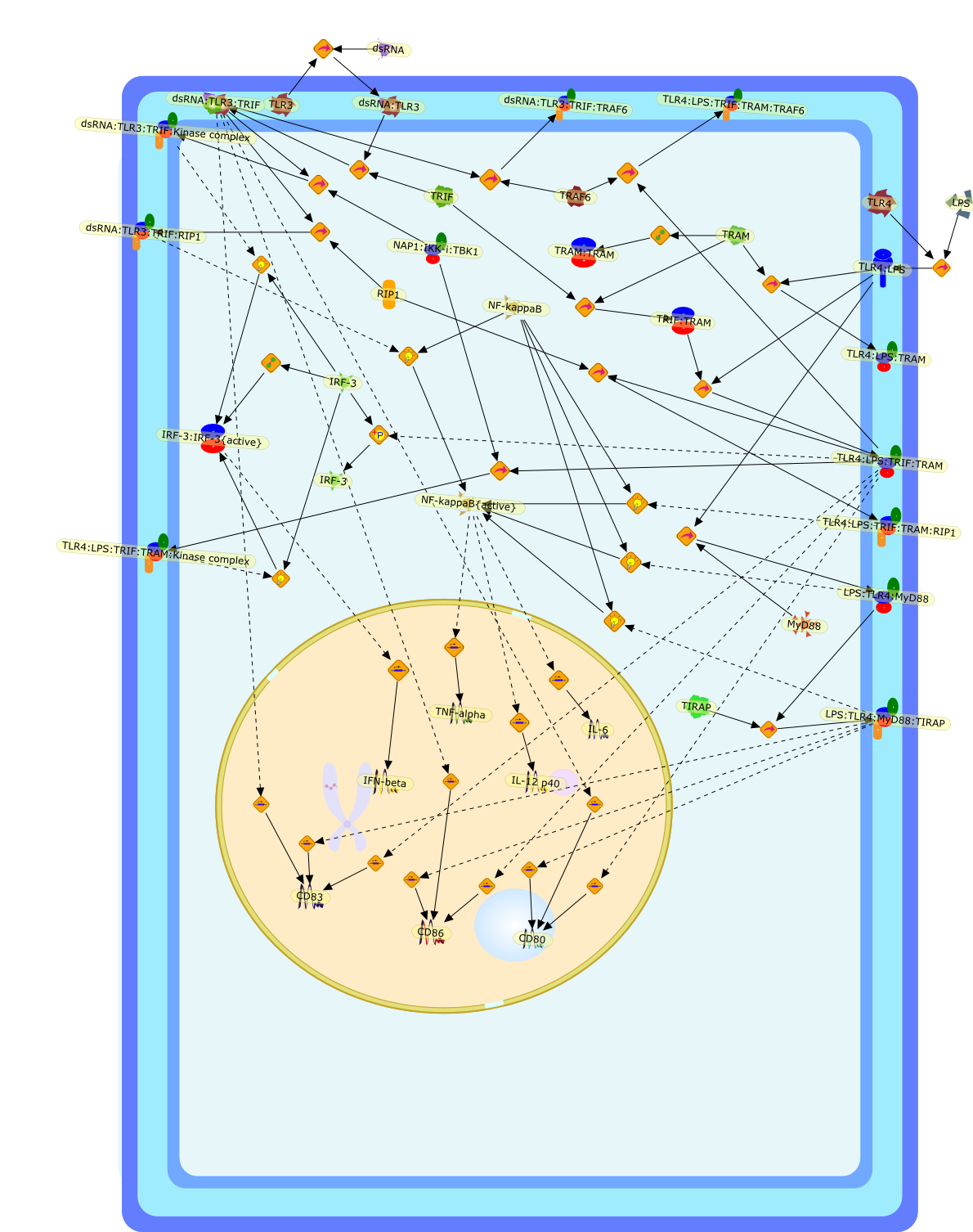{kind=link}
{kind=link}
CD80,CD83,CD86,IFN-beta,IL-12 p40,IL-6,IRF-3,IRF-3:IRF-3{active},LPS,LPS:TLR4:MyD88,LPS:TLR4:MyD88:TIRAP,MyD88,NAP1:IKK-i:TBK1,NF-kappaB,NF-kappaB{active},RIP1,TIRAP,TLR3,TLR4,TLR4:LPS,TLR4:LPS:TRAM,TLR4:LPS:TRIF:TRAM,TLR4:LPS:TRIF:TRAM:Kinase complex,TLR4:LPS:TRIF:TRAM:RIP1,TLR4:LPS:TRIF:TRAM:TRAF6,TNF-alpha,TRAF6,TRAM,TRAM:TRAM,TRIF,TRIF:TRAM,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:Kinase complex,dsRNA:TLR3:TRIF:RIP1,dsRNA:TLR3:TRIF:TRAF6
Toll-like receptors (TLRs) belong to the Toll-interleukin 1 receptorsuperfamily, which is defined by a common intracellular Toll-IL-1 receptor (TIR)domain. A group of TIR domain containing adaptors (MyD88, TIRAP, TRIF and TRAM),are differentially recruited to the Toll-IL-1 receptors, contributing to thespecificity of signaling. The IL-1 mediated signaling pathway serves as a"prototype" for other family members. Genetic and biochemical studies revealthat IL-1R uses adaptor molecule MyD88 to mediate a very complex pathway,involving a cascade of kinases organized by multiple adapter molecules intosignaling complexes, leading to activation of the transcription factor NFkappaB.Several Toll-like receptors utilize variations of the "prototype" pathway byemploying different adaptor molecules. Double-stranded RNA triggered,TLR3-mediated signaling is independent of MyD88, IRAK4, and IRAK. The adaptermolecule TRIF is utilized by TLR3 to mediate the activation of NFkappaB andIRF3. LPS-induced, TLR4-mediated signaling employs multiple TIR-domaincontaining adaptors, MyD88/TIRAP to mediate NFkappaB activation, TRIF/TRAM forIRF3 activation. Recent studies have also begun to unravel how these pathwaysare negatively regulated. SIGIRR (also known as TIR8), a member of TIRsuperfamily that does not activate the transcription factors NFkappaB and IRF3,instead negatively modulates responses. Cells from SIGIRR-null mice showenhanced activation in response to either IL-1 or certain Toll ligands. Inaddition to SIGIRR, several other negative regulators have been shown to inhibitthe TIR signaling, including ST2, IRAKM, MyD88s, SOCS1, and Triad3A. Thecoordinated positive and negative regulation of the TIR signaling ensures theappropriate modulation of the innate and inflammatory responses.
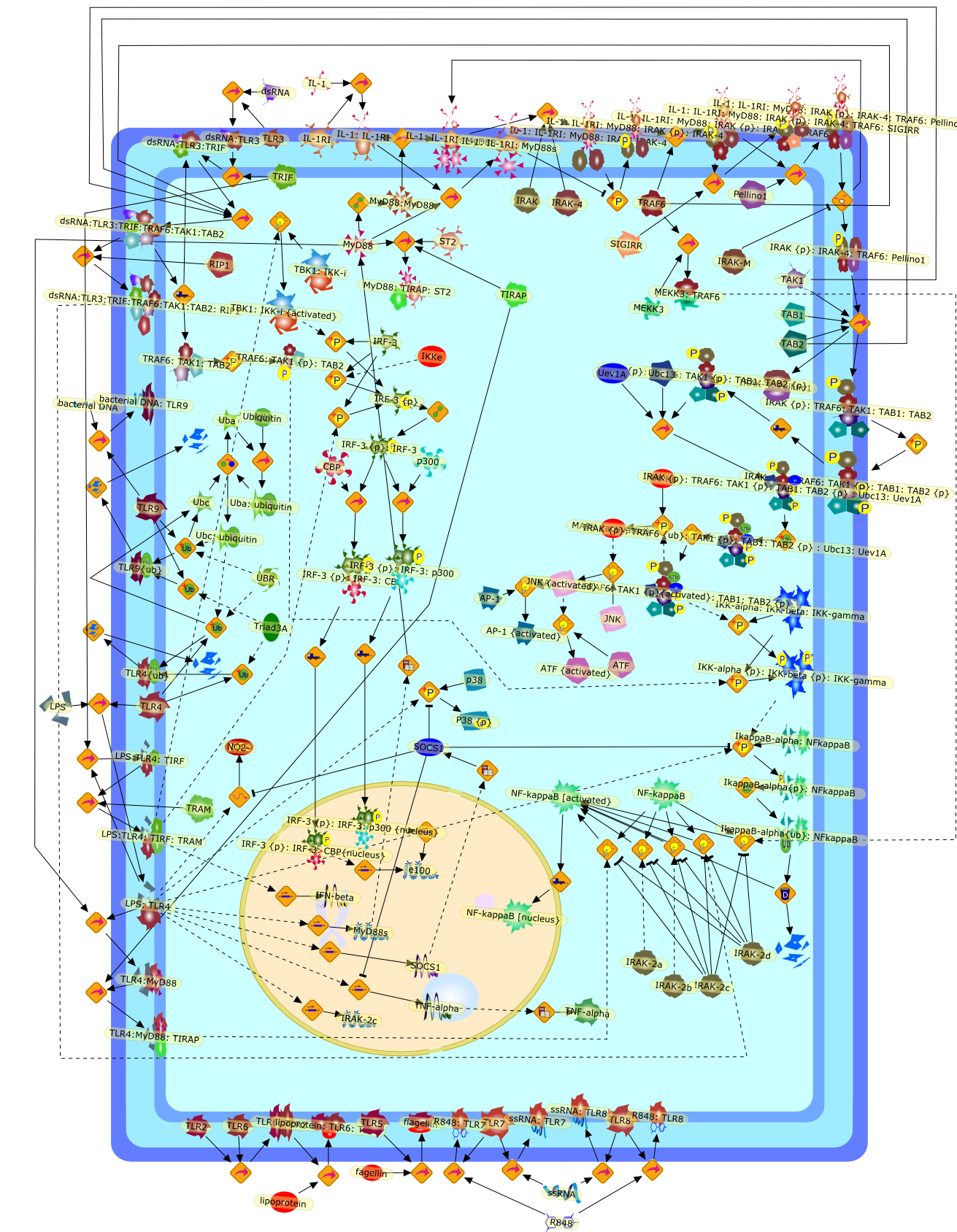{kind=link}
{kind=link}
AP-1,AP-1 {activated},ATF,ATF {activated},CBP,IFN-beta,IKK-alpha {p}: IKK-beta {p}: IKK-gamma,IKK-alpha: IKK-beta: IKK-gamma,IKKe,IL-1,IL-1: IL-1RI,IL-1: IL-1RI: MyD88,IL-1: IL-1RI: MyD88: IRAK [p}: IRAK-4: TRAF6: Pellino1,IL-1: IL-1RI: MyD88: IRAK {p}: IRAK-4,IL-1: IL-1RI: MyD88: IRAK {p}: IRAK-4: TRAF6,IL-1: IL-1RI: MyD88: IRAK {p}: IRAK-4: TRAF6: SIGIRR,IL-1: IL-1RI: MyD88: IRAK: IRAK-4,IL-1: IL-1RI: MyD88s,IL-1RI,IRAK,IRAK {p}: IRAK-4: TRAF6: Pellino1,IRAK {p}: TRAF6 {ub}: TAK1 {p}: TAB1: TAB2 {p} : Ubc13: Uev1A,IRAK {p}: TRAF6: TAK1 {p}: TAB1: TAB2 {p},IRAK {p}: TRAF6: TAK1 {p}: TAB1: TAB2 {p} ,IRAK {p}: TRAF6: TAK1 {p}: TAB1: TAB2 {p} : Ubc13: Uev1A,IRAK {p}: TRAF6: TAK1 {p}{activated}: TAB1: TAB2 {p} ,IRAK {p}: TRAF6: TAK1: TAB1: TAB2,IRAK-2a,IRAK-2b,IRAK-2c,IRAK-2d,IRAK-4,IRAK-4: Pellino1,IRAK-M,IRF-3,IRF-3 {p},IRF-3 {p}: IRF-3,IRF-3 {p}: IRF-3: CBP,IRF-3 {p}: IRF-3: CBP{nucleus},IRF-3 {p}: IRF-3: p300,IRF-3 {p}: IRF-3: p300 {nucleus},IkappaB-alpha {degraded},IkappaB-alpha: NFkappaB,IkappaB-alpha{p}: NFkappaB,IkappaB-alpha{ub}: NFkappaB,JNK,JNK {activated},LPS,LPS: TLR4,LPS:TLR4: TIRF,LPS:TLR4: TIRF: TRAM,MAPK {activated},MAPK6,MEKK3,MEKK3: TRAF6,MyD88,MyD88: TIRAP: ST2,MyD88:MyD88,MyD88s,NF-kappaB,NF-kappaB [activated},NF-kappaB [nucleus},NO2-,P38 {p},Pellino1,R848,R848: TLR7,R848: TLR8,RIP1,SIGIRR,SOCS1,ST2,TAB1,TAB2,TAK1,TBK1: IKK-i,TBK1: IKK-i {activated},TIRAP,TLR2,TLR3,TLR4,TLR4 {degraded},TLR4:MyD88,TLR4:MyD88: TIRAP,TLR4{ub},TLR5,TLR6,TLR6: TLR2,TLR7,TLR8,TLR9,TLR9 {degraded},TLR9{ub},TNF-alpha,TRAF6,TRAF6: TAK1 {p}: TAB2,TRAF6: TAK1: TAB2,TRAM,TRIF,Triad3A,UBR,Uba,Uba: ubiquitin,Ubc,Ubc13,Ubc: ubiquitin,Ubiquitin,Uev1A,bacterial DNA,bacterial DNA: TLR9,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:TRAF6:TAK1:TAB2,dsRNA:TLR3:TRIF:TRAF6:TAK1:TAB2: RIP1,e100,fagellin,flagellin,lipoprotein,lipoprotein: TLR6: TLR2,p300,p38,ssRNA,ssRNA: TLR7,ssRNA: TLR8
Macrophages participate actively in the inflammatory response by releasingcytokines, chemokines and factors that recruit additional cells to sites ofinfection or tissue injury or alteration. In addition to this, activatedmacrophages rapidly activate the expression of genes responsible for thehigh-output synthesis of reactive oxygen and nitrogen species (NO, O2-, H2O2 andperoxynitrite, among others) and bioactive lipids derived from arachidonic acid.All of these agents contribute to the regulation of the inflammatory response.Most of these molecules, when synthesized at these high concentrations, exertpro-apoptotic effects in many cell types. Macrophages themselves are a notableand important exception, being resistant to apoptotic death upon activation.This resistance is necessary to enable these cells to perform their functionalrole during the early phases of an inflammatory response. However, aftercumulative damage, or when the synthesis of inflammatory mediators decreases,macrophages undergo the characteristic mitochondrial-dependent cell deathprogram, contributing in this way to the resolution of the inflammatoryreaction. In the case of infectious diseases, this also helps to prevent thedevelopment of parasitic strategies by phagocytosed pathogens.
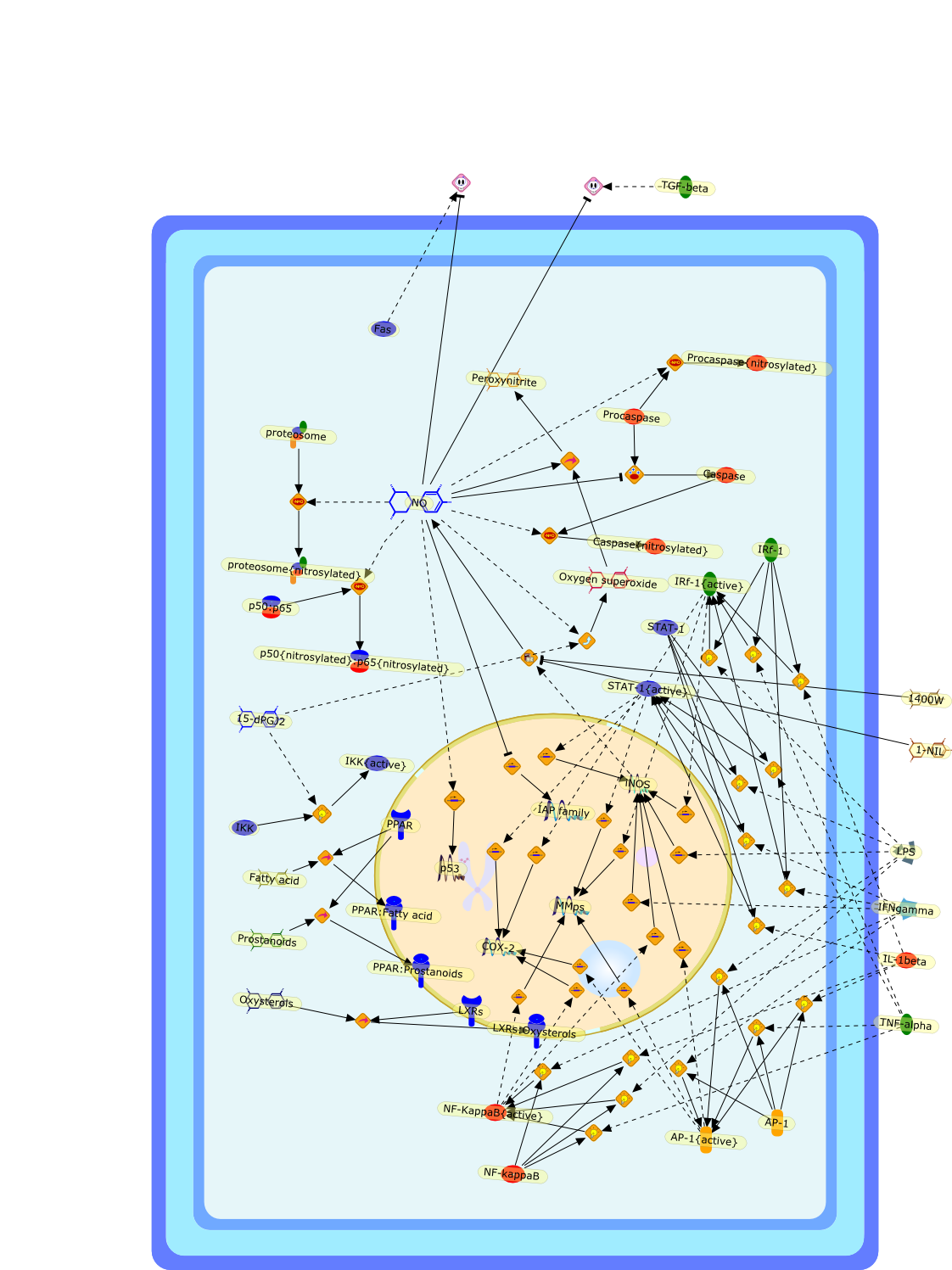{kind=link}
{kind=link}
1-NIL,1400W,15-dPGJ2,AP-1,AP-1{active},COX-2,Caspase,Caspase{nitrosylated},Fas,Fatty acid,IAP family,IFNgamma,IKK,IKK{active},IL-1beta,IRf-1,IRf-1{active},LPS,LXRs,LXRs:Oxysterols,MMps,NF-KappaB{active},NF-kappaB,NO,Oxygen superoxide,Oxysterols,PPAR,PPAR:Fatty acid,PPAR:Prostanoids,Peroxynitrite,Procaspase,Procaspase{nitrosylated},Prostanoids,STAT-1,STAT-1{active},TGF-beta,TNF-alpha,iNOS,p50:p65,p50{nitrosylated}:p65{nitrosylated},p53,proteosome,proteosome{nitrosylated}
The human pim-1 gene encodes a serine/threonine kinase, which belongs to thegroup of calcium/calmodulin-regulated kinases (CAMK). It contains acharacteristic kinase domain, a so-called ATP anchor and an active site. Inmouse and human, two Pim-1 proteins are produced from the same gene by using analternative upstream CUG initiation codon, a 44 kD and another, shorter 34 kDform that both contain the kinase domain. Expression of Pim-1 is widespread andranges from the hematopoietic and lymphoid system to prostate, testis and oralepithelial cells. Two other proteins with significant sequence similaritiesexist, Pim-2 and Pim-3; both are also serine/threonine kinases and have largelyoverlapping functions. Pim-1 is able to phosphorylate different targets, most ofwhich are involved in cell cycle progression or apoptosis. Pim-1 expression canbe induced by several external stimuli in particular by a number of cytokinesrelevant in the immune system, which led to the labeling of Pim-1 as a "booster"for the immune response.
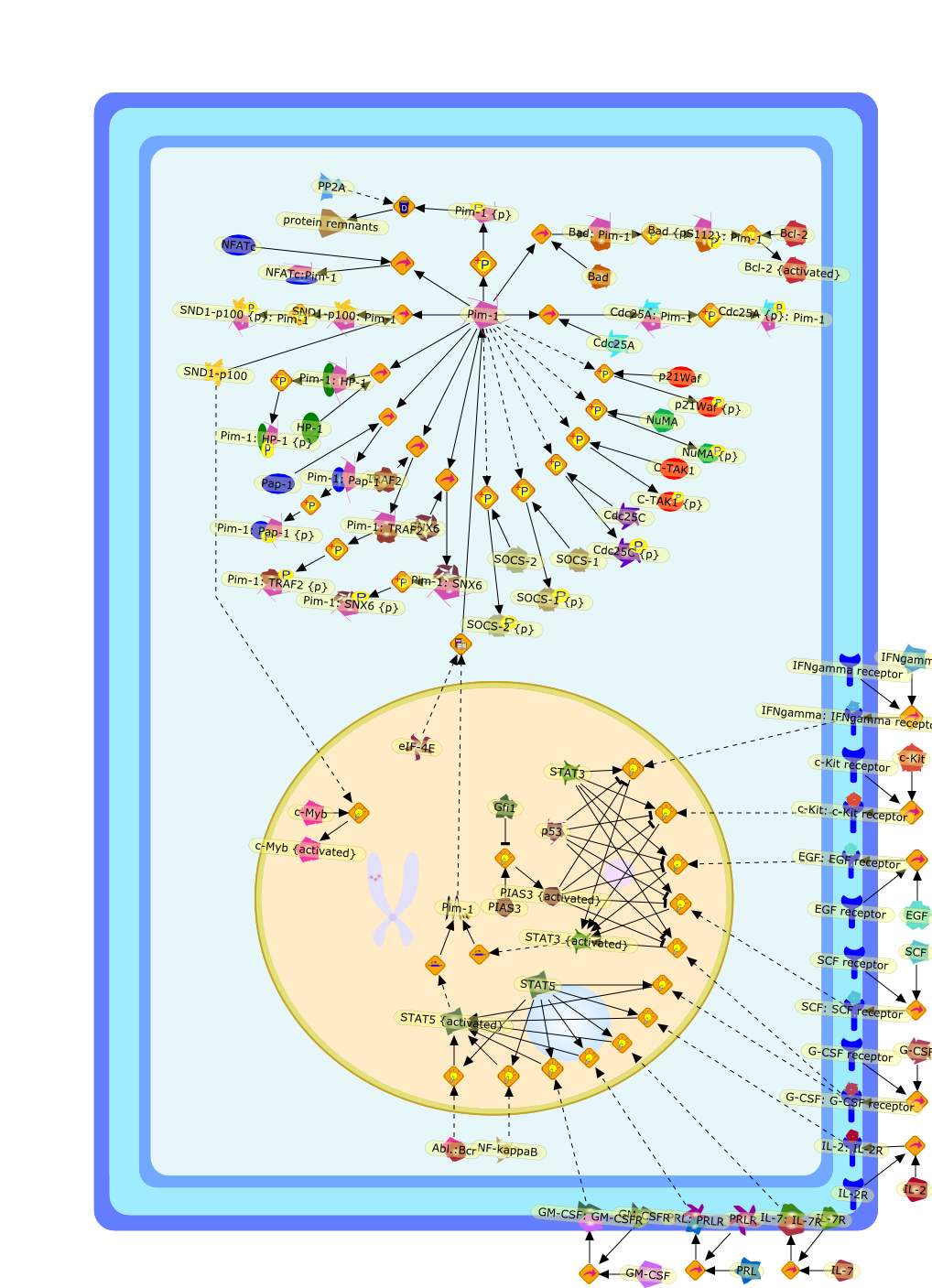{kind=link}
{kind=link}
Abl.:Bcr,Bad,Bad {pS112}: Pim-1,Bad: Pim-1,Bcl-2,Bcl-2 {activated},C-TAK1,C-TAK1 {p},Cdc25A,Cdc25A {p}: Pim-1,Cdc25A: Pim-1,Cdc25C,Cdc25C {p},EGF,EGF receptor,EGF: EGF receptor,G-CSF,G-CSF receptor,G-CSF: G-CSF receptor,GM-CSF,GM-CSF: GM-CSFR,GM-CSFR,Gfi1,HP-1,IFNgamma,IFNgamma receptor,IFNgamma: IFNgamma receptor,IL-2,IL-2: IL-2R,IL-2R,IL-7,IL-7: IL-7R,IL-7R,NF-kappaB,NFATc,NFATc:Pim-1,NuMA,NuMA {p},PIAS3,PIAS3 {activated},PP2A,PRL,PRL: PRLR,PRLR,Pap-1,Pim-1,Pim-1 {p},Pim-1: HP-1,Pim-1: HP-1 {p},Pim-1: Pap-1,Pim-1: Pap-1 {p},Pim-1: SNX6,Pim-1: SNX6 {p},Pim-1: TRAF2,Pim-1: TRAF2 {p},SCF,SCF receptor,SCF: SCF receptor,SND1-p100,SND1-p100 {p}: Pim-1,SND1-p100: Pim-1,SNX6,SOCS-1,SOCS-1 {p},SOCS-2,SOCS-2 {p},STAT3,STAT3 {activated},STAT5,STAT5 {activated},TRAF2,c-Kit,c-Kit receptor,c-Kit: c-Kit receptor,c-Myb,c-Myb {activated},eIF-4E,p21Waf,p21Waf {p},p53,protein remnants
Mammalian Toll-like receptors recognize components of invading microbes andtrigger the first line of innate immune response that is mediated bytranscriptional induction of a large number of cellular genes. Toll-likereceptor 3 (TLR3) is thought to be a major mediator of cellular response toviral infection, because it responds to double-stranded (ds) RNA, a commonby-product of viral replication. This article is focused on the nature of thesignaling pathways activated by TLR3 and dsRNA. The genes induced by TLR3activation include those that encode secreted antiviral cytokines, such asinterferon (IFN), and those that encode intracellular viral stress-inducibleproteins. Recent studies have revealed several unique features of TLR3 signalingthat are highlighted here. Specifically, we discuss the roles of receptortyrosine phosphorylation, PI3 kinase and two-step activation of thetranscription factors, IRF-3 and NF-kappaB, in mediating TLR3-signaling.
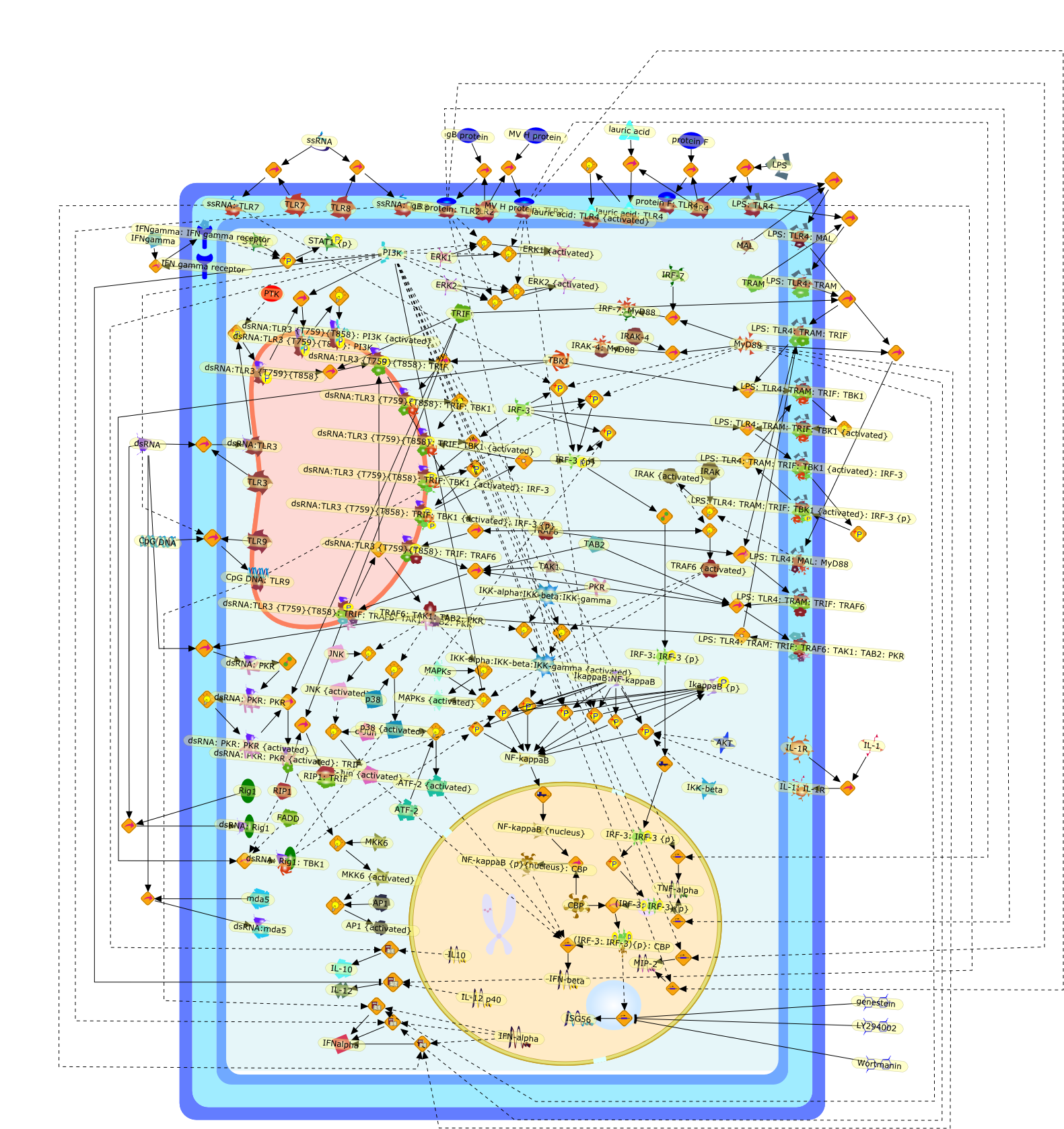{kind=link}
{kind=link}
(IRF-3: IRF-3){p},(IRF-3: IRF-3){p}: CBP,AKT,AP1,AP1 {activated},ATF-2,ATF-2 {activated},CBP,CpG DNA,CpG DNA: TLR9,ERK1,ERK1 {activated},ERK2,ERK2 {activated},FADD,IFN gamma receptor,IFN-alpha,IFN-beta,IFNalpha,IFNgamma,IFNgamma: IFN gamma receptor,IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma {activated},IKK-beta,IL-1,IL-10,IL-12,IL-12 p40,IL-1: IL-1R,IL-1R,IL10,IRAK,IRAK {activated},IRAK-4,IRAK-4: MyD88,IRF-3,IRF-3 {p},IRF-3: IRF-3 {p},IRF-7,IRF-7: MyD88,ISG56,IkappaB {p},IkappaB:NF-kappaB,JNK,JNK {activated},LPS,LPS: TLR4,LPS: TLR4: MAL,LPS: TLR4: MAL: MyD88,LPS: TLR4: TRAM,LPS: TLR4: TRAM: TRIF,LPS: TLR4: TRAM: TRIF: TBK1,LPS: TLR4: TRAM: TRIF: TBK1 {activated},LPS: TLR4: TRAM: TRIF: TBK1 {activated}: IRF-3,LPS: TLR4: TRAM: TRIF: TBK1 {activated}: IRF-3 {p},LPS: TLR4: TRAM: TRIF: TRAF6,LPS: TLR4: TRAM: TRIF: TRAF6: TAK1: TAB2: PKR,LY294002,MAL,MAPKs,MAPKs {activated},MIP-2,MKK6,MKK6 {activated},MV H protein,MV H protein: TLR2,MyD88,NF-kappaB,NF-kappaB {nucleus},NF-kappaB {p}{nucleus}: CBP,PI3K,PKR,PTK,RIP1,RIP1: TRIF,Rig1,STAT1,STAT1 {p},TAB2,TAK1,TBK1,TLR2,TLR3,TLR4,TLR7,TLR8,TLR9,TNF-alpha,TRAF6,TRAF6 {activated},TRAF6: TAK1: TAB2: PKR,TRAM,TRIF,Wortmanin,c-Jun,c-Jun {activated},dsRNA,dsRNA: PKR,dsRNA: PKR: PKR,dsRNA: PKR: PKR {activated},dsRNA: PKR: PKR {activated}: TRIF,dsRNA: Rig1,dsRNA: Rig1: TBK1,dsRNA:TLR3,dsRNA:TLR3 {T759}{T858},dsRNA:TLR3 {T759}{T858}: PI3K,dsRNA:TLR3 {T759}{T858}: PI3K {activated},dsRNA:TLR3 {T759}{T858}: TRIF,dsRNA:TLR3 {T759}{T858}: TRIF: TBK1,dsRNA:TLR3 {T759}{T858}: TRIF: TBK1 {activated},dsRNA:TLR3 {T759}{T858}: TRIF: TBK1 {activated}: IRF-3,dsRNA:TLR3 {T759}{T858}: TRIF: TBK1 {activated}: IRF-3 {p},dsRNA:TLR3 {T759}{T858}: TRIF: TRAF6,dsRNA:TLR3 {T759}{T858}: TRIF: TRAF6: TAK1: TAB2: PKR,dsRNA:mda5,gB protein,gB protein: TLR2,genestein,lauric acid,lauric acid: TLR4,lauric acid: TLR4 {activated},mda5,p38,p38 {activated},protein F,protein F: TLR4,ssRNA,ssRNA: TLR7,ssRNA: TLR8
{kind=link}
{kind=link}
AP-1,AP-1{active},ASC,Bacteria,CARD8,CIAS1,COX-2,COX2,CREB,CREB{active},Caspase-1,Cryopyrin,Cryopyrin:ASC:CARD8:caspase-1,Cryopyrin:MDP,E-selectin,ERK,ERK{active},ICAM,IKK complex(IKKalpha:IKKbeta:IKKgamma),IKappaB-alpha:NF-KappaB,IKappaB-alpha{p}:NF-KappaB,IKk complex(IKKalpha:IKKbeta:IKKgamma){active},IKk complex(IKKalpha:IKKbeta:IKKgamma{p}){active},IL-18,IL-1beta,IL-6,IL-8,JNK,JNK{active},Lipoproteins,Lipoteichoic acid,MAPK,MAPK{active},MCP-1,MCP1,MDP,MIP-1alpha,NF-kappaB,NF-kappaB{active},Nod1,Nod1 oligomer:iE-DAP,Nod1 oligomer:iE-DAP:RIP2,Nod1:iE-DAP,Nod2,Nod2 oligomer:PGN (MDP),Nod2 oligomer:PGN(MDP):RIP2,Nod2:MDP,PGN,PGRP-L,PGRP-L:PGN,PGRPs,PGRPs:Bacteria,PGRPs:PGN,RIP2,Remnants,SRF,SRF{active},TLR2,TLR2:Lipoproteins,TLR2:Lipoteichoic acid,TNF-alpha,VCAM,cryopyrin,iE-DAP,iNOS,p38,p38{active},proIL-18,proIL-1beta,protein remnants
The nuclear factor (NF)-kappaB pathway is a paradigm for gene expression controlby ubiquitin-mediated protein degradation. In stimulated cells, phosphorylationby the IkappaB kinase (IKK) complex primes NF-kappaB-inhibiting IkappaBmolecules for lysine (Lys)-48-linked polyubiquitination and subsequentdestruction by the 26S proteasome. However, recent studies indicate that theubiquitin (Ub) system controls NF-kappaB pathways at many levels. Ub ligases areactivated by different upstream signalling pathways, and they function ascentral regulators of IKK and c-Jun amino-terminal kinase activation. Theassembly of Lys 63 polyUb chains provides docking surfaces for the recruitmentof IKK-activating complexes, a reaction that is counteracted by deubiquitinatingenzymes. Furthermore, Ub conjugation targets upstream signalling mediators aswell as nuclear NF-kappaB for post-inductive degradation to limit the durationof signalling.
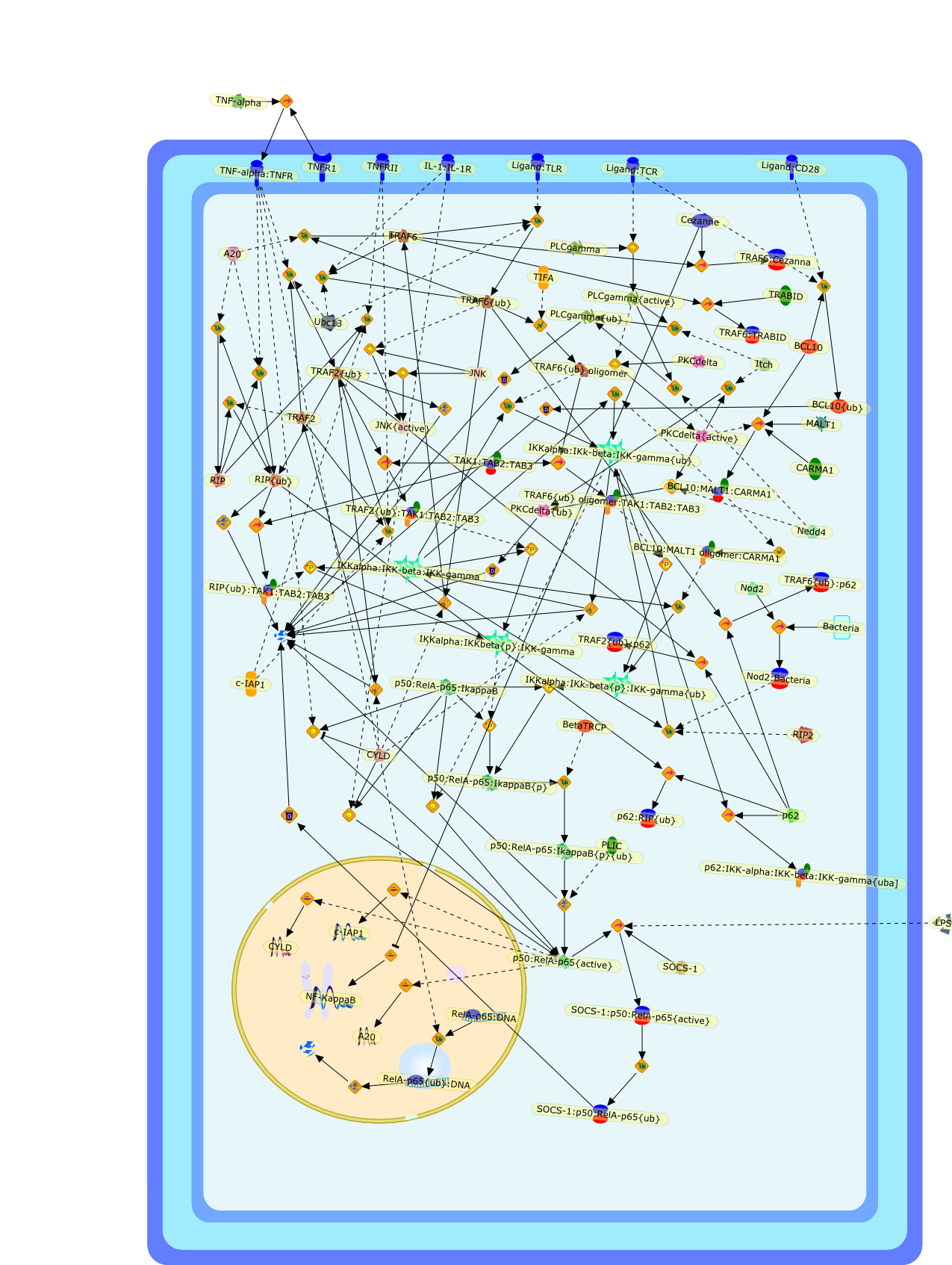{kind=link}
{kind=link}
A20,BCL10,BCL10:MALT1 oligomer:CARMA1,BCL10:MALT1:CARMA1,BCL10{ub},Bacteria,BetaTRCP,CARMA1,CYLD,Cezanne,IKKalpha:IKK-beta:IKK-gamma,IKKalpha:IKKbeta{p}:IKK-gamma,IKKalpha:IKk-beta:IKK-gamma{ub},IKKalpha:IKk-beta{p}:IKK-gamma{ub},IL-1:IL-1R,Itch,JNK,JNK{active},LPS,Ligand:CD28,Ligand:TCR,Ligand:TLR,MALT1,NF-KappaB,Nedd4,Nod2,Nod2:Bacteria,PKCdelta,PKCdelta{active},PKCdelta{ub},PLCgamma,PLCgamma{active},PLCgamma{ub},PLIC,RIP,RIP2,RIP{ub},RIP{ub}:TAK1:TAB2:TAB3,RelA-p65:DNA,RelA-p65{ub}:DNA,SOCS-1,SOCS-1:p50:RelA-p65{active},SOCS-1:p50:RelA-p65{ub},TAK1:TAB2:TAB3,TIFA,TNF-alpha,TNF-alpha:TNFR,TNFR1,TNFRII,TRABID,TRAF2,TRAF2{ub},TRAF2{ub}:TAK1:TAB2:TAB3,TRAF2{ub}:p62,TRAF6,TRAF6:Cezanna,TRAF6:TRABID,TRAF6{ub},TRAF6{ub} oligomer,TRAF6{ub} oligomer:TAK1:TAB2:TAB3,TRAF6{ub}:p62,Ubc13,c-IAP1,p50:RelA-p65:IkappaB,p50:RelA-p65:IkappaB{p},p50:RelA-p65:IkappaB{p}{ub},p50:RelA-p65{active},p62,p62:IKK-alpha:IKK-beta:IKK-gamma{uba],p62:RIP{ub},protein degradants,protein remnants
It is now well established that Toll-like receptors (TLRs) act as primarysensors of microbial compounds. Details of the molecular mechanisms governingTLR responses are emerging steadily and our understanding of the signalingpathways activated these receptors has improved greatly over the last few years.Differences in adaptor usage, cellular localisation and signaling cascades havebeen elucidated. In this review we will summarize the current understanding ofTLR signaling and its regulation.
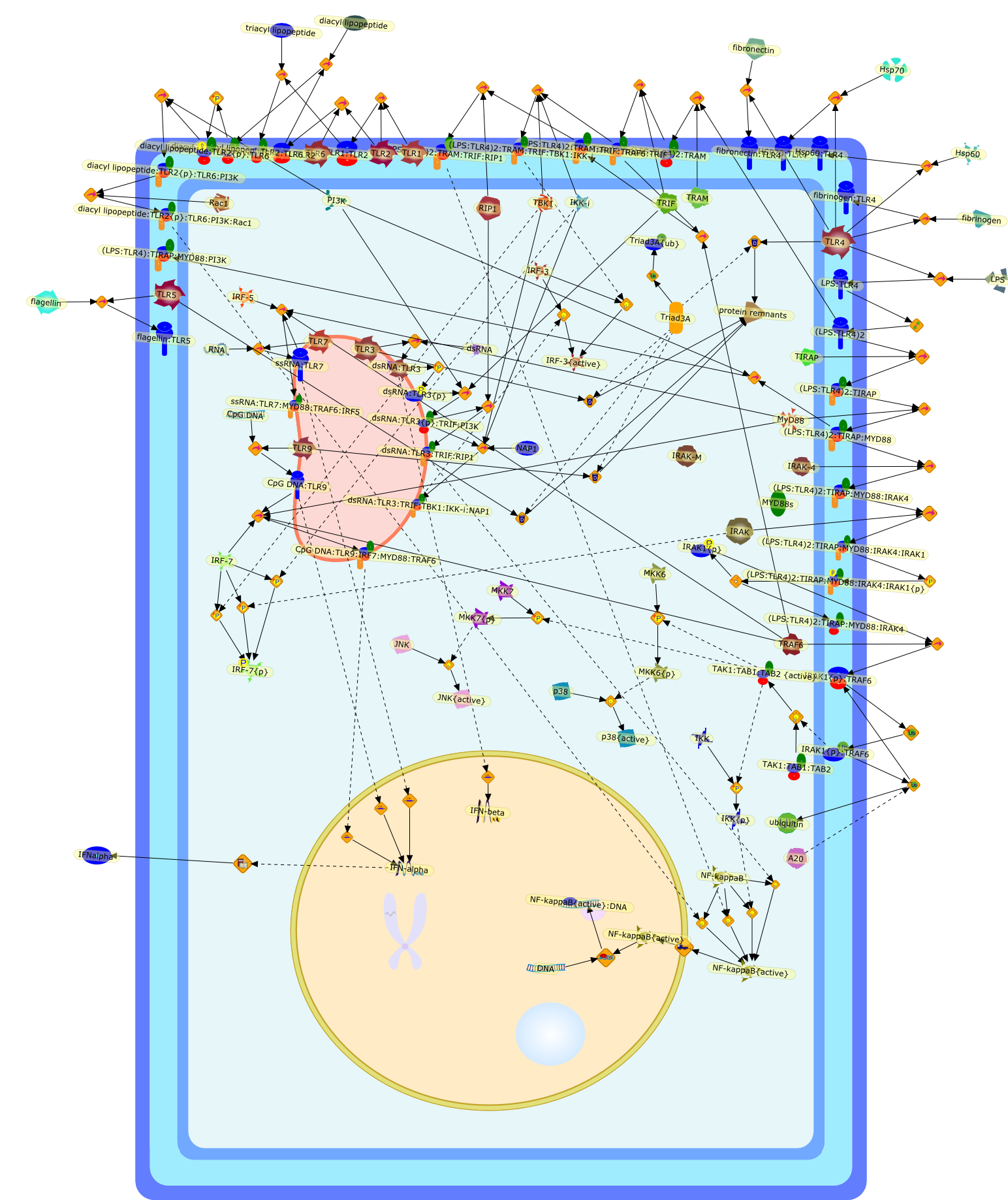{kind=link}
{kind=link}
(LPS:TLR4)2,(LPS:TLR4)2:TIRAP,(LPS:TLR4)2:TIRAP:MYD88,(LPS:TLR4)2:TIRAP:MYD88:IRAK4,(LPS:TLR4)2:TIRAP:MYD88:IRAK4:IRAK1,(LPS:TLR4)2:TIRAP:MYD88:IRAK4:IRAK1{p},(LPS:TLR4)2:TRAM,(LPS:TLR4)2:TRAM:TRIF,(LPS:TLR4)2:TRAM:TRIF:RIP1,(LPS:TLR4)2:TRAM:TRIF:TBK1:IKK-i,(LPS:TLR4)2:TRAM:TRIF:TRAF6,(LPS:TLR4):TIRAP:MYD88:PI3K,A20,CpG DNA,CpG DNA:TLR9,CpG DNA:TLR9:IRF7:MYD88:TRAF6,DNA,HSP70:TLR4,Hsp60,Hsp60:TLR4,Hsp70,IFN-alpha,IFN-beta,IFNalpha,IKK,IKK-i,IKK{p},IRAK,IRAK-4,IRAK-M,IRAK1{p},IRAK1{p}:TRAF6,IRF-3,IRF-3{active},IRF-5,IRF-7,IRF-7{p},JNK,JNK{active},LPS,LPS:TLR4,MKK6,MKK6{p},MKK7,MKK7{p},MYD88s,MyD88,NAP1,NF-kappaB,NF-kappaB{active},NF-kappaB{active}:DNA,PI3K,RIP1,RNA,Rac1,TAK1:TAB1:TAB2,TAK1:TAB1:TAB2 {active},TBK1,TIRAP,TLR1,TLR1:TLR2,TLR2,TLR2:TLR6,TLR3,TLR4,TLR5,TLR6,TLR7,TLR9,TRAF6,TRAM,TRIF,Triad3A,Triad3A{ub},diacyl lipopeptide,diacyl lipopeptide:TLR2:TLR6,diacyl lipopeptide:TLR2{p}:TLR6,diacyl lipopeptide:TLR2{p}:TLR6:PI3K,diacyl lipopeptide:TLR2{p}:TLR6:PI3K:Rac1,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF:RIP1,dsRNA:TLR3:TRIF:TBK1:IKK-i:NAP1,dsRNA:TLR3{p},dsRNA:TLR3{p}:TRIF:PI3K,fibrinogen,fibrinogen:TLR4,fibronectin,fibronectin:TLR4,flagellin,flagellin:TLR5,p38,p38{active},protein remnants,ssRNA:TLR7,ssRNA:TLR7:MYD88:TRAF6:IRF5,triacyl lipopeptide,triacyl lipopeptide:TLR1:TLR2,ubiquitin
Cross-presentation is important for regulating T-cell responses to exogenousantigens and can maintain tolerance (cross-tolerance) or induce immune responses(cross-priming). Recent exciting results on the role of the Toll-like receptorTLR3 in promoting cross-priming of viral antigens provide new insights into themechanisms that allow Toll-like receptor signaling to bridge innate and adaptiveimmune responses.
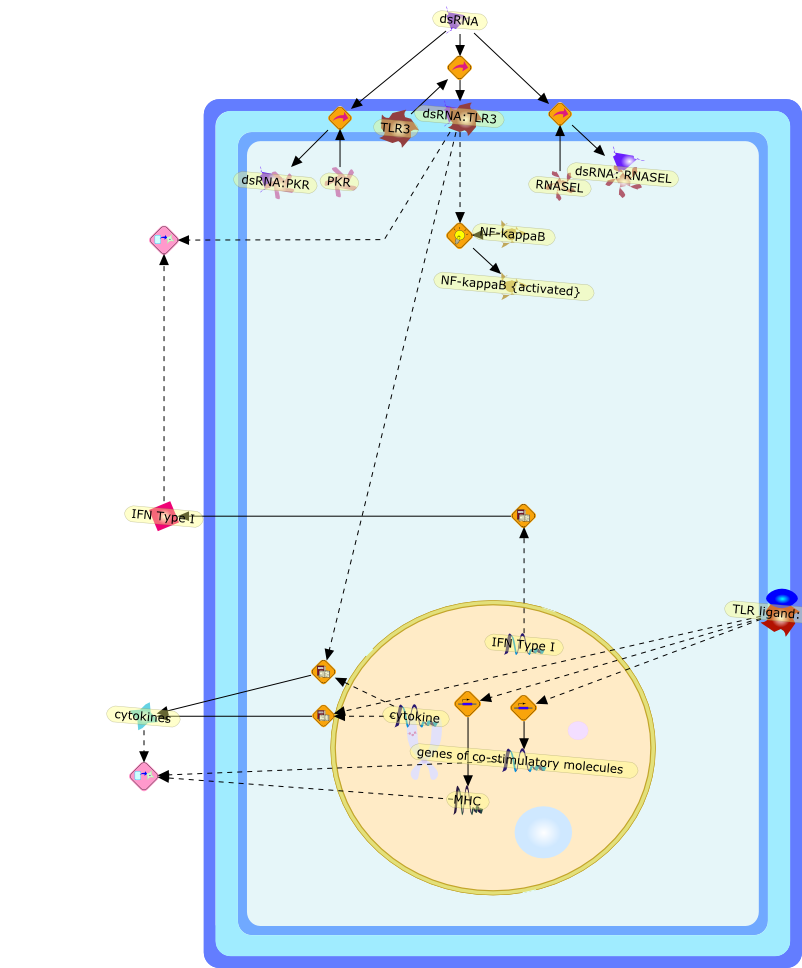{kind=link}
{kind=link}
IFN Type I,MHC,NF-kappaB,NF-kappaB {activated},PKR,RNASEL,TLR ligand: TLR,TLR3,cytokine,cytokines,dsRNA,dsRNA: RNASEL,dsRNA:PKR,dsRNA:TLR3,genes of co-stimulatory molecules
Toll-like receptors (TLRs) and their ligands have emerged as importantregulators of immunity, relevant to a wide range of effector responses fromvaccination to autoimmunity. The most well-studied ligands of TLRs expressed onB cells include the lipopolysaccharides (for TLR4) and CpG-containing DNAs (forTLR9), which induce and/or co-stimulate B cells to undergo proliferation, classswitching and differentiation into antibody-secreting cells. Recent developmentsin this area include advancements into our understanding of the role of thesereceptor pathways in B cells, and in particular the relevance of TLR9, which hasreceived substantial attention as both a Th1-like inflammatory immunomodulatorand a pathogenic co-stimulator of autoreactive B cell responses.
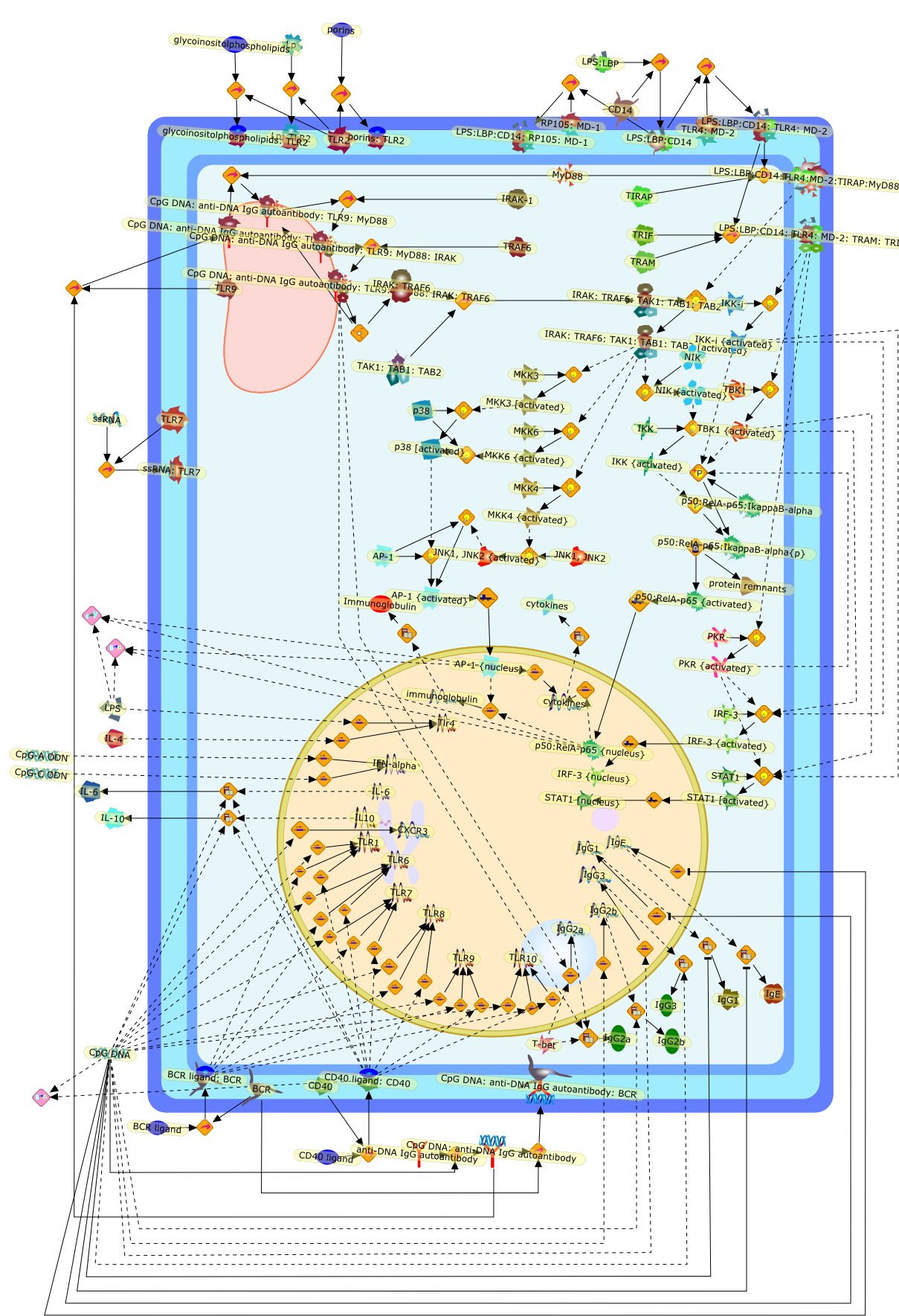{kind=link}
{kind=link}
AP-1,AP-1 {activated},AP-1 {nucleus},BCR,BCR ligand,BCR ligand: BCR,CD14,CD40,CD40 ligand,CD40 ligand: CD40,CXCR3,CpG DNA,CpG DNA: anti-DNA IgG autoantibody,CpG DNA: anti-DNA IgG autoantibody: BCR,CpG DNA: anti-DNA IgG autoantibody: TLR9,CpG DNA: anti-DNA IgG autoantibody: TLR9: MyD88,CpG DNA: anti-DNA IgG autoantibody: TLR9: MyD88: IRAK,CpG DNA: anti-DNA IgG autoantibody: TLR9: MyD88: IRAK: TRAF6,CpG-A ODN,CpG-C ODN,IFN-alpha,IKK,IKK {activated},IKK-i,IKK-i {activated},IL-10,IL-4,IL-6,IL10,IRAK-1,IRAK: TRAF6,IRAK: TRAF6: TAK1: TAB1: TAB2,IRAK: TRAF6: TAK1: TAB1: TAB2 [activated},IRF-3,IRF-3 {activated},IRF-3 {nucleus},IgE,IgG1,IgG2a,IgG2b,IgG3,Immunoglobulin,JNK1, JNK2,JNK1, JNK2 {activated},LPS,LPS:LBP,LPS:LBP:CD14,LPS:LBP:CD14: RP105: MD-1,LPS:LBP:CD14: TLR4: MD-2,LPS:LBP:CD14: TLR4: MD-2: TRAM: TRIF,LPS:LBP:CD14:TLR4:MD-2:TIRAP:MyD88,Lp,Lp: TLR2,MKK3,MKK3 [activated},MKK4,MKK4 {activated},MKK6,MKK6 {activated},MyD88,NIK,NIK {activated},PKR,PKR {activated},RP105: MD-1,STAT1,STAT1 [activated},STAT1 [nucleus},T-bet,TAK1: TAB1: TAB2,TBK1,TBK1 {activated},TIRAP,TLR1,TLR10,TLR2,TLR4: MD-2,TLR6,TLR7,TLR8,TLR9,TRAF6,TRAM,TRIF,Tlr4,anti-DNA IgG autoantibody,cytokines,glycoinositolphospholipids,glycoinositolphospholipids: TLR2,immunoglobulin,p38,p38 [activated},p50:RelA-p65 {activated},p50:RelA-p65 {nucleus},p50:RelA-p65:IkappaB-alpha,p50:RelA-p65:IkappaB-alpha{p},porins,porins: TLR2,protein remnants,ssRNA,ssRNA: TLR7
The innate immune system is an evolutionarily conserved system of defenseagainst microbial infections. The family of Toll-like receptors is a major classof receptors that sense molecular patterns associated with a broad range ofpathogens including bacteria, viruses, fungi and protozoa. Following pathogenrecognition, Toll-like receptors initiate intracellular signal transduction thatresults in the expression of genes involved in inflammation, antiviral responsesand maturation of dendritic cells. Individual Toll-like receptors activatecommon and unique transcription factors through different signaling pathways todrive specific biological responses against microorganisms.
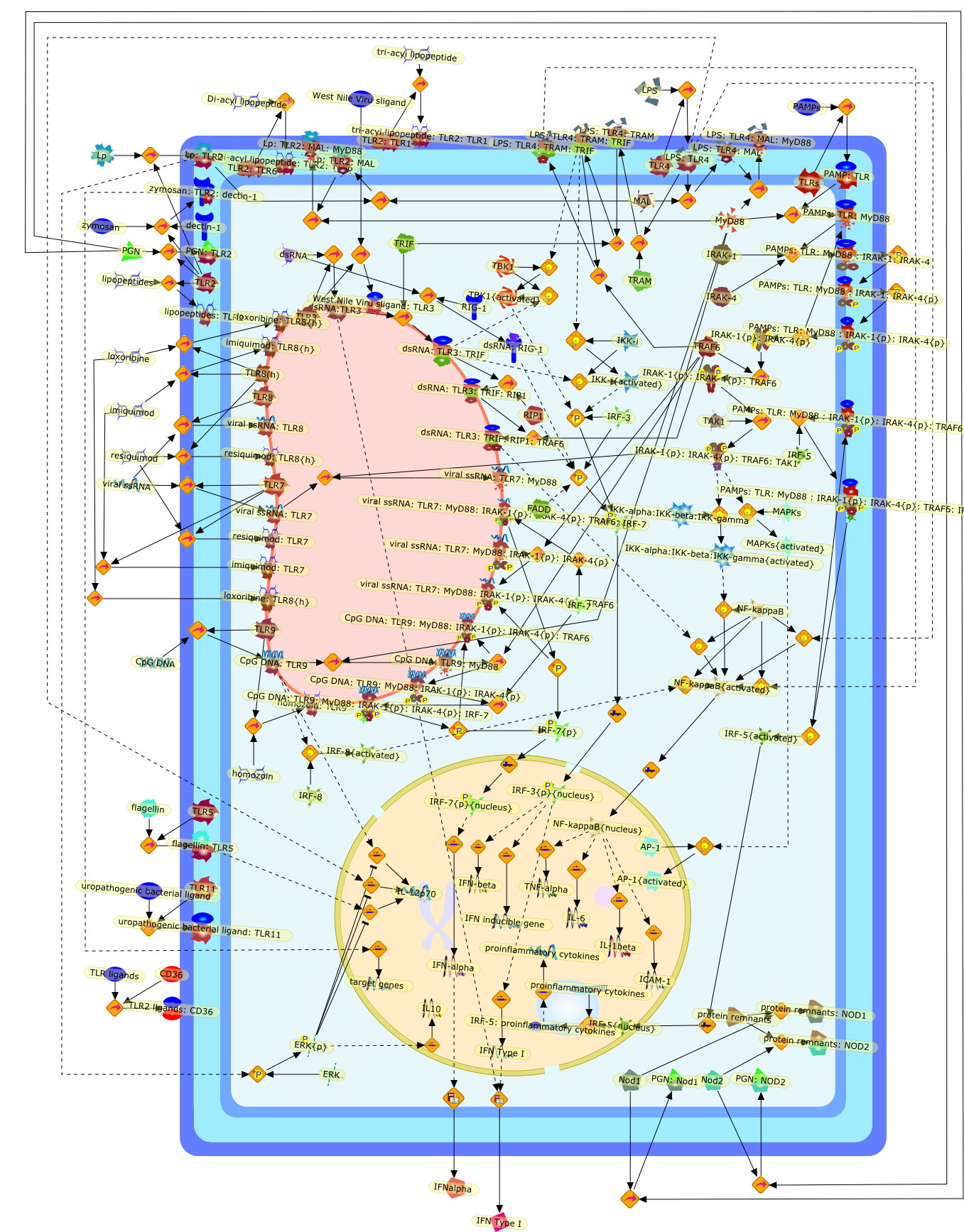{kind=link}
{kind=link}
AP-1,AP-1{activated},CD36,CpG DNA,CpG DNA: TLR9,CpG DNA: TLR9: MyD88,CpG DNA: TLR9: MyD88: IRAK-1{p}: IRAK-4{p},CpG DNA: TLR9: MyD88: IRAK-1{p}: IRAK-4{p}: IRF-7,CpG DNA: TLR9: MyD88: IRAK-1{p}: IRAK-4{p}: TRAF6,Di-acyl lipopeptide,Di-acyl lipopeptide: TLR2: TLR6,ERK,ERK{p},FADD,ICAM-1,IFN Type I,IFN inducible gene,IFN-alpha,IFN-beta,IFNalpha,IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma{activated},IKK-i,IKK-i{activated},IL-12p70,IL-1beta,IL-6,IL10,IRAK-1,IRAK-1{p}: IRAK-4{p},IRAK-1{p}: IRAK-4{p}: TRAF6,IRAK-1{p}: IRAK-4{p}: TRAF6: TAK1,IRAK-4,IRF-3,IRF-3{p},IRF-3{p}{nucleus},IRF-5,IRF-5: proinflammatory cytokines,IRF-5{activated},IRF-5{nucleus},IRF-7,IRF-7{p},IRF-7{p}{nucleus},IRF-8,IRF-8{activated},LPS,LPS: TLR4,LPS: TLR4: MAL,LPS: TLR4: MAL: MyD88,LPS: TLR4: TRAM,LPS: TLR4: TRAM: TRIF,Lp,Lp: TLR2,Lp: TLR2: MAL,Lp: TLR2: MAL: MyD88,MAL,MAPKs,MAPKs{activated},MyD88,NF-kappaB,NF-kappaB{activated},NF-kappaB{nucleus},Nod1,Nod2,PAMP: TLR,PAMPs,PAMPs: TLR: MyD88,PAMPs: TLR: MyD88 : IRAK-1: IRAK-4,PAMPs: TLR: MyD88 : IRAK-1: IRAK-4{p},PAMPs: TLR: MyD88 : IRAK-1{p}: IRAK-4{p},PAMPs: TLR: MyD88 : IRAK-1{p}: IRAK-4{p}: TRAF6,PAMPs: TLR: MyD88 : IRAK-1{p}: IRAK-4{p}: TRAF6: IRF-5,PGN,PGN: NOD2,PGN: Nod1,PGN: TLR2,RIG-1,RIP1,TAK1,TBK1,TBK1{activated},TLR ligands,TLR11,TLR2,TLR2 ligands: CD36,TLR2: TLR1,TLR2: TLR6,TLR3,TLR4,TLR5,TLR7,TLR8,TLR8(h),TLR9,TLRs,TNF-alpha,TRAF6,TRAM,TRIF,West Nile Viru sligand,West Nile Viru sligand: TLR3,dectin-1,dsRNA,dsRNA: RIG-1,dsRNA: TLR3: TRIF,dsRNA: TLR3: TRIF: RIP1,dsRNA: TLR3: TRIF: RIP1: TRAF6,dsRNA:TLR3,flagellin,flagellin: TLR5,homozoin,homozoin: TLR9,imiquimod,imiquimod: TLR7,imiquimod: TLR8{h},lipopeptides,lipopeptides: TLR2,loxoribine,loxoribine: TLR8{h},proinflammatory cytokines,protein remnants,protein remnants: NOD1,protein remnants: NOD2,resiquimod,resiquimod: TLR7,resiquimod: TLR8{h},target genes,tri-acyi lipopeptide,tri-acyi lipopeptide: TLR2: TLR1,uropathogenic bacterial ligand,uropathogenic bacterial ligand: TLR11,viral ssRNA,viral ssRNA: TLR7,viral ssRNA: TLR7: MyD88,viral ssRNA: TLR7: MyD88: IRAK-1{p}: IRAK-4{p},viral ssRNA: TLR7: MyD88: IRAK-1{p}: IRAK-4{p}: TRAF6,viral ssRNA: TLR7: MyD88: IRAK-1{p}: IRAK-4{p}: TRAF6: IRF-7,viral ssRNA: TLR8,zymosan,zymosan: TLR2: dectin-1
The innate immune system serves as the first line of host defense against thedeleterious effects of invading infectious pathogens. Fever is the hallmarkamong the defense mechanisms evoked by the entry into the body of suchpathogens. The conventional view of the steps that lead to fever production isthat they begin with the biosynthesis of pyrogenic cytokines by mononuclearphagocytes stimulated by the pathogens, their release into the circulation andtransport to the thermoregulatory center in the preoptic area (POA) of theanterior hypothalamus, and their induction there of cyclooxygenase(COX)-2-dependent prostaglandin (PG)E(2), the putative final mediator of thefebrile response. But data accumulated over the past 5 years have graduallychallenged this classical concept, due mostly to the temporal incompatibility ofthe newer findings with this concatenation of events. Thus, the former studiesgenerally overlooked that the production of cytokines and the transduction oftheir pyrogenic signals into fever-mediating PGE(2) proceed at relatively slowrates, significantly slower certainly than the onset latency of fever producedby the i.v. injection of bacterial endotoxic lipopolysaccharides (LPS). Here, wereview the conflicts between the earlier and the more recent findings andsummarize new data that reconcile many of the contradictions. A unified modelbased on these data explicating the generation and maintenance of the febrileresponse is presented. It postulates that the steps in the production of LPSfever occur in the following sequence: the immediate activation by LPS of thecomplement (C) cascade, the stimulation by the anaphylatoxic C component C5a ofKupffer cells, their consequent, virtually instantaneous release of PGE(2), itsexcitation of hepatic vagal afferents, their transmission of the induced signalsto the POA via the ventral noradrenergic bundle, and the activation by the thus,locally released norepinephrine (NE) of neural alpha(1)- and glialalpha(2)-adrenoceptors. The activation of the first causes an immediate,PGE(2)-independent rise in core temperature (T(c)) [the early phase of fever; anantioxidant-sensitive PGE(2) rise, however, accompanies this first phase], andof the second a delayed, PGE(2)-dependent T(c) rise [the late phase of fever].Meanwhile-generated pyrogenic cytokines and their consequent upregulation ofblood-brain barrier cells COX-2 also contribute to the latter rise. Theconsecutive steps that initiate the febrile response to LPS would now appear,therefore, to occur in an order different than conceived originally.
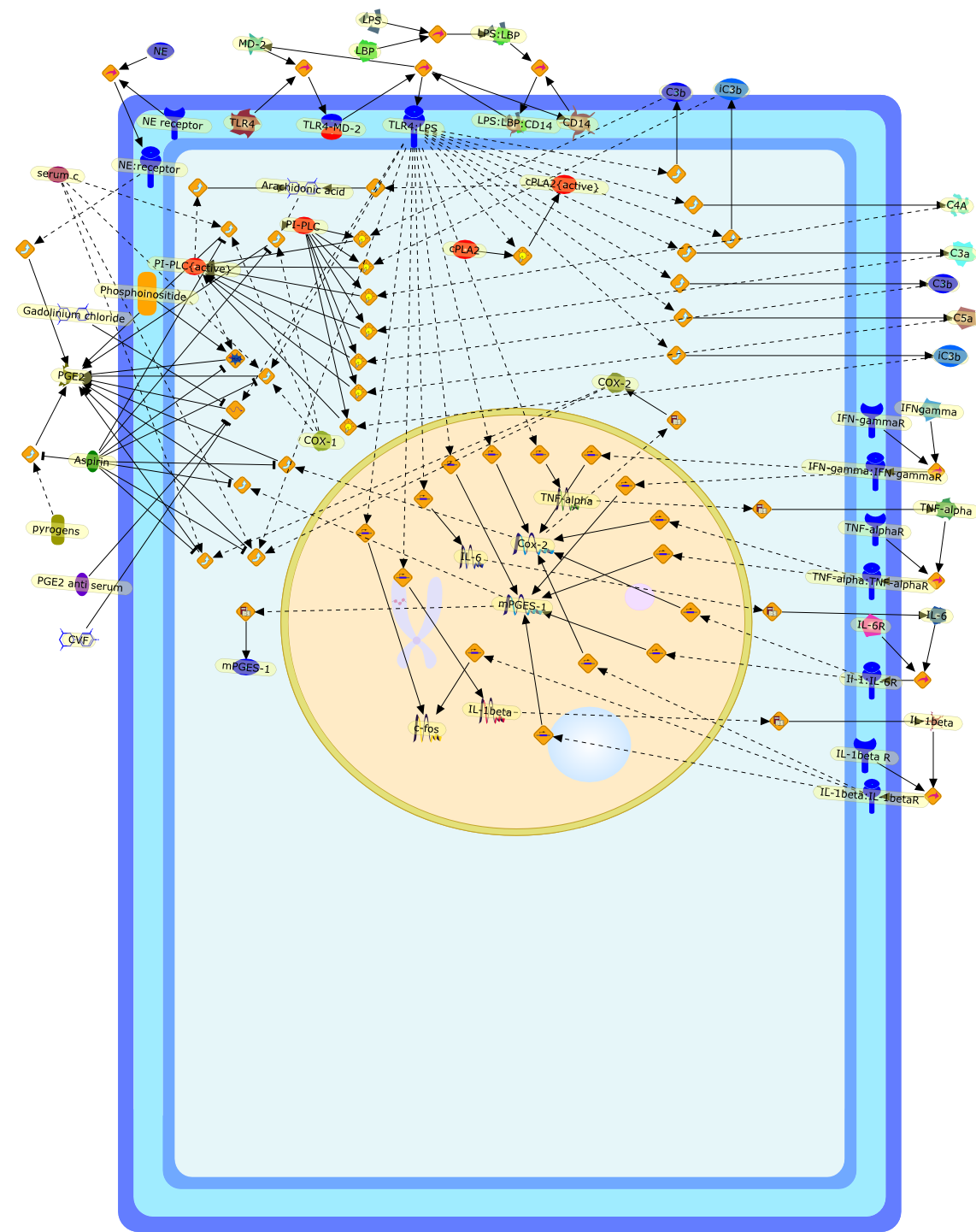{kind=link}
{kind=link}
Arachidonic acid,Aspirin,C3a,C3b,C4A,C5a,CD14,COX-1,COX-2,CVF,Cox-2,Gadolinium chloride,IFN-gamma:IFN-gammaR,IFN-gammaR,IFNgamma,IL-1beta,IL-1beta R,IL-1beta:IL-1betaR,IL-6,IL-6R,Il-1:IL-6R,LBP,LPS,LPS:LBP,LPS:LBP:CD14,MD-2,NE,NE receptor,NE:receptor,PGE2,PGE2 anti serum,PI-PLC,PI-PLC{active},Phosphoinositide,TLR4,TLR4-MD-2,TLR4:LPS,TNF-alpha,TNF-alpha:TNF-alphaR,TNF-alphaR,c-fos,cPLA2,cPLA2{active},iC3b,mPGES-1,pyrogens,serum c
Many genes, and conceivably most genes, are constitutively expressed yet haveconditional functions. Their products are utilized only under specialcircumstances, and enforce homeostatic regulation. Mutations do not disclose thefunction of such genes unless the proper conditions are applied. The genes thatencode the Toll-like receptors (TLRs) fall into this category. The TLRsrepresent the principal sensors of infection in mammals. Absent infection,mammals have little need for the TLRs; they are essential only when microbesgain access to the interior milieu of the host. The function of the TLRs inmammals was first disclosed by a spontaneous mutation in a locus called Lps,when it was shown by positional cloning to be identical to Tlr4. Random germlinemutagenesis has since permitted an estimate of the total number of proteinsrequired for TLR signaling to the level of tumor necrosis factor (TNF) synthesisand activity, and has also shown that these sensors are extremely broad in theirability to detect microbes. Ultimately, the TLRs are responsible for mostinfection-related phenomena, both good and bad. These include the development offever, shock, and tissue injury, but also the activation of innate and adaptiveeffector mechanisms that lead to the elimination of microbes.
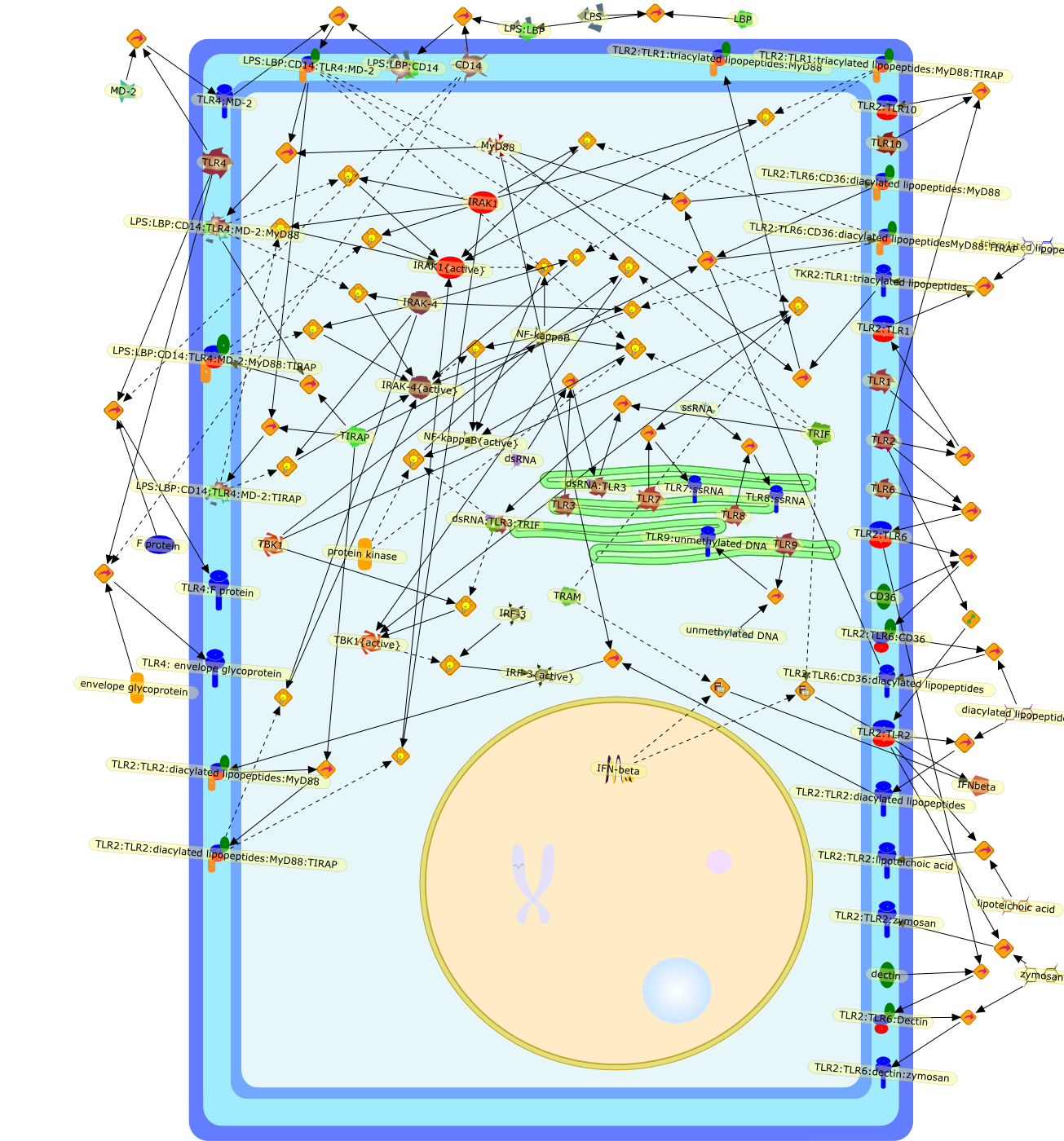{kind=link}
{kind=link}
CD14,CD36,F protein,IFN-beta,IFNbeta,IRAK-4,IRAK-4{active},IRAK1,IRAK1{active},IRF-3,IRF-3{active},LBP,LPS,LPS:LBP,LPS:LBP:CD14,LPS:LBP:CD14:TLR4:MD-2,LPS:LBP:CD14:TLR4:MD-2:MyD88,LPS:LBP:CD14:TLR4:MD-2:MyD88:TIRAP,LPS:LBP:CD14:TLR4:MD-2:TIRAP,MD-2,MyD88,NF-kappaB,NF-kappaB{active},TBK1,TBK1{active},TIRAP,TKR2:TLR1:triacylated lipopeptides,TLR1,TLR10,TLR2,TLR2:TLR1,TLR2:TLR10,TLR2:TLR1:triacylated lipopeptides:MyD88,TLR2:TLR1:triacylated lipopeptides:MyD88:TIRAP,TLR2:TLR2,TLR2:TLR2:diacylated lipopeptides,TLR2:TLR2:diacylated lipopeptides:MyD88,TLR2:TLR2:diacylated lipopeptides:MyD88:TIRAP,TLR2:TLR2:lipoteichoic acid,TLR2:TLR2:zymosan,TLR2:TLR6,TLR2:TLR6:CD36,TLR2:TLR6:CD36:diacylated lipopeptides,TLR2:TLR6:CD36:diacylated lipopeptides:MyD88,TLR2:TLR6:CD36:diacylated lipopeptidesMyD88:TIRAP,TLR2:TLR6:Dectin,TLR2:TLR6:dectin:zymosan,TLR3,TLR4,TLR4: envelope glycoprotein,TLR4:F protein,TLR4:MD-2,TLR6,TLR7,TLR7:ssRNA,TLR8,TLR8:ssRNA,TLR9,TLR9:unmethylated DNA,TRAM,TRIF,dectin,diacylated lipopeptides,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,envelope glycoprotein,lipoteichoic acid,protein kinase,ssRNA,triacylated lipopeptides,unmethylated DNA,zymosan
Toll-like receptors (TLRs) are crucially important in the sensing of infectiousagents. They serve to recognize pathogen-associated molecules and respond bytriggering the induction of specific profiles of proteins that are tailored tothe successful removal of the invading pathogens. The induction ofTLR-responsive genes is mediated by the activation of transcription factors, andmost interest has focussed on NF-kappaB, a transcription factor that isuniversally used by all TLRs. However, there has recently been a burgeoningeffort to increase our appreciation of the importance of members of theinterferon-regulatory factor (IRF) family in TLR signalling. This review willdiscuss the most recent findings relating to the regulation of IRF activity byTLRs and will highlight the rapidly increasing complexity of TLR signallingpathways.
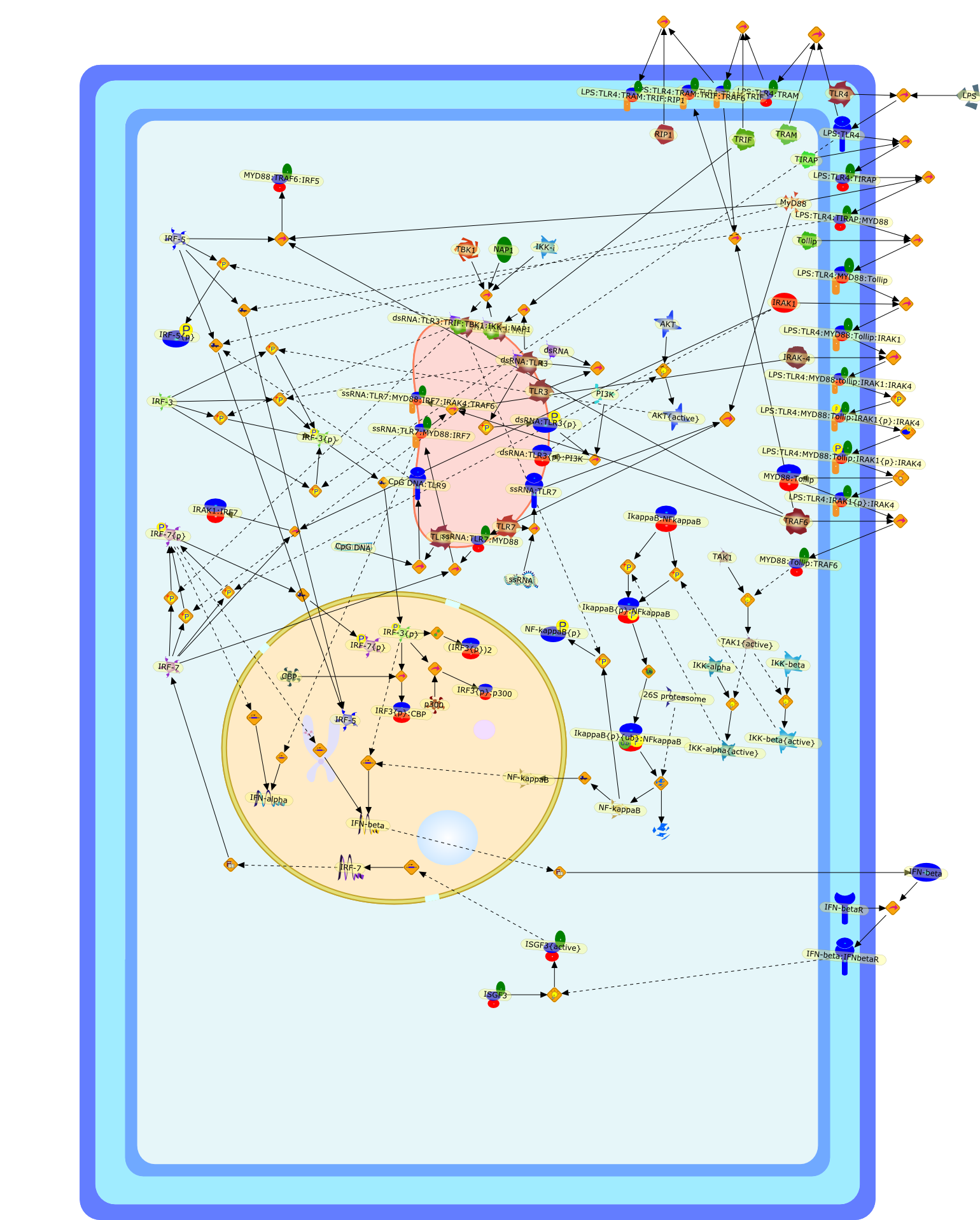{kind=link}
{kind=link}
(IRF3{p})2,26S proteasome,AKT,AKT{active},CBP,CpG DNA,CpG DNA:TLR9,IFN-alpha,IFN-beta,IFN-beta:IFNbetaR,IFN-betaR,IKK-alpha,IKK-alpha{active},IKK-beta,IKK-beta{active},IKK-i,IRAK-4,IRAK1,IRAK1:IRF7,IRF-3,IRF-3{p},IRF-5,IRF-5{p},IRF-7,IRF-7{p},IRF3{p}:CBP,IRF3{p}:p300,ISGF3,ISGF3{active},IkappaB:NFkappaB,IkappaB{p}:NFkappaB,IkappaB{p}{ub}:NFkappaB,LPS,LPS:TLR4,LPS:TLR4:IRAK1{p}:IRAK4,LPS:TLR4:MYD88:Tollip,LPS:TLR4:MYD88:Tollip:IRAK1,LPS:TLR4:MYD88:Tollip:IRAK1{p}:IRAK4,LPS:TLR4:MYD88:tollip:IRAK1:IRAK4,LPS:TLR4:TIRAP,LPS:TLR4:TIRAP:MYD88,LPS:TLR4:TRAM,LPS:TLR4:TRAM:TRIF,LPS:TLR4:TRAM:TRIF:RIP1,LPS:TLR4:TRAM:TRIF:TRAF6,MYD88:TRAF6:IRF5,MYD88:Tollip,MYD88:Tollip:TRAF6,MyD88,NAP1,NF-kappaB,NF-kappaB{p},PI3K,RIP1,TAK1,TAK1{active},TBK1,TIRAP,TLR3,TLR4,TLR7,TLR9,TRAF6,TRAM,TRIF,Tollip,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:TBK1:IKK-i:NAP1,dsRNA:TLR3{p},dsRNA:TLR3{p}:PI3K,p300,protein remnants,ssRNA,ssRNA:TLR7,ssRNA:TLR7:MYD88,ssRNA:TLR7:MYD88:IRF7,ssRNA:TLR7:MYD88:IRF7:IRAK4:TRAF6
Toll-like receptor 3 (TLR3), which recognizes double-stranded (ds)RNA, was thefirst identified antiviral TLR and, because dsRNA is a universal viral molecularpattern, TLR3 has been assumed to have a central role in the host response toviruses. However, this role has recently been questioned by in vivo studies andthe discovery of several other antiviral pattern-recognition receptors. In thisreview, the function of TLR3 in the context of these other receptors, namelyTLR7, 8 and 9 and the newly identified dsRNA-receptor retinoic-acid induciblegene-I (RIG-I) is discussed. Also, recent research concerning the expressionprofile of TLR3, its evasion by viruses and a potential role in crosspriming isaddressed, which reveals a clearer appreciation of the contribution of TLR3 toantiviral immunity.
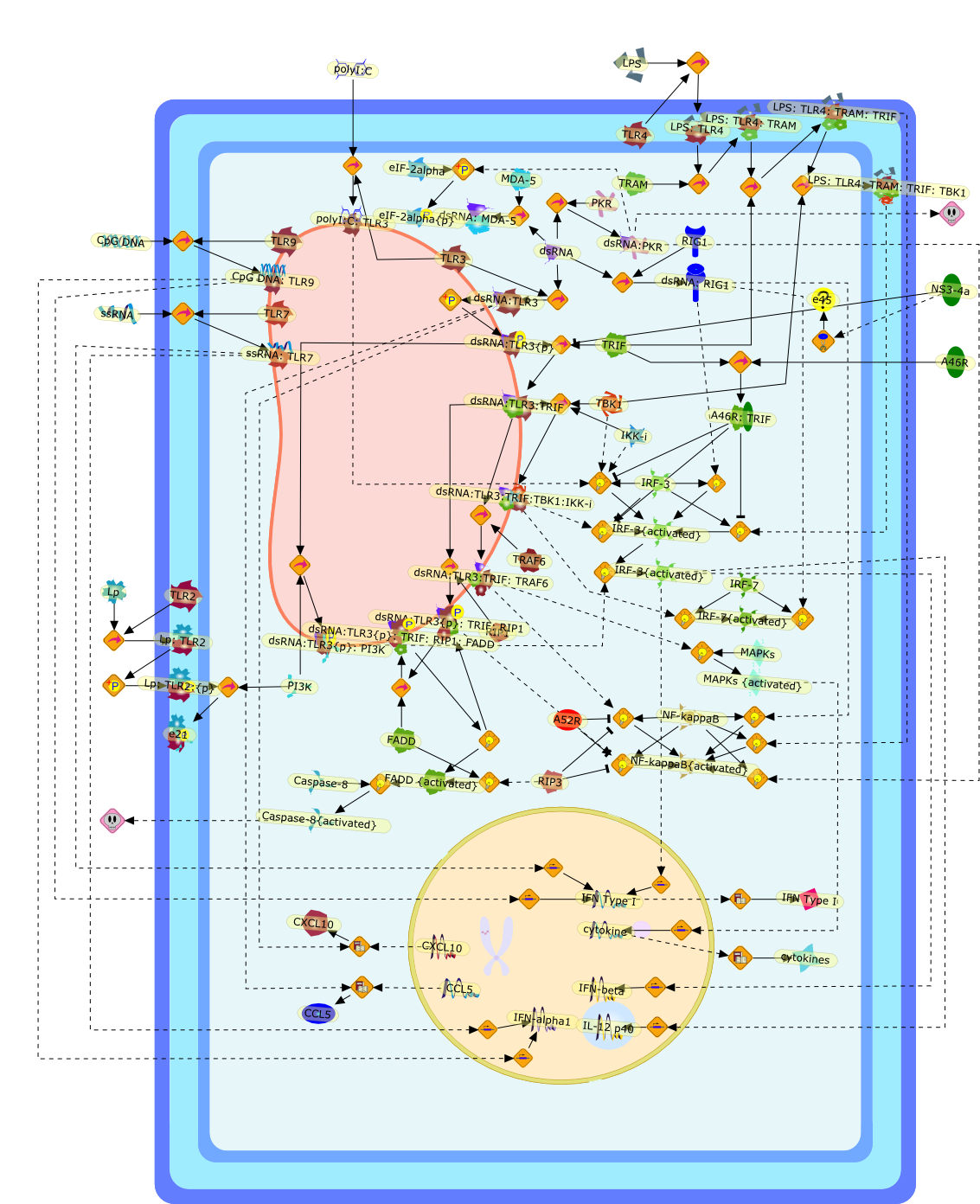{kind=link}
{kind=link}
A46R,A46R: TRIF,A52R,CCL5,CXCL10,Caspase-8,Caspase-8{activated},CpG DNA,CpG DNA: TLR9,FADD,FADD {activated},IFN Type I,IFN-alpha1,IFN-beta,IKK-i,IL-12 p40,IRF-3,IRF-3{activated},IRF-7,IRF-7{activated},LPS,LPS: TLR4,LPS: TLR4: TRAM,LPS: TLR4: TRAM: TRIF,LPS: TLR4: TRAM: TRIF: TBK1,Lp,Lp: TLR2,Lp: TLR2:{p},MAPKs,MAPKs {activated},MDA-5,NF-kappaB,NF-kappaB{activated},NS3-4a,PI3K,PKR,RIG1,RIP1,RIP3,TBK1,TLR2,TLR3,TLR4,TLR7,TLR9,TRAF6,TRAM,TRIF,cytokine,cytokines,dsRNA,dsRNA: MDA-5,dsRNA: RIG1,dsRNA:PKR,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF: TRAF6,dsRNA:TLR3:TRIF:TBK1:IKK-i,dsRNA:TLR3{p},dsRNA:TLR3{p}: PI3K,dsRNA:TLR3{p}: TRIF: RIP1,dsRNA:TLR3{p}: TRIF: RIP1: FADD,e21,e45,eIF-2alpha,eIF-2alpha{p},polyI:C,polyI:C: TLR3,ssRNA,ssRNA: TLR7
Many infectious agents elicit a type I interferon response but, until recently,the molecular details that coordinate interferon (IFN)-alpha and -betaexpression during infection were unknown. Innate immune pattern recognitionreceptors, including Toll-like receptors and cytoplasmic RNA helicases such asretinoic acid-inducible gene, that sense viral nucleic acids have beendiscovered. Using distinct mechanisms, these receptors trigger cellularsignaling pathways, culminating in the activation of interferon regulatoryfactors that transcriptionally induce IFN-alpha and IFN-beta genes.
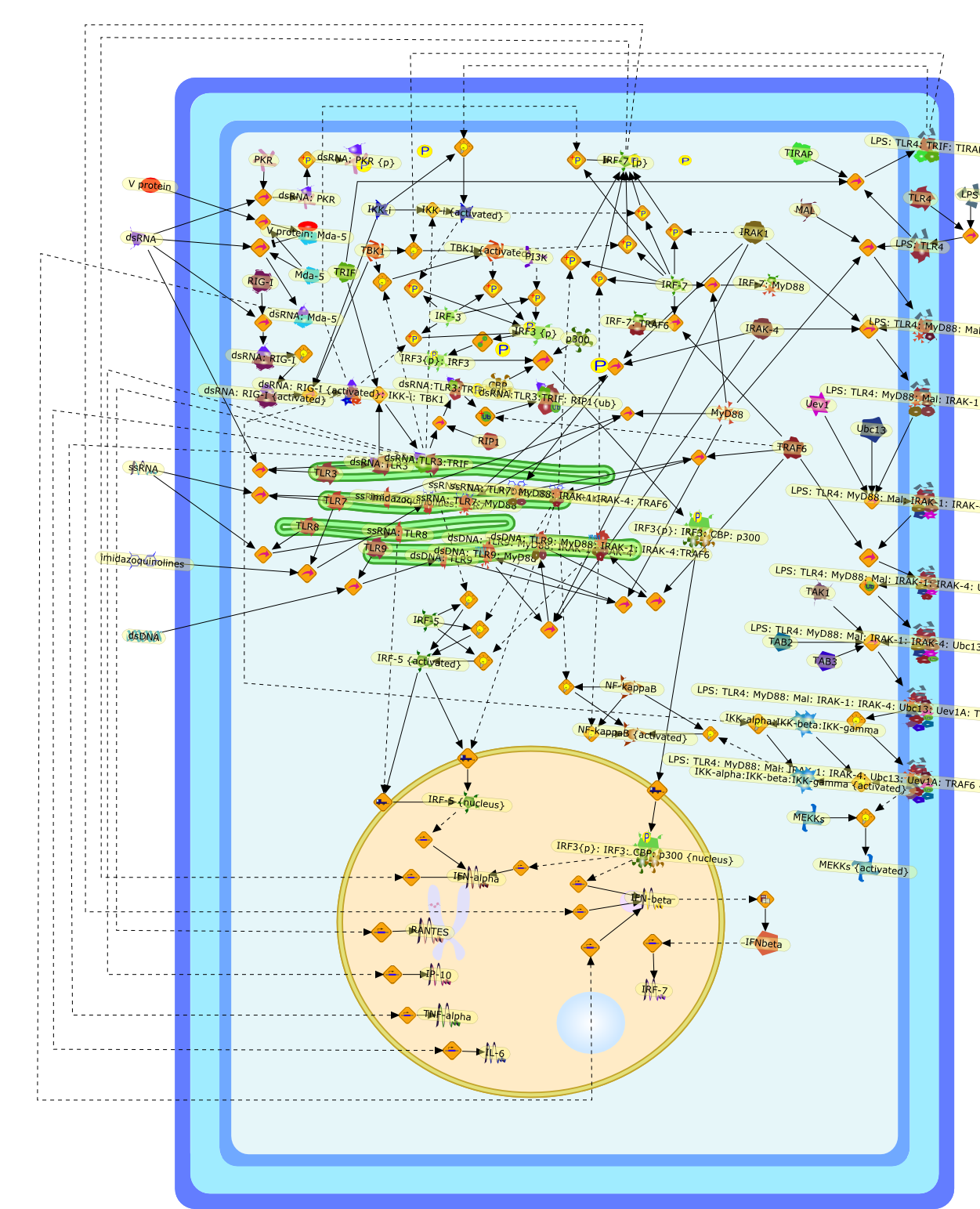{kind=link}
{kind=link}
CBP,IFN-alpha,IFN-beta,IFNbeta,IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma {activated},IKK-i,IKK-i {activated},IL-6,IP-10,IRAK-4,IRAK1,IRF-3,IRF-5,IRF-5 {activated},IRF-5 {nucleus},IRF-7,IRF-7 [p},IRF-7: MyD88,IRF-7: TRAF6,IRF3 {p},IRF3{p}: IRF3,IRF3{p}: IRF3: CBP: p300,IRF3{p}: IRF3: CBP: p300 {nucleus},LPS,LPS: TLR4,LPS: TLR4: MyD88: Mal,LPS: TLR4: MyD88: Mal: IRAK-1: IRAK-4,LPS: TLR4: MyD88: Mal: IRAK-1: IRAK-4: Ubc13: Uev1A,LPS: TLR4: MyD88: Mal: IRAK-1: IRAK-4: Ubc13: Uev1A: TRAF6,LPS: TLR4: MyD88: Mal: IRAK-1: IRAK-4: Ubc13: Uev1A: TRAF6 {ub (n)},LPS: TLR4: MyD88: Mal: IRAK-1: IRAK-4: Ubc13: Uev1A: TRAF6 {ub (n)}: TAK1: TAB2: TAB3,LPS: TLR4: MyD88: Mal: IRAK-1: IRAK-4: Ubc13: Uev1A: TRAF6 {ub (n)}: TAK1{activated}: TAB2: TAB3,LPS: TLR4: TRIF: TIRAP,MAL,MEKKs,MEKKs {activated},Mda-5,MyD88,NF-kappaB,NF-kappaB {activated},PI3K,PKR,RANTES,RIG-I,RIP1,TAB2,TAB3,TAK1,TBK1,TBK1 {activated},TIRAP,TLR3,TLR4,TLR7,TLR8,TLR9,TNF-alpha,TRAF6,TRIF,Ubc13,Uev1,V protein,V protein: Mda-5,dsDNA,dsDNA: TLR9,dsDNA: TLR9: MyD88,dsDNA: TLR9: MyD88: IRAK-1: IRAK-4,dsDNA: TLR9: MyD88: IRAK-1: IRAK-4:TRAF6,dsRNA,dsRNA: Mda-5,dsRNA: PKR,dsRNA: PKR {p},dsRNA: RIG-I,dsRNA: RIG-I {activated},dsRNA: RIG-I {activated}: IKK-i: TBK1,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF: RIP1,dsRNA:TLR3:TRIF: RIP1{ub},imidazoquinolines,imidazoquinolines: TLR7,p300,ssRNA,ssRNA: TLR7,ssRNA: TLR7: MyD88,ssRNA: TLR7: MyD88: IRAK-1:IRAK-4,ssRNA: TLR7: MyD88: IRAK-1:IRAK-4: TRAF6,ssRNA: TLR8
CpG methylation of DNA silences TLR9-mediated innate immune recognition. In thisissue of Immunity, Kariko et al (2005) suggest that the innate immunerecognition of RNA by TLR3, TLR7, or TLR8 is in fact controlled by modificationof nucleotides, including methylation.
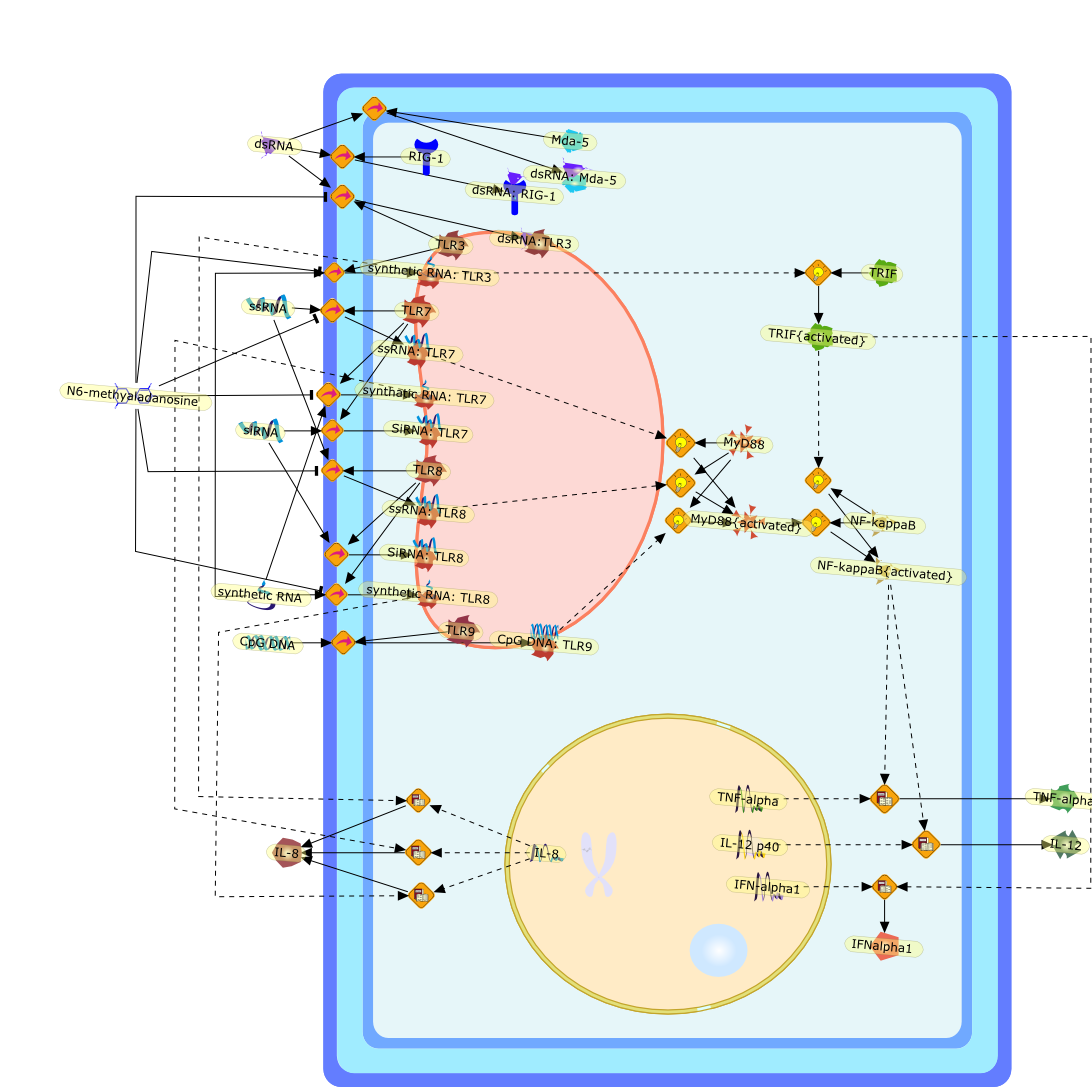{kind=link}
{kind=link}
CpG DNA,CpG DNA: TLR9,IFN-alpha1,IFNalpha1,IL-12,IL-12 p40,IL-8,Mda-5,MyD88,MyD88{activated},N6-methyaladanosine,NF-kappaB,NF-kappaB{activated},RIG-1,SiRNA: TLR7,SiRNA: TLR8,TLR3,TLR7,TLR8,TLR9,TNF-alpha,TRIF,TRIF{activated},dsRNA,dsRNA: Mda-5,dsRNA: RIG-1,dsRNA:TLR3,siRNA,ssRNA,ssRNA: TLR7,ssRNA: TLR8,synthatic RNA: TLR7,synthetic RNA,synthetic RNA: TLR3,synthetic RNA: TLR8
Classical immunoreceptors like lymphocyte antigen receptors and Fc-receptors(FcR) are central players of the adaptive immune response. These receptorsutilize a common signal transduction mechanism, which relies on immunoreceptortyrosine-based activation motifs (ITAMs) present in the receptor complex. Uponligand binding to the receptors, tyrosines within the ITAM sequence arephosphorylated by Src-family kinases, leading to an SH2-domain mediatedrecruitment and activation of the Syk or the related ZAP-70 tyrosine kinase.These kinases then initiate further downstream signaling events. Here we reviewrecent evidence indicating that components of this ITAM-based signalingmachinery are also present in a number of non-lymphoid or even non-immune celltypes and they participate in diverse biological functions beyond the adaptiveimmune response, including innate immune mechanisms, platelet activation, boneresorption or tumor development. These results suggest that the ITAM-basedsignaling paradigm has much wider implications than previously anticipated.
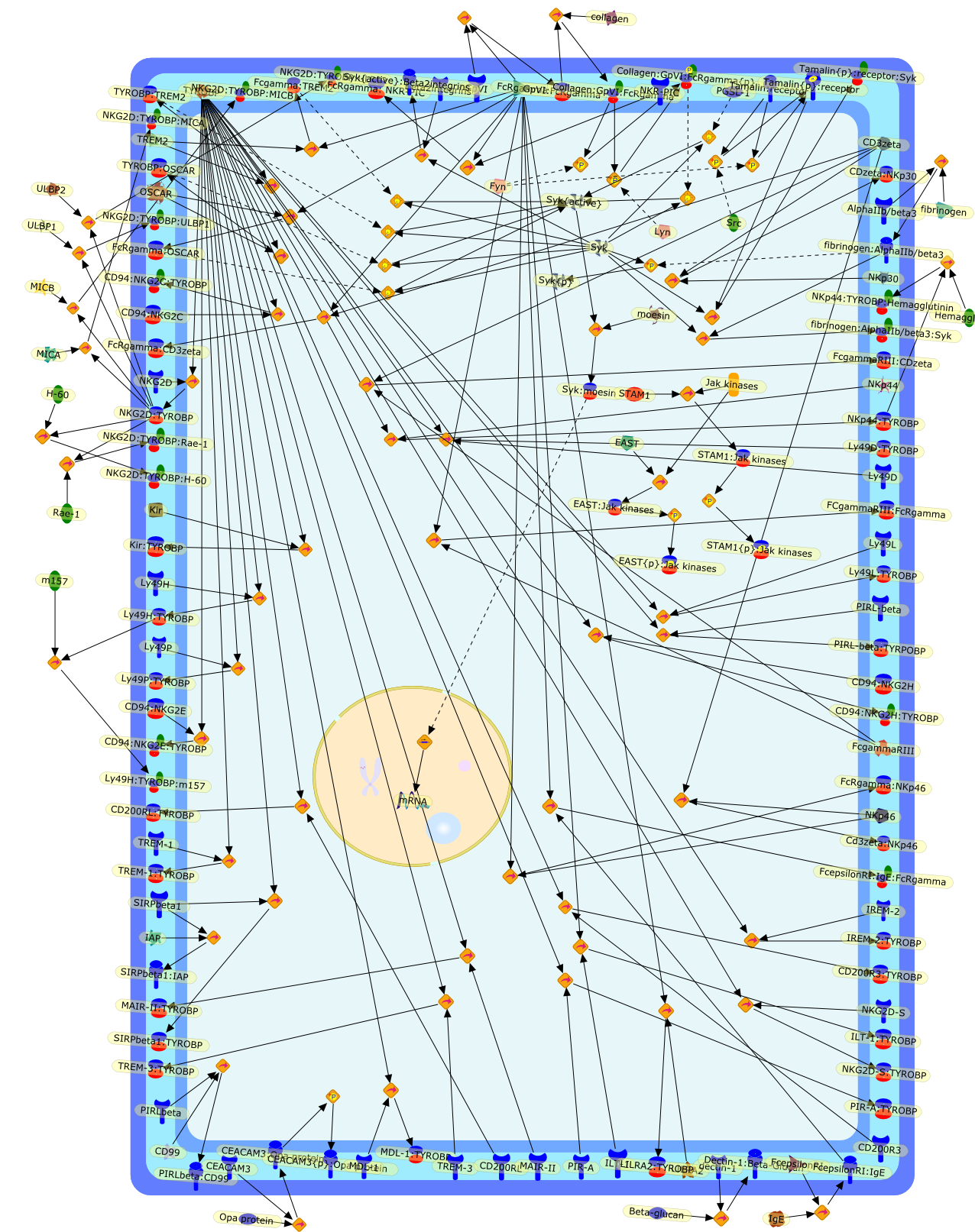{kind=link}
{kind=link}
CDzeta:NKp30,AlphaIIb/beta3,Beta-glucan,Beta2integrins,CD200R3,CD200R3:TYROBP,CD200RL,CD200RL:TYROBP,CD3zeta,CD94:NKG2C,CD94:NKG2C:TYROBP,CD94:NKG2E,CD94:NKG2E:TYROBP,CD94:NKG2H,CD94:NKG2H:TYROBP,CD99,CEACAM3,CEACAM3:Opa protein,CEACAM3{p}:Opa protein,Cd3zeta:NKp46,Collagen:GpVI:FcRgamma,Collagen:GpVI:FcRgamma{p},Dectin-1:Beta-Glucan,EAST,EAST:Jak kinases,EAST{p}:Jak kinases,FCgammaRIII:FcRgamma,FcRgamma,FcRgamma: NKR-PIC,FcRgamma:CD3zeta,FcRgamma:NKp46,FcRgamma:OSCAR,FcepsilonRI,FcepsilonRI:IgE,FcepsilonRI:IgE:FcRgamma,Fcgamma:TREM2,FcgammaRIII,FcgammaRIII:CDzeta,Fyn,GpVI,GpVI:FcRgamma,H-60,Hemagglutinin,IAP,ILT-1,ILT-1:TYROBP,IREM-2,IREM-2:TYROBP,IgE,Jak kinases,Kir,Kir:TYROBP,LILRA2,LILRA2:TYROBP,Ly49D,Ly49D:TYROBP,Ly49H,Ly49H:TYROBP,Ly49H:TYROBP:m157,Ly49L,Ly49L:TYROBP,Ly49P,Ly49P:TYROBP,Lyn,MAIR-II,MAIR-II:TYROBP,MDL-1,MDL-1:TYROBP,MICA,MICB,NKG2D,NKG2D-S,NKG2D-S:TYROBP,NKG2D:TYROBP,NKG2D:TYROBP:H-60,NKG2D:TYROBP:MICA,NKG2D:TYROBP:MICB,NKG2D:TYROBP:Rae-1,NKG2D:TYROBP:ULBP1,NKG2D:TYROBP:ULBP2,NKR-PIC,NKp30,NKp44,NKp44:TYROBP,NKp44:TYROBP:Hemagglutinin,NKp46,OSCAR,Opa protein,PGSL-1,PIR-A,PIR-A:TYROBP,PIRL-beta,PIRL-beta:TYRPOBP,PIRLbeta,PIRLbeta:CD99,Rae-1,SIRPbeta1,SIRPbeta1:IAP,SIRPbeta1:TYROBP,STAM1,STAM1:Jak kinases,STAM1{p}:Jak kinases,Src,Syk,Syk:moesin,Syk{active},Syk{active}:Beta2integrins,Syk{p},TREM-1,TREM-1:TYROBP,TREM-3,TREM-3:TYROBP,TREM2,TYROBP,TYROBP:OSCAR,TYROBP:TREM2,Tamalin:receptor,Tamalin{p}:receptor,Tamalin{p}:receptor:Syk,ULBP1,ULBP2,collagen,dectin-1,fibrinogen,fibrinogen:AlphaIIb/beta3,fibrinogen:AlphaIIb/beta3:Syk,m157,mRNA,moesin
Signal transduction pathways activated by Toll-like receptors (TLRs) havecontinued to be a major focus of research for investigators interested in theinitiation of innate immune responses and the induction of pro-inflammatorycytokines and type I interferons during infection. Biochemical details of themajor signalling pathways have now been obtained, and the specific signallingpathways activated by different TLRs are being elucidated. New insights into theactivation of IRF family members, notably IRF3, IRF5 and IRF7, have beenobtained, and interesting spatiotemporal aspects of signalling by MyD88 leadingto IRF7 activation revealed. Adapters in TLR signalling are targets forinhibition, both by endogenous regulators and by virally derived proteins.Selective targeting of pathways by anti-inflammatory glucocorticoids alsoindicates the potential for modulating TLR signalling therapeutically.
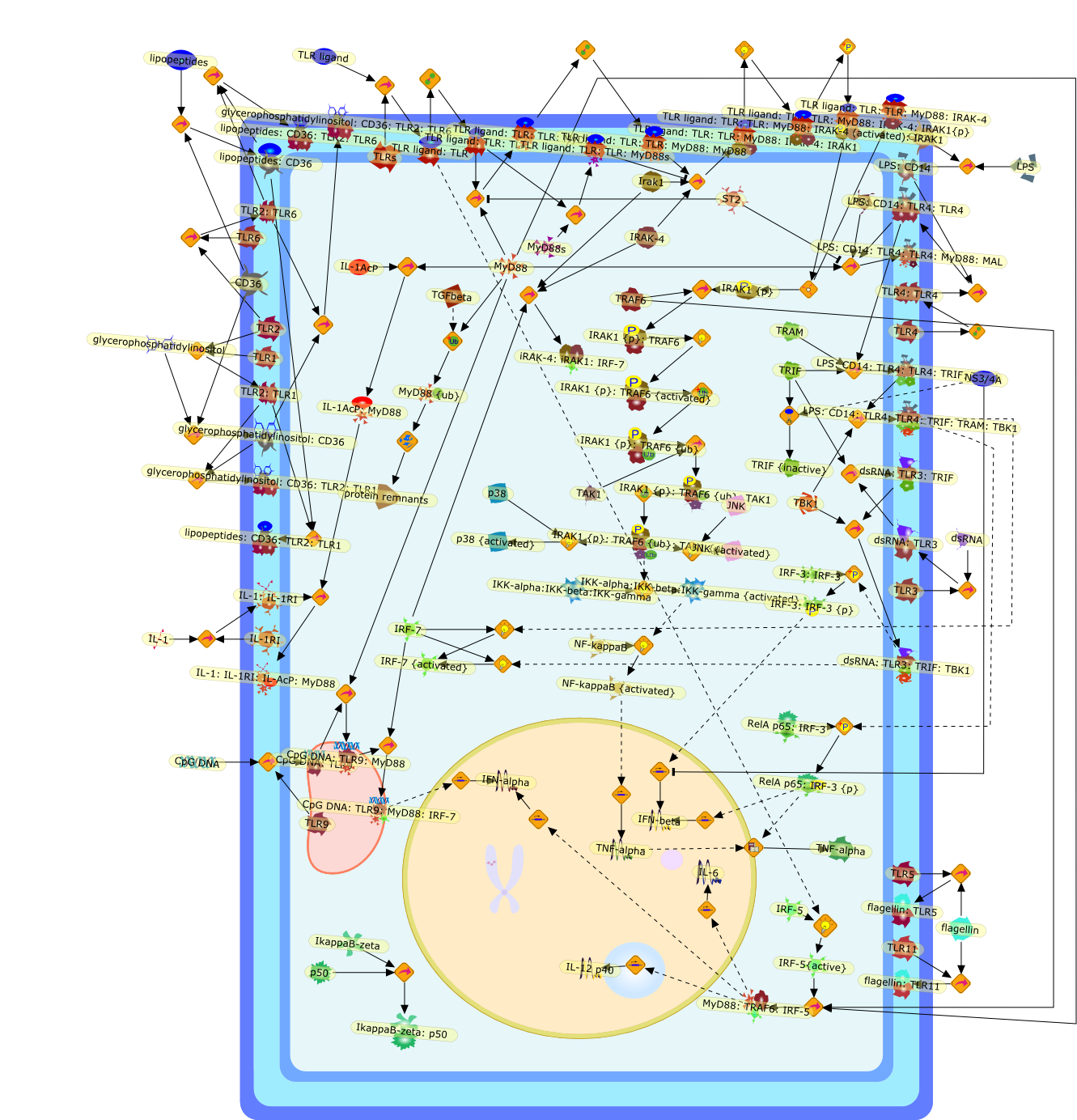{kind=link}
{kind=link}
CD14,CD36,CpG DNA,CpG DNA: TLR9,CpG DNA: TLR9: MyD88,CpG DNA: TLR9: MyD88: IRF-7,IFN-alpha,IFN-beta,IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma {activated},IL-1,IL-12 p40,IL-1: IL-1RI,IL-1: IL-1RI: IL-AcP: MyD88,IL-1AcP,IL-1AcP: MyD88,IL-1RI,IL-6,IRAK-4,IRAK1 {p},IRAK1 {p}: TRAF6,IRAK1 {p}: TRAF6 {activated},IRAK1 {p}: TRAF6 {ub},IRAK1 {p}: TRAF6 {ub}: TAK1,IRAK1 {p}: TRAF6 {ub}: TAK1 {ub},IRF-3: IRF-3,IRF-3: IRF-3 {p},IRF-5,IRF-5{active},IRF-7,IRF-7 {activated},IkappaB-zeta,IkappaB-zeta: p50,Irak1,JNK,JNK {activated},LPS,LPS: CD14,LPS: CD14: TLR4: TLR4,LPS: CD14: TLR4: TLR4: MyD88: MAL,LPS: CD14: TLR4: TLR4: TRIF: TRAM,LPS: CD14: TLR4: TLR4: TRIF: TRAM: TBK1,MAL,MyD88,MyD88 {ub},MyD88: TRAF6: IRF-5,MyD88s,NF-kappaB,NF-kappaB {activated},NS3/4A,RelA p65: IRF-3,RelA p65: IRF-3 {p},ST2,TAK1,TBK1,TGFbeta,TLR ligand,TLR ligand: TLR,TLR ligand: TLR: TLR,TLR ligand: TLR: TLR: MyD88,TLR ligand: TLR: TLR: MyD88: IRAK-4,TLR ligand: TLR: TLR: MyD88: IRAK-4 (activated): IRAK1,TLR ligand: TLR: TLR: MyD88: IRAK-4: IRAK1,TLR ligand: TLR: TLR: MyD88: IRAK-4: IRAK1{p},TLR ligand: TLR: TLR: MyD88: MyD88,TLR ligand: TLR: TLR: MyD88s,TLR1,TLR11,TLR2,TLR2: TLR1,TLR2: TLR6,TLR3,TLR4,TLR4: TLR4,TLR5,TLR6,TLR9,TLRs,TNF-alpha,TRAF6,TRAM,TRIF,TRIF {inactive},dsRNA,dsRNA: TLR3,dsRNA: TLR3: TRIF,dsRNA: TLR3: TRIF: TBK1,flagellin,flagellin: TLR11,flagellin: TLR5,glycerophosphatidylinositol,glycerophosphatidylinositol: CD36,glycerophosphatidylinositol: CD36: TLR2: TLR1,glycerophosphatidylinositol: CD36: TLR2: TLR6,iRAK-4: iRAK1: IRF-7,lipopeptides,lipopeptides: CD36,lipopeptides: CD36: TLR2: TLR1,lipopeptides: CD36: TLR2: TLR6,p38,p38 {activated},p50,protein remnants
Sepsis is the systemic immune response to severe bacterial infection. The innateimmune recognition of bacterial and viral products is mediated by a family oftransmembrane receptors known as Toll-like receptors (TLRs). In endothelialcells, exposure to lipopolysaccharide (LPS), a major cell wall constituent ofGram-negative bacteria, results in endothelial activation through a receptorcomplex consisting of TLR4, CD14 and MD2. Recruitment of the adaptor proteinmyeloid differentiation factor (MyD88) initiates an MyD88-dependent pathway thatculminates in the early activation of nuclear factor-kappaB (NF-kappaB) and themitogen-activated protein kinases. In parallel, a MyD88-independent pathwayresults in a late-phase activation of NF-kappaB. The outcome is the productionof various proinflammatory mediators and ultimately cellular injury, leading tothe various vascular sequelae of sepsis. This review will focus on the signalingpathways initiated by LPS binding to the TLR4 receptor in endothelial cells andthe coordinated regulation of this pathway.
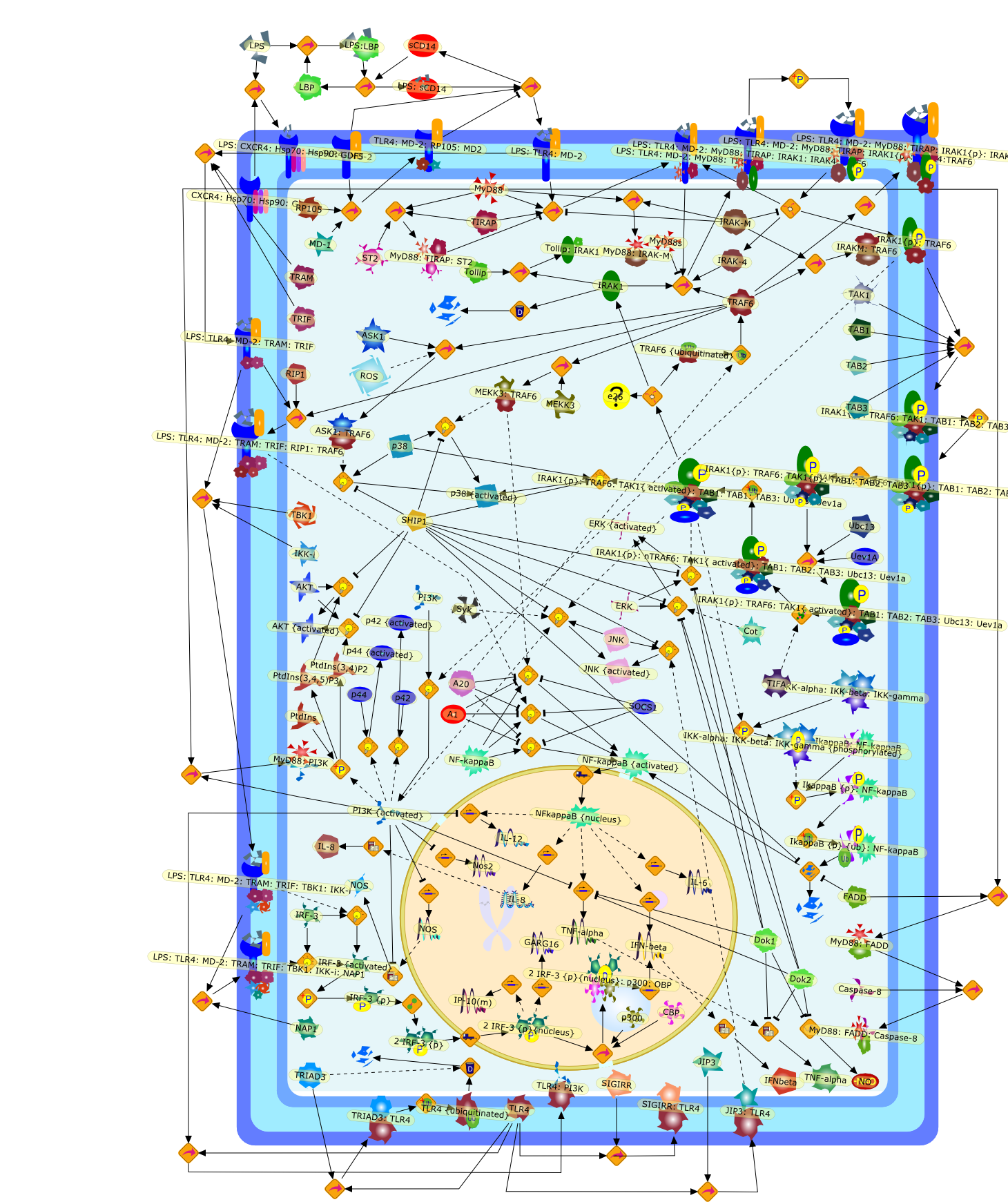{kind=link}
{kind=link}
2 IRF-3 {p},2 IRF-3 {p}{nucleus},2 IRF-3 {p}{nucleus}: p300: OBP,A1,A20,AKT,AKT {activated},ASK1,ASK1: TRAF6,CBP,CXCR4: Hsp70: Hsp90: GDF5,Caspase-8,Cot,Dok1,Dok2,ERK,ERK {activated},FADD,GARG16,IFN-beta,IFNbeta,IKK-alpha: IKK-beta: IKK-gamma,IKK-alpha: IKK-beta: IKK-gamma {phosphorylated},IKK-i,IL-12,IL-6,IL-8,IP-10(m),IRAK-4,IRAK-M,IRAK1,IRAK1 { degraded } ,IRAK1{p}: TRAF6,IRAK1{p}: TRAF6: TAK1: TAB1: TAB2: TAB3,IRAK1{p}: TRAF6: TAK1{ activated}: TAB1: TAB1: TAB3: Ubc13: Uev1a,IRAK1{p}: TRAF6: TAK1{ activated}: TAB1: TAB2: TAB3: Ubc13: Uev1a,IRAK1{p}: TRAF6: TAK1{p}: TAB1: TAB2: TAB3,IRAK1{p}: nTRAF6: TAK1{ activated}: TAB1: TAB2: TAB3: Ubc13: Uev1a,IRAKM: TRAF6,IRF-3,IRF-3 {activated},IRF-3 {p},IkappaB {degraded},IkappaB {p} {ub}: NF-kappaB,IkappaB {p}: NF-kappaB,IkappaB: NF-kappaB,JIP3,JIP3: TLR4,JNK,JNK {activated},LBP,LPS,LPS: CXCR4: Hsp70: Hsp90: GDF5,LPS: TLR4: MD-2,LPS: TLR4: MD-2: MyD88: TIRAP,LPS: TLR4: MD-2: MyD88: TIRAP: IRAK1: IRAK4:TRAF6,LPS: TLR4: MD-2: MyD88: TIRAP: IRAK1{p}: IRAK4: TRAF6,LPS: TLR4: MD-2: MyD88: TIRAP: IRAK1{p}: IRAK4:TRAF6,LPS: TLR4: MD-2: TRAM: TRIF,LPS: TLR4: MD-2: TRAM: TRIF: RIP1: TRAF6,LPS: TLR4: MD-2: TRAM: TRIF: TBK1: IKK-i,LPS: TLR4: MD-2: TRAM: TRIF: TBK1: IKK-i: NAP1,LPS: sCD14,LPS:LBP,MD-1,MEKK3,MEKK3: TRAF6,MyD88,MyD88: FADD,MyD88: FADD: Caspase-8,MyD88: IRAK-M,MyD88: PI3K,MyD88: TIRAP: ST2,MyD88s,NAP1,NF-kappaB,NF-kappaB {activated},NFkappaB {nucleus},NO,NOS,Nos2,PI3K,PI3K {activated},PtdIns,PtdIns(3,4)P2,PtdIns(3,4,5)P3,RIP1,ROS,RP105,SHIP1,SIGIRR,SIGIRR: TLR4,SOCS1,ST2,Syk,TAB1,TAB2,TAB3,TAK1,TBK1,TIFA,TIRAP,TLR4,TLR4 {degraded},TLR4 {ubiquitinated},TLR4: MD-2,TLR4: MD-2: RP105: MD2,TLR4: PI3K,TNF-alpha,TRAF6,TRAF6 {ubiquitinated},TRAM,TRIAD3,TRIAD3: TLR4,TRIF,Tollip,Tollip: IRAK1,Ubc13,Uev1A,e26,p300,p38,p38 {activated},p42,p42 {activated},p44,p44 {activated},sCD14
The Toll-like receptor (TLR) family plays an instructive role in innate immuneresponses against microbial pathogens, as well as the subsequent induction ofadaptive immune responses. TLRs recognize specific molecular patterns found in abroad range of microbial pathogens such as bacteria and viruses, triggeringinflammatory and antiviral responses and dendritic cell maturation, which resultin the eradication of invading pathogens. Individual TLRs interact withdifferent combinations of adapter proteins and activate various transcriptionfactors such as nuclear factor (NF)-kappaB, activating protein-1 and interferonregulatory factors, driving a specific immune response. This review outlines therecent advances in our understanding of TLR-signaling pathways and their rolesin immune responses. Further, we also discuss a new concept of TLR-independentmechanisms for recognition of microbial pathogens.
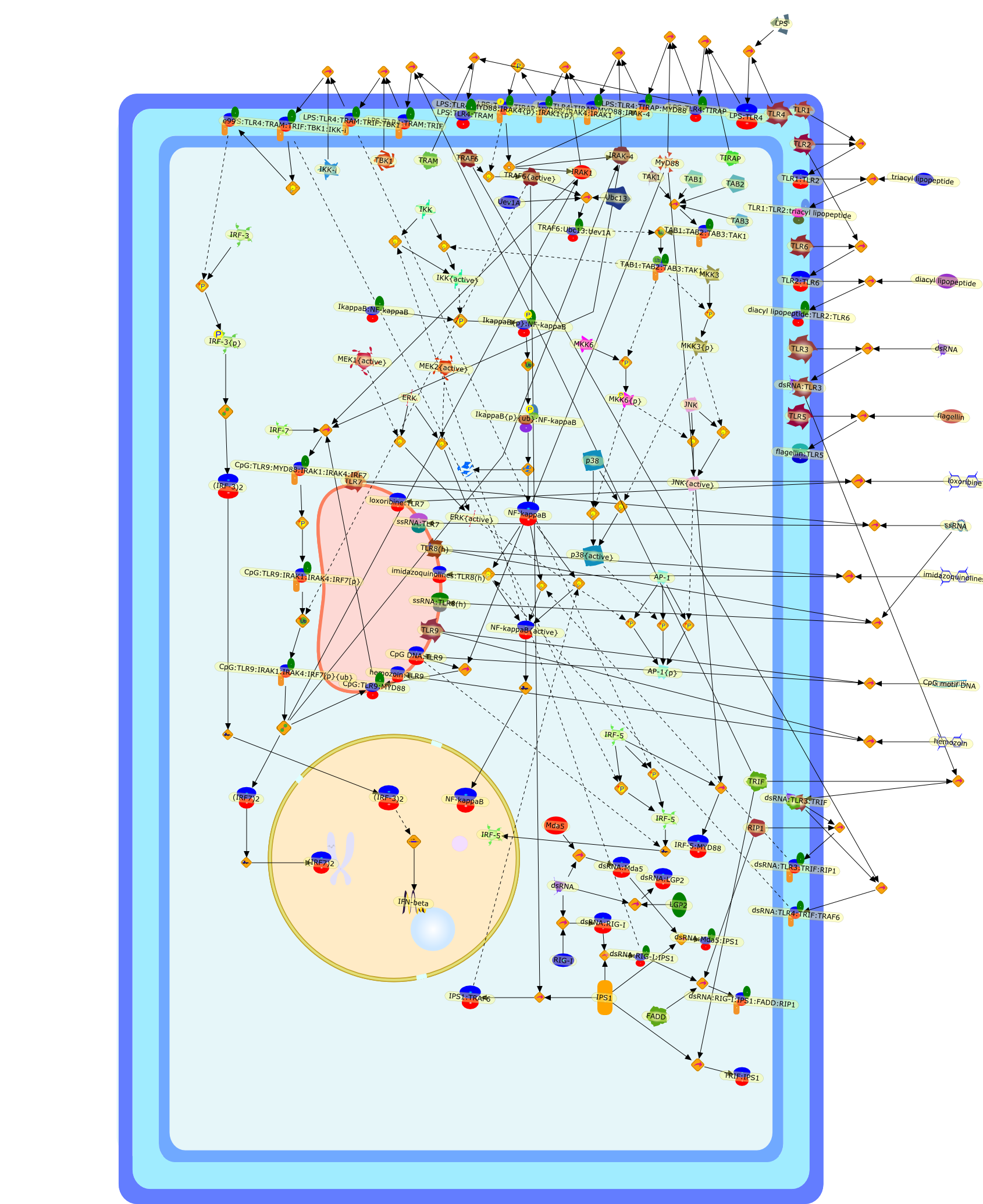{kind=link}
{kind=link}
(IRF-3)2,(IRF7)2,AP-1,AP-1{p},CpG DNA:TLR9,CpG motif DNA,CpG:TLR9:IRAK1:IRAK4:IRF7[p},CpG:TLR9:IRAK1:IRAK4:IRF7[p}{ub},CpG:TLR9:MYD88,CpG:TLR9:MYD88:IRAK1:IRAK4:IRF7,ERK,ERK{active},FADD,IFN-beta,IKK,IKK-i,IKK{active},IPS1,IPS1:TRAF6,IRAK-4,IRAK1,IRF-3,IRF-3{p},IRF-5,IRF-5:MYD88,IRF-7,IkappaB:NF-kappaB,IkappaB{p}:NF-kappaB,IkappaB{p}{ub}:NF-kappaB,JNK,JNK{active},LGP2,LPS,LPS:TLR4,LPS:TLR4:MYD88:IRAK4{p}:IRAK1{p},LPS:TLR4:TIRAP,LPS:TLR4:TIRAP:MYD88,LPS:TLR4:TIRAP:MYD88:IRAK-4,LPS:TLR4:TIRAP:MYD88:IRAK4:IRAK1,LPS:TLR4:TRAM,LPS:TLR4:TRAM:TRIF,LPS:TLR4:TRAM:TRIF:TBK1,LPS:TLR4:TRAM:TRIF:TBK1:IKK-i,MEK1{active},MEK2{active},MKK3,MKK3{p},MKK6,MKK6{p},Mda5,MyD88,NF-kappaB,NF-kappaB{active},RIG-I,RIP1,TAB1,TAB1:TAB2:TAB3:TAK1,TAB2,TAB3,TAK1,TBK1,TIRAP,TLR1,TLR1:TLR2,TLR1:TLR2:triacyl lipopeptide,TLR2,TLR2:TLR6,TLR3,TLR4,TLR5,TLR6,TLR7,TLR8(h),TLR9,TRAF6,TRAF6:Ubc13:Uev1A,TRAF6{active},TRAM,TRIF,TRIF:IPS1,Ubc13,Uev1A,diacyl lipopeptide,diacyl lipopeptide:TLR2:TLR6,dsRNA,dsRNA:LGP2,dsRNA:Mda5,dsRNA:Mda5:IPS1,dsRNA:RIG-I,dsRNA:RIG-I:IPS1,dsRNA:RIG-I:IPS1:FADD:RIP1,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:RIP1,dsRNA:TLR4:TRIF:TRAF6,e99,flagellin,flagellin:TLR5,hemozoin,hemozoin:TLR9,imidazoquinolines,imidazoquinolines:TLR8(h),loxoribine,loxoribine:TLR7,p38,p38{active},protein remnants,ssRNA,ssRNA:TLR7,ssRNA:TLR8(h),triacyl lipopeptide
The innate immune system senses invading microorganisms by a phylogeneticallyconserved family of proteins--TLRs. They are expressed in several types of cellsthat represent a route of entry of pathogens into the host organism and that cancontribute to protection against infection. Except for cells of the immunesystem, TLRs are present in epithelial cells of the skin, respiratory,intestinal, and genitourinary tracts that form the first protective barrier toinvading pathogens. Polarized regulation of TLR expression in epithelial cellsexplains why pathogenic but not commensal bacteria elicit inflammatoryresponses. TLR-induced intracellular signalling pathways show remarkablecomplexity: apart from a common signalling pathway, additional signallingpathways specific for each of the TLRs are responsible for a fine tuning of theimmune response, thus securing effective pathogen-directed biological responses.
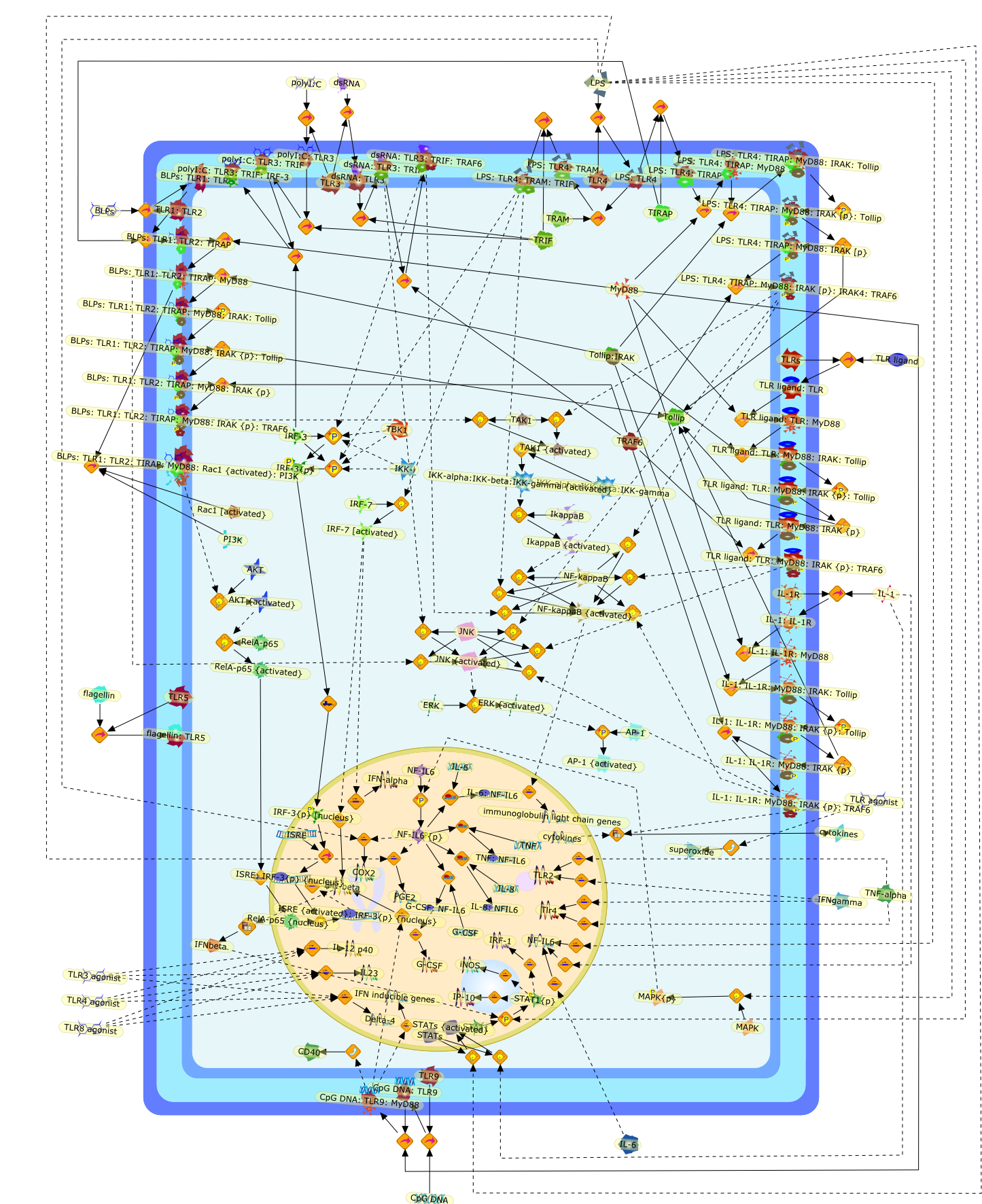{kind=link}
{kind=link}
AKT,AKT {activated},AP-1,AP-1 {activated},BLPs,BLPs: TLR1: TLR2,BLPs: TLR1: TLR2: TIRAP,BLPs: TLR1: TLR2: TIRAP: MyD88,BLPs: TLR1: TLR2: TIRAP: MyD88: IRAK {p},BLPs: TLR1: TLR2: TIRAP: MyD88: IRAK {p}: TRAF6,BLPs: TLR1: TLR2: TIRAP: MyD88: IRAK {p}: Tollip,BLPs: TLR1: TLR2: TIRAP: MyD88: IRAK: Tollip,BLPs: TLR1: TLR2: TIRAP: MyD88: Rac1 {activated}: PI3K,CD40,COX2,CpG DNA,CpG DNA: TLR9,CpG DNA: TLR9: MyD88,Delta-4,ERK,ERK {activated},G-CSF,G-CSF: NF-IL6,IFN inducible genes,IFN-alpha,IFN-beta,IFNbeta,IFNgamma,IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma {activated},IKK-i,IL-1,IL-12 p40,IL-1: IL-1R,IL-1: IL-1R: MyD88,IL-1: IL-1R: MyD88: IRAK {p},IL-1: IL-1R: MyD88: IRAK {p}: TRAF6,IL-1: IL-1R: MyD88: IRAK {p}: Tollip,IL-1: IL-1R: MyD88: IRAK: Tollip,IL-1R,IL-6,IL-6: NF-IL6,IL-8,IL-8: NFIL6,IL23,IP-10,IRF-1,IRF-3,IRF-3{p},IRF-3{p} [nucleus},IRF-7,IRF-7 [activated},ISRE,ISRE {activated}: IRF-3{p} {nucleus},ISRE: IRF-3{p} {nucleus},IkappaB,IkappaB {activated},JNK,JNK {activated},LPS,LPS: TLR4,LPS: TLR4: TIRAP,LPS: TLR4: TIRAP: MyD88,LPS: TLR4: TIRAP: MyD88: IRAK [p},LPS: TLR4: TIRAP: MyD88: IRAK [p}: IRAK4: TRAF6,LPS: TLR4: TIRAP: MyD88: IRAK [p}: Tollip,LPS: TLR4: TIRAP: MyD88: IRAK: Tollip,LPS: TLR4: TRAM,LPS: TLR4: TRAM: TRIF,MAPK,MAPK{p},MyD88,NF-IL6,NF-IL6 {p},NF-kappaB,NF-kappaB {activated},PGE2,PI3K,Rac1 [activated},RelA-p65,RelA-p65 {activated},RelA-p65 {nucleus} ,STAT1,STAT1{p},STATs,STATs {activated},TAK1,TAK1 {activated},TBK1,TIRAP,TLR agonist,TLR ligand,TLR ligand: TLR,TLR ligand: TLR: MyD88,TLR ligand: TLR: MyD88: IRAK {p},TLR ligand: TLR: MyD88: IRAK {p}: TRAF6,TLR ligand: TLR: MyD88: IRAK {p}: Tollip,TLR ligand: TLR: MyD88: IRAK: Tollip,TLR1: TLR2,TLR2,TLR3,TLR3 agonist,TLR4,TLR4 agonist,TLR5,TLR8 agonist,TLR9,TLRs,TNF,TNF-alpha,TNF: NF-IL6,TRAF6,TRAM,TRIF,Tlr4,Tollip,Tollip:IRAK,cytokines,dsRNA,dsRNA: TLR3,dsRNA: TLR3: TRIF,dsRNA: TLR3: TRIF: TRAF6,flagellin,flagellin: TLR5,iNOS,immunoglobulin light chain genes,polyI:C,polyI:C: TLR3,polyI:C: TLR3: TRIF,polyI:C: TLR3: TRIF: IRF-3,superoxide
Immunological defense depends on the ability of the innate immune system torecognize invading microbes as foreign and thus eliminate them. The Toll-likereceptors (TLRs) help detect foreign invaders by sensing variouspathogen-associated molecules, including microbial RNA and DNA. At present, itis unclear whether and how the immune system distinguishes between microbial andself nucleic acids, as host-derived RNA and DNA also stimulate TLRs. Inaddition, recent studies have revealed the existence of TLR-independent pathwaysthat are activated in response to microbial and host nucleic acids.
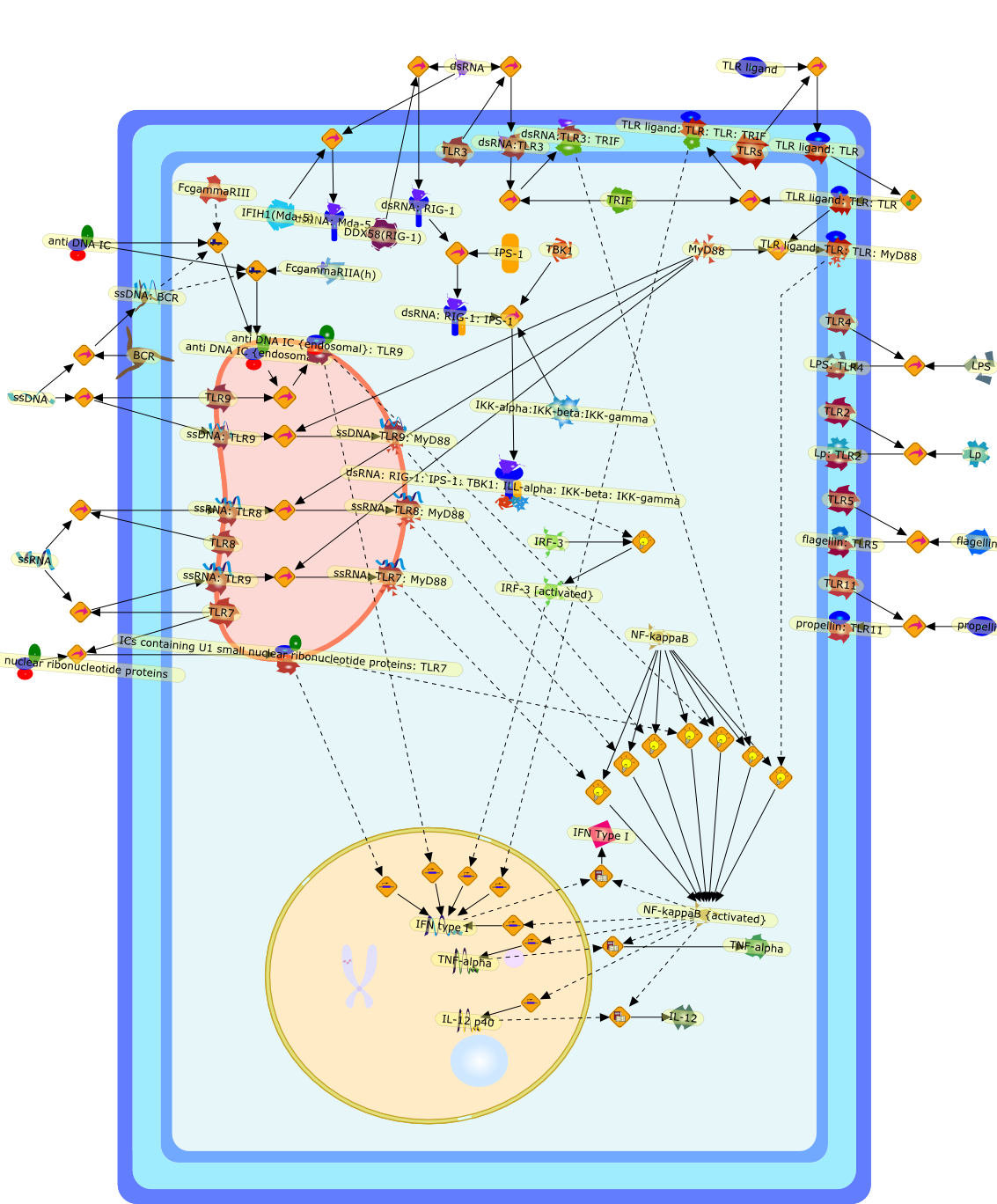{kind=link}
{kind=link}
BCR,DDX58(RIG-1),FcgammaRIIA(h),FcgammaRIII,ICs containing U1 small nuclear ribonucleotide proteins,ICs containing U1 small nuclear ribonucleotide proteins: TLR7,IFIH1(Mda-5),IFN Type I,IFN type I,IKK-alpha:IKK-beta:IKK-gamma,IL-12,IL-12 p40,IPS-1,IRF-3,IRF-3 [activated},LPS,LPS: TLR4,Lp,Lp: TLR2,MyD88,NF-kappaB,NF-kappaB {activated},TBK1,TLR ligand,TLR ligand: TLR,TLR ligand: TLR: TLR,TLR ligand: TLR: TLR: MyD88,TLR ligand: TLR: TLR: TRIF,TLR11,TLR2,TLR3,TLR4,TLR5,TLR7,TLR8,TLR9,TLRs,TNF-alpha,TRIF,anti DNA IC,anti DNA IC {endosomal},anti DNA IC {endosomal}: TLR9,dsRNA,dsRNA: Mda-5,dsRNA: RIG-1,dsRNA: RIG-1: IPS-1,dsRNA: RIG-1: IPS-1: TBK1: ILL-alpha: IKK-beta: IKK-gamma,dsRNA:TLR3,dsRNA:TLR3: TRIF,flagellin,flagellin: TLR5,propellin,propellin: TLR11,ssDNA,ssDNA: BCR,ssDNA: TLR9,ssDNA: TLR9: MyD88,ssRNA,ssRNA: TLR7: MyD88,ssRNA: TLR8,ssRNA: TLR8: MyD88,ssRNA: TLR9
Recent studies have uncovered two signaling pathways that activate the hostinnate immunity against viral infection. One of the pathways utilizes members ofthe Toll-like receptor (TLR) family to detect viruses that enter the endosomethrough endocytosis. The TLR pathway induces interferon production throughseveral signaling proteins that ultimately lead to the activation of thetranscription factors NF-kappaB, IRF3 and IRF7. The other antiviral pathway usesthe RNA helicase RIG-I as the receptor for intracellular viral double-strandedRNA. RIG-I activates NF-kappaB and IRFs through the recently identified adaptorprotein MAVS, a CARD domain containing protein that resides in the mitochondrialmembrane. MAVS is essential for antiviral innate immunity, but it also serves asa target of Hepatitis C virus (HCV), which employs a viral protease to cleaveMAVS off the mitochondria, thereby allowing HCV to escape the host immunesystem.
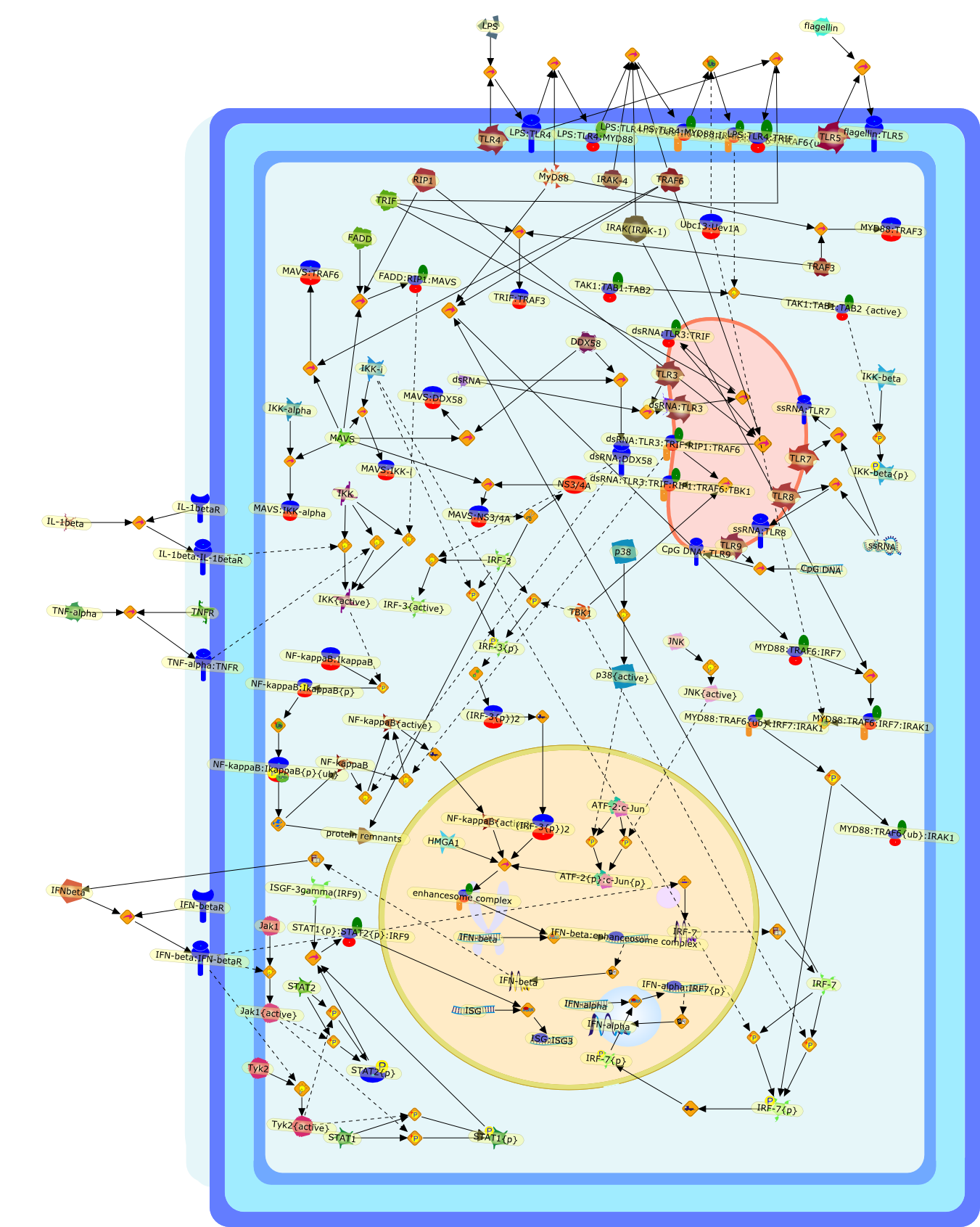{kind=link}
{kind=link}
(IRF-3{p})2,ATF-2:c-Jun,ATF-2{p}:c-Jun{p},CpG DNA,CpG DNA: TLR9,DDX58,FADD,FADD:RIP1:MAVS,HMGA1,IFN-alpha,IFN-alpha:IRF7{p},IFN-beta,IFN-beta:IFN-betaR,IFN-beta:enhanceosome complex,IFN-betaR,IFNbeta,IKK,IKK-alpha,IKK-beta,IKK-beta{p},IKK-i,IKK{active},IL-1beta,IL-1beta:IL-1betaR,IL-1betaR,IRAK(IRAK-1),IRAK-4,IRF-3,IRF-3{active},IRF-3{p},IRF-7,IRF-7{p},ISG,ISG:ISG3,ISGF-3gamma(IRF9),JNK,JNK{active},Jak1,Jak1{active},LPS,LPS:TLR4,LPS:TLR4:MYD88,LPS:TLR4:MYD88:IRAK1:IRAK4:TRAF6,LPS:TLR4:MYD88:IRAK1:IRAK4:TRAF6{ub},LPS:TLR4:TRIF,MAVS,MAVS:DDX58,MAVS:IKK-alpha,MAVS:IKK-i,MAVS:NS3/4A,MAVS:TRAF6,MYD88:TRAF3,MYD88:TRAF6:IRF7,MYD88:TRAF6:IRF7:IRAK1,MYD88:TRAF6{ub}:IRAK1,MYD88:TRAF6{ub}:IRF7:IRAK1,MyD88,NF-kappaB,NF-kappaB:IkappaB,NF-kappaB:IkappaB{p},NF-kappaB:IkappaB{p}{ub},NF-kappaB{active},NS3/4A,RIP1,STAT1,STAT1{p},STAT1{p}:STAT2{p}:IRF9,STAT2,STAT2{p},TAK1:TAB1:TAB2,TAK1:TAB1:TAB2 {active},TBK1,TLR3,TLR4,TLR5,TLR7,TLR8,TLR9,TNF-alpha,TNF-alpha:TNFR,TNFR,TRAF3,TRAF6,TRIF,TRIF:TRAF3,Tyk2,Tyk2{active},Ubc13:Uev1A,dsRNA,dsRNA:DDX58,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:RIP1:TRAF6,dsRNA:TLR3:TRIF:RIP1:TRAF6:TBK1,enhancesome complex,flagellin,flagellin:TLR5,p38,p38{active},protein remnants,ssRNA,ssRNA:TLR7,ssRNA:TLR8
Toll-like receptors (TLRs) have an important role in innate immunity in mammalsby recognizing conserved microbial components that are known aspathogen-associated molecular patterns (PAMPs). Although the majority of thesereceptors sense pathogen components on the cell surface, a subset of them (TLR3,TLR7, TLR8 and TLR9) senses viral and bacterial nucleic acids in endosomalcompartments. Of considerable interest is the recent finding that TLR7 and TLR8can also recognize small interfering RNA (siRNA), which is the main effector inRNA interference. This immune activation by siRNAs can be abrogated by the2'-ribose modification of uridines. Here, we discuss the recent developmentsthat have expanded the understanding of self-non-self discrimination of RNAs bythe innate immune system, and consider future directions for therapeuticapplications of these findings.
{kind=link}
{kind=link}
CpG DNA,CpG DNA:TLR9,CpG DNA:TLR9:MYD88,DDX58,IFN-alpha,IFN-alpha:IFNAR,IFN-beta,IFN-beta:IFNBr,IFNAR,IFNBR,IFn-alpha,IRAK,IRAK{active},IRF-3,IRF-3{active},IRF-7,IRF-7{active},ISG20,LPS,LPS:TLR4,LPS:TLR4:MYD88,MCP1,MyD88,NF-kappaB,NF-kappaB{active},OAS1,PKR,Peptidoglycan:TLR2,R-848,R848:TLR7,TLR2,TLR3,TLR4,TLR7,TLR8,TLR9,TNF-alpha,TRAF6,TRAF6{active},TRIF,bropirimine,bropirimine:TLR7,dsRNA,dsRNA:DDX58,dsRNA:PKR,dsRNA:PKR{p},dsRNA:TLR3,dsRNA:TLR3:TRIF,eIF2-alpha,eIF2-alpha{p},envelope protein,envelope protein:TLR4,imiquimod,imiquimod:TLR7,loxoribine,loxoribine:TLR7,peptidoglycan,peptidoglycan:TLR2:MYD88,siRNA,siRNA:TLR7,siRNA:TLR8,ssRNA,ssRNA:TLR7,ssRNA:TLR7:MYD88,ssRNA:TLR8,ssRNA:TLR8:MYD88,type I IFN
Toll-like receptor (TLR) activation is dictated by a number of factors includingthe ligand itself and the localization of the receptor, in terms of expressionprofile and subcellular localization and the signal transduction pathway thathas been activated. Recent work into TLR signal transduction has revealedcomplex regulation at a number of different levels including regulation byphosphorylation, targeted degradation, and sequestration of signaling molecules.Here, we describe recent advances that have been made in our understanding ofhow TLR signaling is regulated at the biochemical level.
{kind=link}
{kind=link}
26S proteasome,A20,Btk,Caspase-8,CpG DNA,CpG DNA:TLR9,CpG DNA:TLR9:TRIAD3A,DNA:{IRF-3(p)}2,Fliih,IFN-alpha,IFN-beta,IKK,IKK-i,IKK-i:TRAF3,IKK{active},IL-1beta,IRAK,IRAK-4,IRAK1,IRAK1c,IRAK1c:IRAK2:MYD88:TRAF6:Tollip,IRAK1{p}:(TRAF6)n,IRAK1{p}:TRAF6{ub}n,IRAK2,IRAK2c,IRAK{p}:TRAF6,IRF-3,IRF-3{p},IkappaB:NF-kappaB,LPS,LPS:TLR4,LPS:TLR4:TIRAP,LPS:TLR4:TIRAP:IRAK4:IRAK,LPS:TLR4:TIRAP:IRAK4:IRAK:Tollip,LPS:TLR4:TIRAP:IRAK4:IRAK{p}:Tollip:TRAF6,LPS:TLR4:TIRAP:IRAK4:IRAK{p}:Tollip{p}:TRAF6,LPS:TLR4:TIRAP:MYD88,LPS:TLR4:TIRAP:MYD88:IRAK4,LPS:TLR4:TRAM,LPS:TLR4:TRAM:TRIF,LPS:TLR4:TRAM:TRIF:RIP1:TRAF6,LPS:TLR4:TRAM:TRIF:TBK1:IKK-i,LPS:TLR4:TRIAD3A,LPS:TLR4{ub},MYD88:Fliih,MYD88:MYD88s,MYD88:MYD88s:IRAK,MYD88:TRAF1,MYD88s,MYD88{p},Mal:TRAF1,Mal:TRAF6,Mdm2,Mdm2:beta-arrestin,MyD88,NF-kappaB,NF-kappaB{active},PKR,PKR:TRAF3,R848,RIG-1,RIP1,SOCS-1,ST2L,ST2L:MYD88,ST2L:TIRAP,Socs-1:TIRAP{p}{ub},TAB1,TAB2,TAB3,TAK1,TAK1:TAB1:TAB2:TAB3,TAK1:TAB1:TAB2:TAB3{active},TBK1,TGF-beta,TIRAP,TIRAP{p},TLR11,TLR1:TLR2:TIRAP,TLR1:TLR2:TIRAP:MYD88,TLR2:TLR1,TLR2:TLR6,TLR2:TLR6:TIRAP,TLR2:TLR6:TIRAP:MYD88,TLR3,TLR4,TLR5,TLR7,TLR8,TLR9,TRAF1,TRAF1(164���416),TRAF1(1���163),TRAF1:TRIF,TRAF3,TRAF3:IRAK,TRAF3:TBK1,TRAF3:TRIF,TRAF6,TRAM,TRAM{p},TRIF,TRIF{p},Tollip,Triad3A,beta-arrestin,beta-arrestin:TRAF6,diacyl lipopreteins:TLR6:TLR2,diacyl lipoproteins,dsRNA,dsRNA:RIG-1,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:TRAF6,flagellin,flagellin:TLR5,imidazoquinoline,imidazoquinoline-like molecule,imidazoquinoline-like molecule:TLR7,imidazoquinoline:TLR8,loxoribine,loxoribine:TLR7,profilin like protein:TLR11,profilin-like protein,protein remnants,socs-1:Mal{p},src kinase,src kinase:beta arrestin,ssRNA,ssRNA:TLR7,ssRNA:TLR8,triacyl lipoprotein,triacyl lipoprotein:TLR1:TLR2,type I IFN,type I IFN DNA,type I IFNs,{IRF-3(p)}2
Viruses trigger signaling pathways of innate immunity. In this issue, it isshown that the mitochondrial antiviral signaling protein is critical forintracellular detection signaling, but is dispensable for the activation ofinnate immunity via Toll-like receptors.
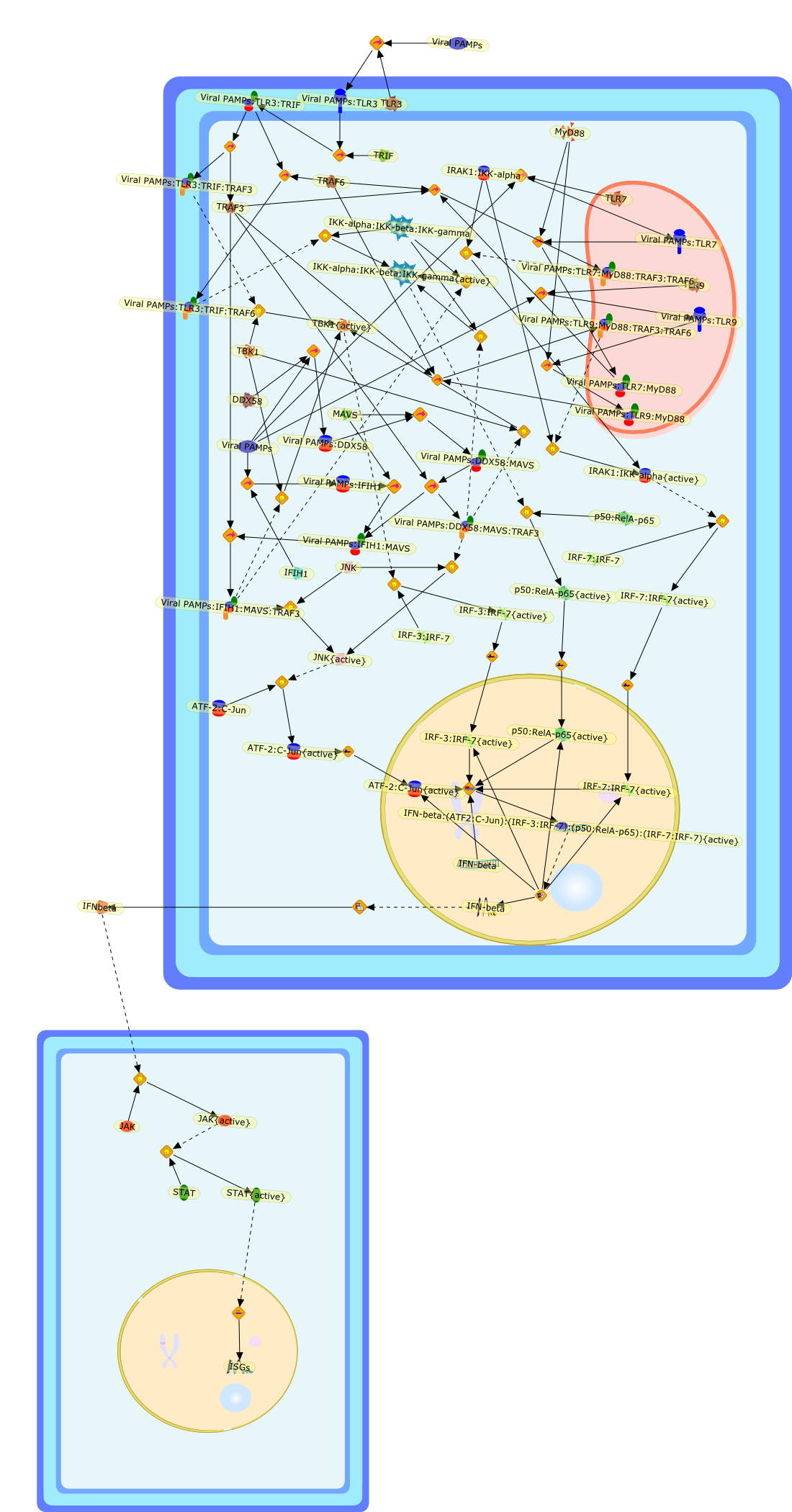{kind=link}
{kind=link}
ATF-2:C-Jun,ATF-2:C-Jun{active},DDX58,IFIH1,IFN-beta,IFN-beta:(ATF2:C-Jun):(IRF-3:IRF-7):(p50:RelA-p65):(IRF-7:IRF-7){active},IFNbeta,IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma{active},IRAK1:IKK-alpha,IRAK1:IKK-alpha{active},IRF-3:IRF-7,IRF-3:IRF-7{active},IRF-7:IRF-7,IRF-7:IRF-7{active},ISGs,JAK,JAK{active},JNK,JNK{active},MAVS,MyD88,STAT,STAT{active},TBK1,TBK1{active},TLR3,TLR7,TLR9,TRAF3,TRAF6,TRIF,Viral PAMPs,Viral PAMPs:DDX58,Viral PAMPs:DDX58:MAVS,Viral PAMPs:DDX58:MAVS:TRAF3,Viral PAMPs:IFIH1,Viral PAMPs:IFIH1:MAVS,Viral PAMPs:IFIH1:MAVS:TRAF3,Viral PAMPs:TLR3,Viral PAMPs:TLR3:TRIF,Viral PAMPs:TLR3:TRIF:TRAF3,Viral PAMPs:TLR3:TRIF:TRAF6,Viral PAMPs:TLR7,Viral PAMPs:TLR7:MyD88,Viral PAMPs:TLR7:MyD88:TRAF3:TRAF6,Viral PAMPs:TLR9,Viral PAMPs:TLR9:MyD88,Viral PAMPs:TLR9:MyD88:TRAF3:TRAF6,p50:RelA-p65,p50:RelA-p65{active}
The transcription factor NF-kappaB plays a major role in coordinating innate andadaptative immunity, cellular proliferation, apoptosis and development. Sincethe discovery in 1991 that NF-kappaB may be activated by H(2)O(2), severallaboratories have put a considerable effort into dissecting the molecularmechanisms underlying this activation. Whereas early studies revealed anatypical mechanism of activation, leading to IkappaBalpha Y42 phosphorylationindependently of IkappaB kinase (IKK), recent findings suggest that H(2)O(2)activates NF-kappaB mainly through the classical IKK-dependent pathway. Themolecular mechanisms leading to IKK activation are, however, cell-type specificand will be presented here. In this review, we also describe the effect of otherROS (HOCl and (1)O(2)) and reactive nitrogen species on NF-kappaB activation.Finally, we critically review the recent data highlighting the role of ROS inNF-kappaB activation by proinflammatory cytokines (TNF-alpha and IL-1beta) andlipopolysaccharide (LPS), two major components of innate immunity.
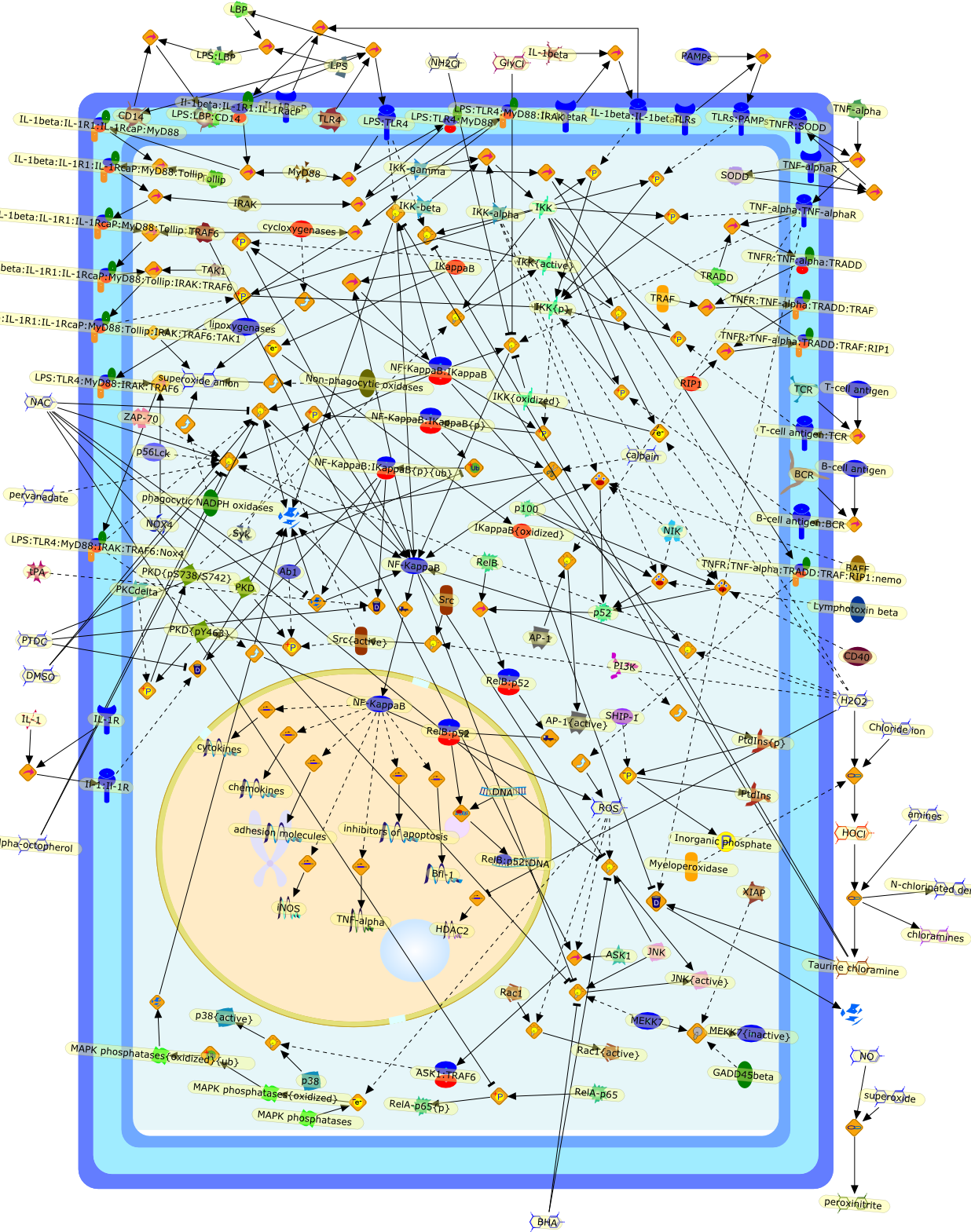{kind=link}
{kind=link}
degradants,AP-1,AP-1{active},ASK1,ASK1:TRAF6,Ab1,B-cell antigen,B-cell antigen:BCR,BAFF,BCR,BHA,Bfl-1,CD14,CD40,Chloride ion,DMSO,DNA,GADD45beta,GlyCl,H2O2,HDAC2,HOCl,IKK,IKK-alpha,IKK-beta,IKK-gamma,IKK{active},IKK{oxidized},IKK{p},IKappaB,IKappaB{oxidized},IL-1,IL-1R,IL-1RacP,IL-1beta,IL-1beta:IL-1R1:IL-1RcaP:MyD88,IL-1beta:IL-1R1:IL-1RcaP:MyD88:Tollip,IL-1beta:IL-1R1:IL-1RcaP:MyD88:Tollip:IRAK,IL-1beta:IL-1R1:IL-1RcaP:MyD88:Tollip:IRAK:TRAF6,IL-1beta:IL-1R1:IL-1RcaP:MyD88:Tollip:IRAK:TRAF6:TAK1,IL-1beta:IL-1betaR,IL-1betaR,IRAK,Il-1:Il-1R,Il-1beta:IL-1R1:IL-1RacP,Inorganic phosphate,JNK,JNK{active},LBP,LPS,LPS:LBP,LPS:LBP:CD14,LPS:TLR4,LPS:TLR4:MyD88,LPS:TLR4:MyD88:IRAK,LPS:TLR4:MyD88:IRAK:TRAF6,LPS:TLR4:MyD88:IRAK:TRAF6:Nox4,Lymphotoxin beta,MAPK phosphatases,MAPK phosphatases{oxidized},MAPK phosphatases{oxidized}{ub},MEKK7,MEKK7{inactive},MyD88,Myeloperoxidase,N-chlorinated derivatives,NAC,NF-KappaB,NF-KappaB:IKappaB,NF-KappaB:IKappaB{p},NF-KappaB:IKappaB{p}{ub},NH2Cl,NIK,NO,NOX4,Non-phagocytic oxidases,PAMPs,PI3K,PKCdelta,PKD,PKD{pS738/S742},PKD{pY463},PTDC,PtdIns,PtdIns{p},RIP1,ROS,Rac1,Rac1{active},RelA-p65,RelA-p65{p},RelB,RelB:p52,RelB:p52:DNA,SHIP-1,SODD,Src,Src{active},Syk,T-cell antigen,T-cell antigen:TCR,TAK1,TCR,TLR4,TLRs,TLRs:PAMPs,TNF-alpha,TNF-alpha:TNF-alphaR,TNF-alphaR,TNFR:SODD,TNFR:TNF-alpha:TRADD:TRAF,TNFR:TNF-alpha:TRADD:TRAF:RIP1,TNFR:TNF-alpha:TRADD:TRAF:RIP1:nemo,TNFR:TNF:alpha:TRADD,TRADD,TRAF,TRAF6,Taurine chloramine,Tollip,XIAP,ZAP-70,adhesion molecules,alpha-octopherol,amines,calpain,chemokines,chloramines,cycloxygenases,cytokines,degradants,iNOS,inhibitors of apoptosis,lipoxygenases,p100,p38,p38{active},p52,p56Lck,peroxinitrite,pervanadate,phagocytic NADPH oxidases,superoxide,superoxide anion,tPA
Innate immune responses to pathogens such as bacteria and viruses are triggeredby recognition of specific structures of invading pathogens calledpathogen-associated molecular patterns (PAMPs) by cellular pattern recognitionreceptors (PRRs) that are located at plasma membrane or inside cells.Stimulation of different PAMPs activates Toll-like receptor (TLR)-dependent and-independent signaling pathways that lead to activation of transcription factorsnuclear factor-kappaB (NF-kappaB), interferon regulatory factor 3/7 (IRF3/7)and/or activator protein-1 (AP-1), which collaborate to induce transcription ofa large number of downstream genes. This review focuses on the rapid progressthat has recently improved our understanding of the crosstalk among the pathwaysand the precise regulation of transcription of the downstream genes.
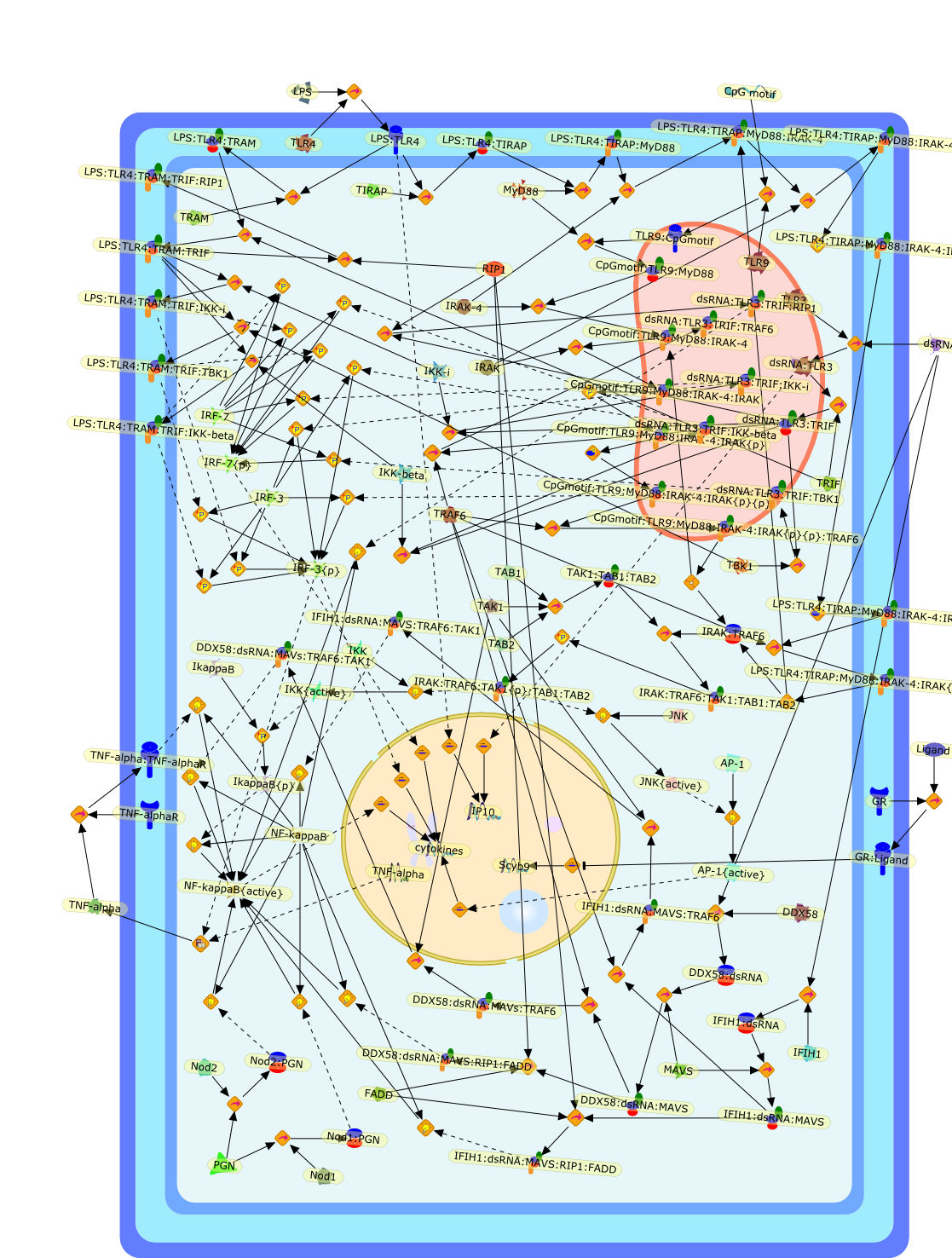{kind=link}
{kind=link}
AP-1,AP-1{active},CpG motif,CpGmotif:TLR9:MyD88,CpGmotif:TLR9:MyD88:IRAK-4,CpGmotif:TLR9:MyD88:IRAK-4:IRAK,CpGmotif:TLR9:MyD88:IRAK-4:IRAK{p},CpGmotif:TLR9:MyD88:IRAK-4:IRAK{p}{p},CpGmotif:TLR9:MyD88:IRAK-4:IRAK{p}{p}:TRAF6,DDX58,DDX58:dsRNA,DDX58:dsRNA:MAVS,DDX58:dsRNA:MAVS:RIP1:FADD,DDX58:dsRNA:MAVs:TRAF6,DDX58:dsRNA:MAVs:TRAF6:TAK1,FADD,GR,GR:Ligand,IFIH1,IFIH1:dsRNA,IFIH1:dsRNA:MAVS,IFIH1:dsRNA:MAVS:RIP1:FADD,IFIH1:dsRNA:MAVS:TRAF6,IFIH1:dsRNA:MAVS:TRAF6:TAK1,IKK,IKK-beta,IKK-i,IKK{active},IP10,IRAK,IRAK-4,IRAK:TRAF6,IRAK:TRAF6:TAK1:TAB1:TAB2,IRAK:TRAF6:TAK1{p}:TAB1:TAB2,IRF-3,IRF-3{p},IRF-7,IRF-7{p},IkappaB,IkappaB{p},JNK,JNK{active},LPS,LPS:TLR4,LPS:TLR4:TIRAP,LPS:TLR4:TIRAP:MyD88,LPS:TLR4:TIRAP:MyD88:IRAK-4,LPS:TLR4:TIRAP:MyD88:IRAK-4:IRAK,LPS:TLR4:TIRAP:MyD88:IRAK-4:IRAK{p},LPS:TLR4:TIRAP:MyD88:IRAK-4:IRAK{p}{p},LPS:TLR4:TIRAP:MyD88:IRAK-4:IRAK{p}{p}:TRAF6,LPS:TLR4:TRAM,LPS:TLR4:TRAM:TRIF,LPS:TLR4:TRAM:TRIF:IKK-beta,LPS:TLR4:TRAM:TRIF:IKK-i,LPS:TLR4:TRAM:TRIF:RIP1,LPS:TLR4:TRAM:TRIF:TBK1,Ligand,MAVS,MyD88,NF-kappaB,NF-kappaB{active},Nod1,Nod1:PGN,Nod2,Nod2:PGN,PGN,RIP1,Scyb9,TAB1,TAB2,TAK1,TAK1:TAB1:TAB2,TBK1,TIRAP,TLR3,TLR4,TLR9,TLR9:CpGmotif,TNF-alpha,TNF-alpha:TNF-alphaR,TNF-alphaR,TRAF6,TRAM,TRIF,cytokines,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:IKK-beta,dsRNA:TLR3:TRIF:IKK-i,dsRNA:TLR3:TRIF:RIP1,dsRNA:TLR3:TRIF:TBK1,dsRNA:TLR3:TRIF:TRAF6
Phagocytosis is a central event in the innate immune responses that aretriggered by the association between ligands on the surface of pathogens andreceptors on the membrane of phagocytes. Particularly, complement-mediatedphagocytosis is accomplished by specific recognition of bound complementcomponents by the corresponding complement receptors on the phagocytes. Theprotein-tyrosine kinase, Syk, plays a central role in Fcgamma receptor-mediatedphagocytosis in the adaptive immune system. From recent studies using amacrophage-like differentiated cell line and serum-treated zymosan, it was foundthat Syk also plays an essential role in complement-mediated phagocytosis ininnate immunity. Serum-treated zymosan particles promptly attached to the cellsand were subsequently engulfed via complement receptor3. During this process,Syk became tyrosine-phosphorylated and accumulated around the nascentphagosomes. The transfer of Syk-siRNA or dominant-negative Syk (DN-Syk) intomacrophages resulted in impaired engulfment of pathogen. Collectively, Syk isrequired for the engulfment of pathogen in complement-mediated phagocytosis.
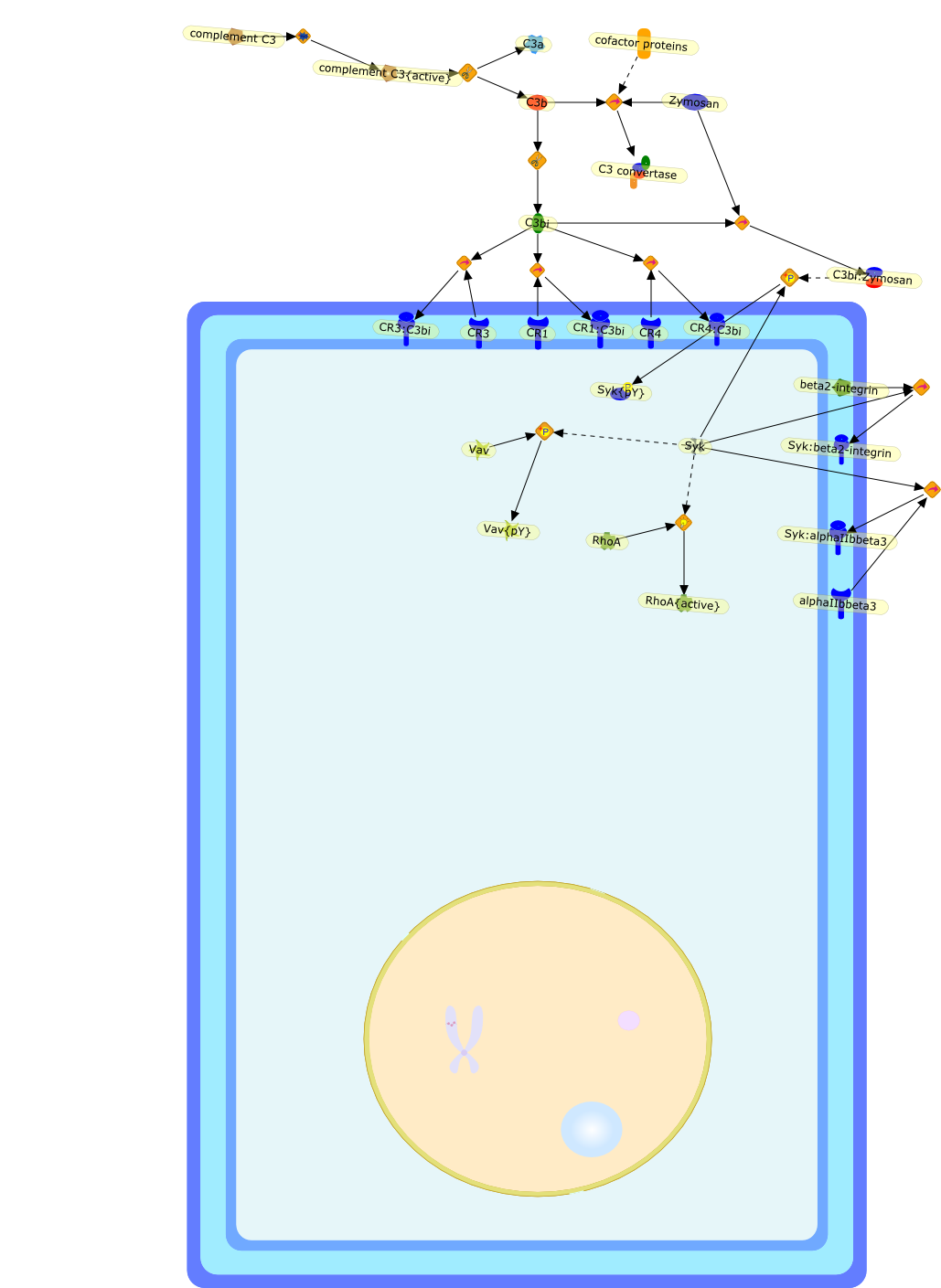{kind=link}
{kind=link}
C3 convertase,C3a,C3b,C3bi,C3bi:Zymosan,CR1,CR1:C3bi,CR3,CR3:C3bi,CR4,CR4:C3bi,RhoA,RhoA{active},Syk,Syk:alphaIIbbeta3,Syk:beta2-integrin,Syk{pY},Vav,Vav{pY},Zymosan,alphaIIbbeta3,beta2-integrin,cofactor proteins,complement C3,complement C3{active}
The early detection by the host of invading microorganisms, including viruses,depends on a limited number of specific receptors that recognizepathogen-associated molecular patterns (PAMPs). A few of these PAMPs, includingssRNA and dsRNA, are recognized by Toll-like receptors (TLR)-7/8 and TLR3,respectively. Activation of an antiviral TLR-dependent signaling cascade leadsto the activation of the key transcription factors IRF and NF-kappaB, whichpromote antiviral responses through induction of specific genes. Recently, asecond system has been described, which relies on the cytoplasmic recognition ofdsRNA by RNA helicases such as RIG-I. In this review, we discuss the mechanisticaspects of these important arms of the host innate response to dsRNA and a fewviral strategies utilized to counteract them.
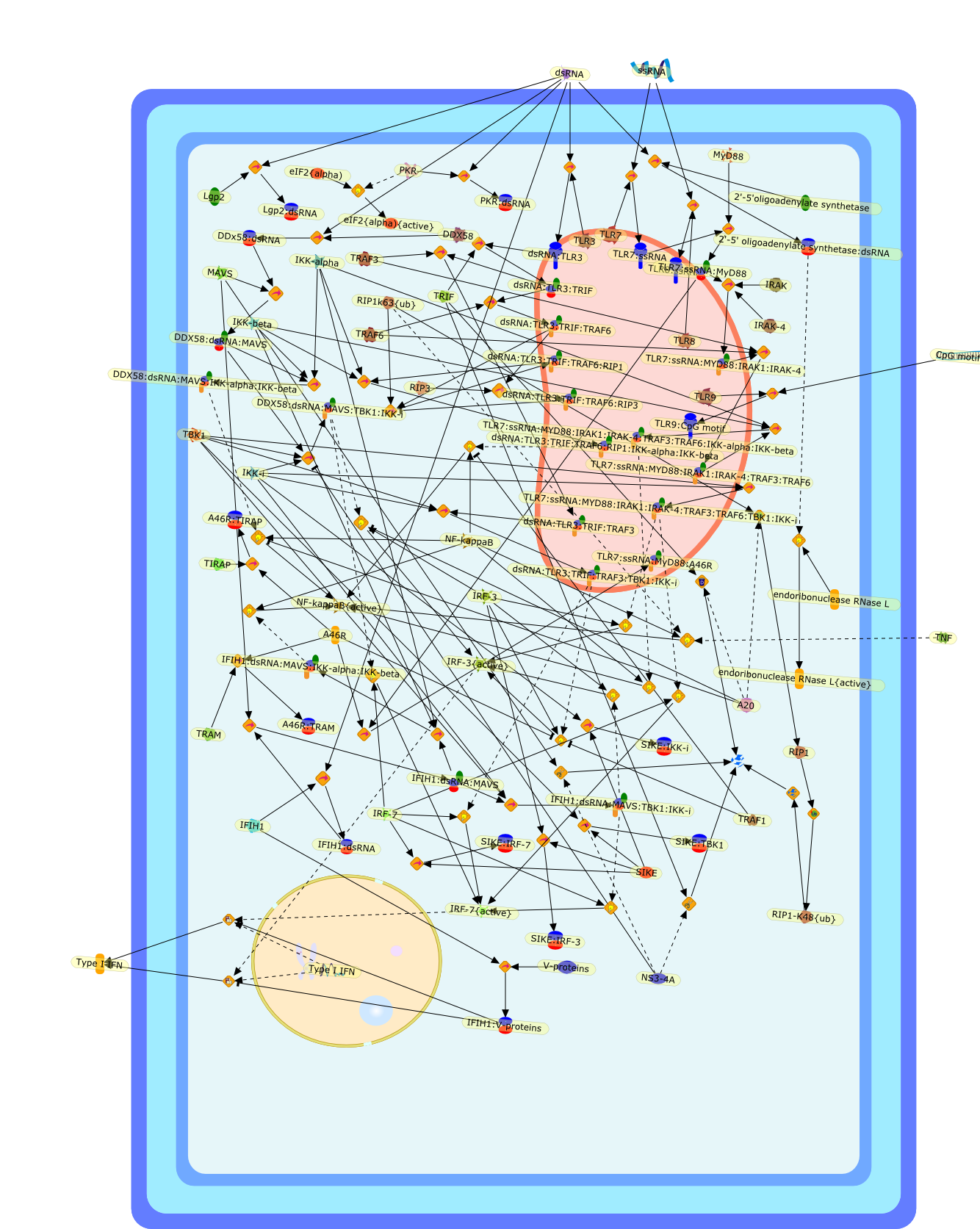{kind=link}
{kind=link}
2'-5' oligoadenylate synthetase:dsRNA,2'-5'oligoadenylate synthetase,A20,A46R,A46R:TIRAP,A46R:TRAM,CpG motif,DDX58,DDX58:dsRNA:MAVS,DDX58:dsRNA:MAVS:IKK-alpha:IKK-beta,DDX58:dsRNA:MAVS:TBK1:IKK-i,DDx58:dsRNA,IFIH1,IFIH1:V-proteins,IFIH1:dsRNA,IFIH1:dsRNA:MAVS,IFIH1:dsRNA:MAVS:IKK-alpha:IKK-beta,IFIH1:dsRNA:MAVS:TBK1:IKK-i,IKK-alpha,IKK-beta,IKK-i,IRAK,IRAK-4,IRF-3,IRF-3{active},IRF-7,IRF-7{active},Lgp2,Lgp2:dsRNA,MAVS,MyD88,NF-kappaB,NF-kappaB{active},NS3-4A,PKR,PKR:dsRNA,RIP1,RIP1-K48{ub},RIP1k63{ub},RIP3,SIKE,SIKE:IKK-i,SIKE:IRF-3,SIKE:IRF-7,SIKE:TBK1,TBK1,TIRAP,TLR3,TLR7,TLR7:ssRNA,TLR7:ssRNA:MYD88:IRAK1:IRAK-4,TLR7:ssRNA:MYD88:IRAK1:IRAK-4:TRAF3:TRAF6,TLR7:ssRNA:MYD88:IRAK1:IRAK-4:TRAF3:TRAF6:IKK-alpha:IKK-beta,TLR7:ssRNA:MYD88:IRAK1:IRAK-4:TRAF3:TRAF6:TBK1:IKK-i,TLR7:ssRNA:MyD88,TLR7:ssRNA:MyD88:A46R,TLR8,TLR8:ssRNA,TLR9,TLR9:CpG motif,TNF,TRAF1,TRAF3,TRAF6,TRAM,TRIF,Type I IFN,V-proteins,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:TRAF3,dsRNA:TLR3:TRIF:TRAF3:TBK1:IKK-i,dsRNA:TLR3:TRIF:TRAF6,dsRNA:TLR3:TRIF:TRAF6:RIP1,dsRNA:TLR3:TRIF:TRAF6:RIP1:IKK-alpha:IKK-beta,dsRNA:TLR3:TRIF:TRAF6:RIP3,eIF2{alpha),eIF2{alpha){active},endoribonuclease RNase L,endoribonuclease RNase L{active},protein remnants,ssRNA
TLRs are crucial sensors of microbial infection. Maintaining structuralintegrity of TLR signaling components is essential for subsequent immunologicalprotection. Alterations to the structure of these signaling molecules are oftenassociated with profound clinical outcomes and susceptibility to variousinfectious diseases. These changes in structure are sometimes the result of asingle nucleotide polymorphism (SNP). Numerous SNPs have been found incomponents of the TLR signaling pathway. Recently, the medical consequences andeffects on TLR signaling of several of these SNPs have been elucidated. Inaddition, there have been numerous structures solved that are important to ourunderstanding of the TLR signaling pathway at the molecular level. The scope ofthis review is to tie together current structural, biochemical, and geneticinformation of TLR signaling.
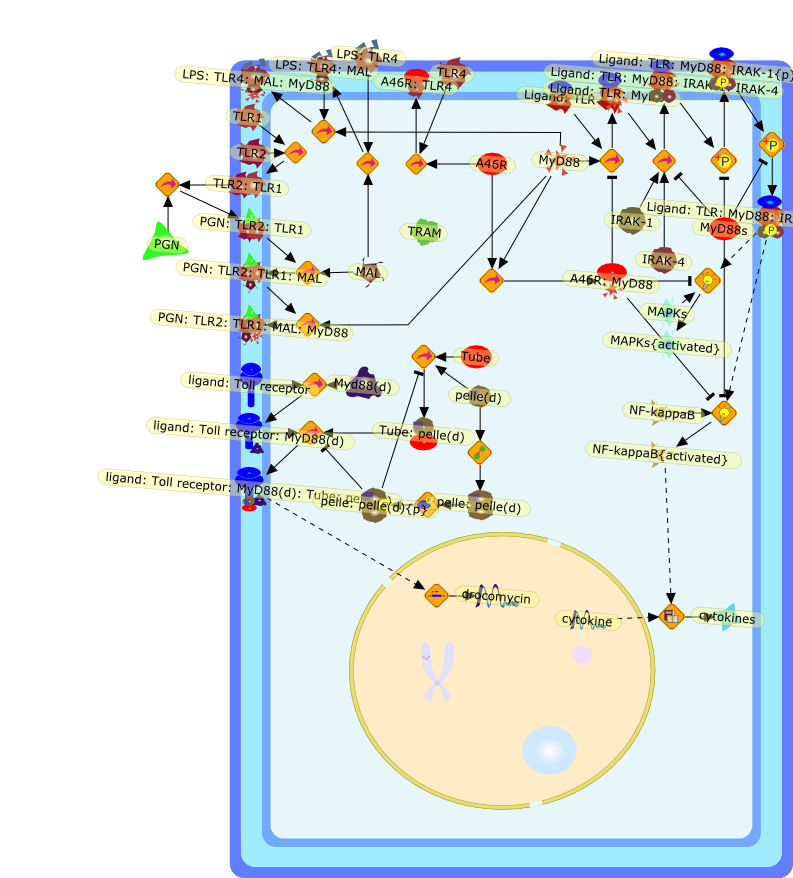{kind=link}
{kind=link}
A46R,A46R: MyD88,A46R: TLR4,IRAK-1,IRAK-4,LPS: TLR4,LPS: TLR4: MAL,LPS: TLR4: MAL: MyD88,Ligand: TLR,Ligand: TLR: MyD88,Ligand: TLR: MyD88: IRAK-1: IRAK-4,Ligand: TLR: MyD88: IRAK-1{p}: IRAK-4,MAL,MAPKs,MAPKs{activated},MyD88,MyD88s,Myd88(d),NF-kappaB,NF-kappaB{activated},PGN,PGN: TLR2: TLR1,PGN: TLR2: TLR1: MAL,PGN: TLR2: TLR1: MAL: MyD88,TLR1,TLR2,TLR2: TLR1,TLR4,TRAM,Tube,Tube: pelle(d),cytokine,cytokines,drocomycin,ligand: Toll receptor,ligand: Toll receptor: MyD88(d),ligand: Toll receptor: MyD88(d): Tube: pelle(d),pelle(d),pelle: pelle(d),pelle: pelle(d){p}
Type I IFN (IFN-alpha/beta) have important biological functions ranging fromimmune cell development and activation, to tumor cell killing and mostimportantly inhibition of virus replication. Following viral infection oractivation of Toll-like receptors (TLRs) via distinct ligands, IFN-alpha/betaare produced. Two members of the interferon regulatory factor (IRF) family -IRF-3 and IRF-7 - are the major modulators of IFN gene expression. Activation ofIRF-3 and IRF-7 by TBK1/IKKvarepsilon mediated phosphorylation promotes IFN geneexpression and potentiates the production of IFN responsive genes important tothe development of an effective antiviral immune response. IFN treatment canaugment anti-tumor properties and they are potentially key players in cancertherapy. For example, adoptive transfer of IFN-gamma-activated macrophages canmediate tumor cell killing via direct cell-cell contact, as well as release ofsoluble cytotoxic pro-inflammatory molecules. A recent study investigatedwhether IRF-3 and IRF-7 could mediate the acquisition of new anti-tumor effectorfunctions in macrophages. Adenovirus mediated transduction of the active form ofIRF-7 into primary macrophages resulted in the production of type I IFN,upregulation of target genes including TRAIL and increased tumoricidal activityof macrophages; in contrast, the active form of IRF-3 led to induction of celldeath. These studies indicate that IRF-7 transduced macrophages may be anattractive candidate for in vivo adoptive therapy of cancer.
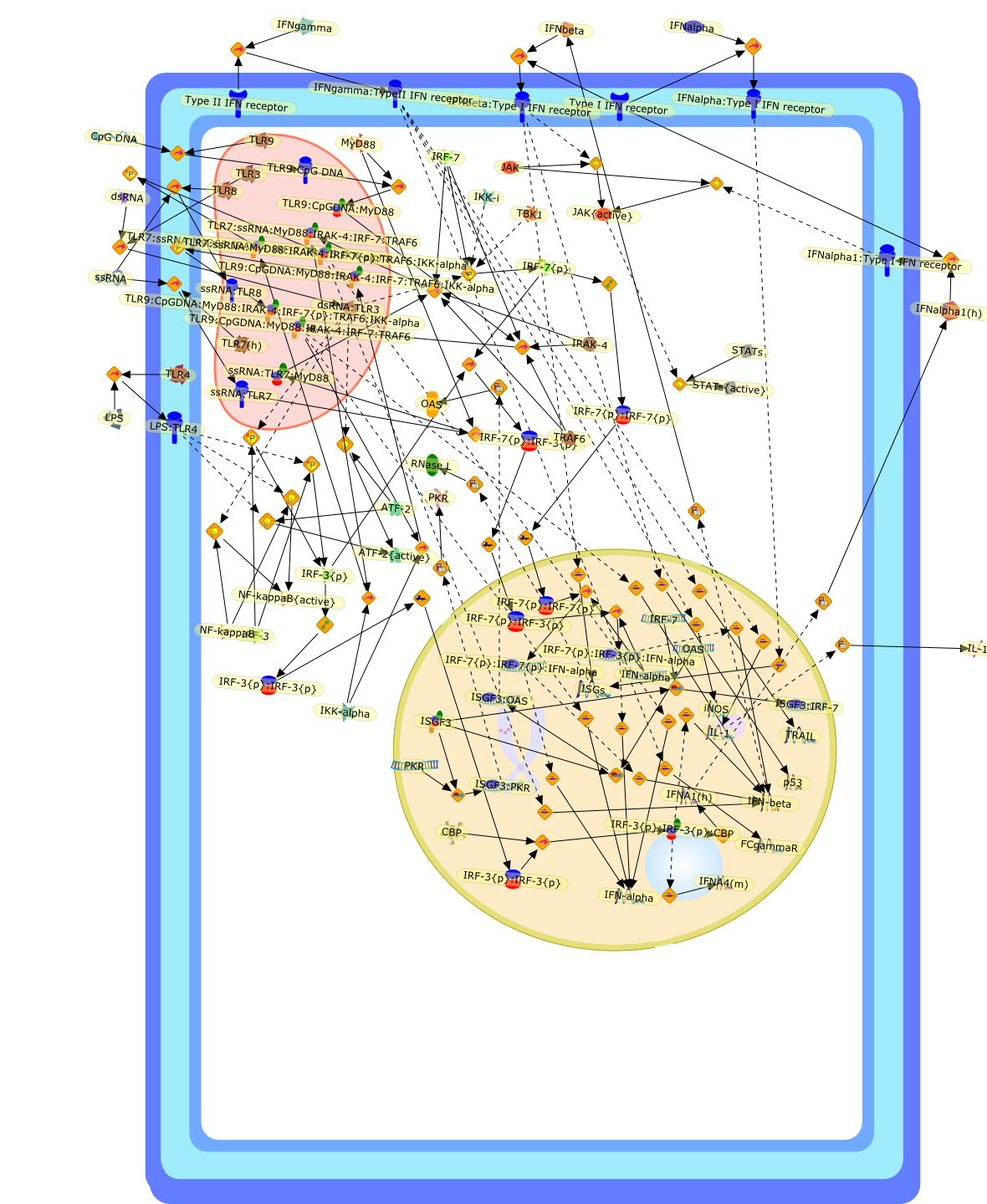{kind=link}
{kind=link}
ATF-2,ATF-2{active},CBP,CpG DNA,FCgammaR,IFN-alpha,IFN-beta,IFNA1(h),IFNA4(m),IFNalpha,IFNalpha1(h),IFNalpha1:Type I IFN receptor,IFNalpha:Type I IFN receptor,IFNbeta,IFNbeta:Type I IFN receptor,IFNgamma,IFNgamma:TypeII IFN receptor,IKK-alpha,IKK-i,IL-1,IRAK-4,IRF-3,IRF-3{p},IRF-3{p}:IRF-3{p},IRF-3{p}:IRF-3{p}:CBP,IRF-7,IRF-7{p},IRF-7{p}:IRF-3{p},IRF-7{p}:IRF-3{p}:IFN-alpha,IRF-7{p}:IRF-7{p},IRF-7{p}:IRF-7{p}:IFN-alpha,ISGF3,ISGF3:IRF-7,ISGF3:OAS,ISGF3:PKR,ISGs,JAK,JAK{active},LPS,LPS:TLR4,MyD88,NF-kappaB,NF-kappaB{active},OAS,PKR,RNase L,STATs,STATs{active},TBK1,TLR3,TLR4,TLR7(h),TLR7:ssRNA:MyD88:IRAK-4:IRF-7:TRAF6,TLR7:ssRNA:MyD88:IRAK-4:IRF-7:TRAF6:IKK-alpha,TLR7:ssRNA:MyD88:IRAK-4:IRF-7{p}:TRAF6:IKK-alpha,TLR8,TLR9,TLR9:CpG DNA,TLR9:CpGDNA:MyD88,TLR9:CpGDNA:MyD88:IRAK-4:IRF-7:TRAF6,TLR9:CpGDNA:MyD88:IRAK-4:IRF-7:TRAF6:IKK-alpha,TLR9:CpGDNA:MyD88:IRAK-4:IRF-7{p}:TRAF6:IKK-alpha,TRAF6,TRAIL,Type I IFN receptor,Type II IFN receptor,dsRNA,dsRNA:TLR3,iNOS,p53,ssRNA,ssRNA:TLR7,ssRNA:TLR7:MyD88,ssRNA:TLR8
LPS tolerance has been the focus of extensive scientific and clinical researchover the last several decades in an attempt to elucidate the sequence of changesthat occur at a molecular level in tolerized cells. Tolerance to components ofgram-positive bacterial cell walls such as bacterial lipoprotein andlipoteichoic acid is a much lesser studied, although equally important,phenomenon. This review will focus on cellular reprogramming by gram-positivebacterial components and examines the alterations in cell surface receptorexpression, changes in intracellular signaling, gene expression and cytokineproduction, and the phenomenon of cross-tolerance.
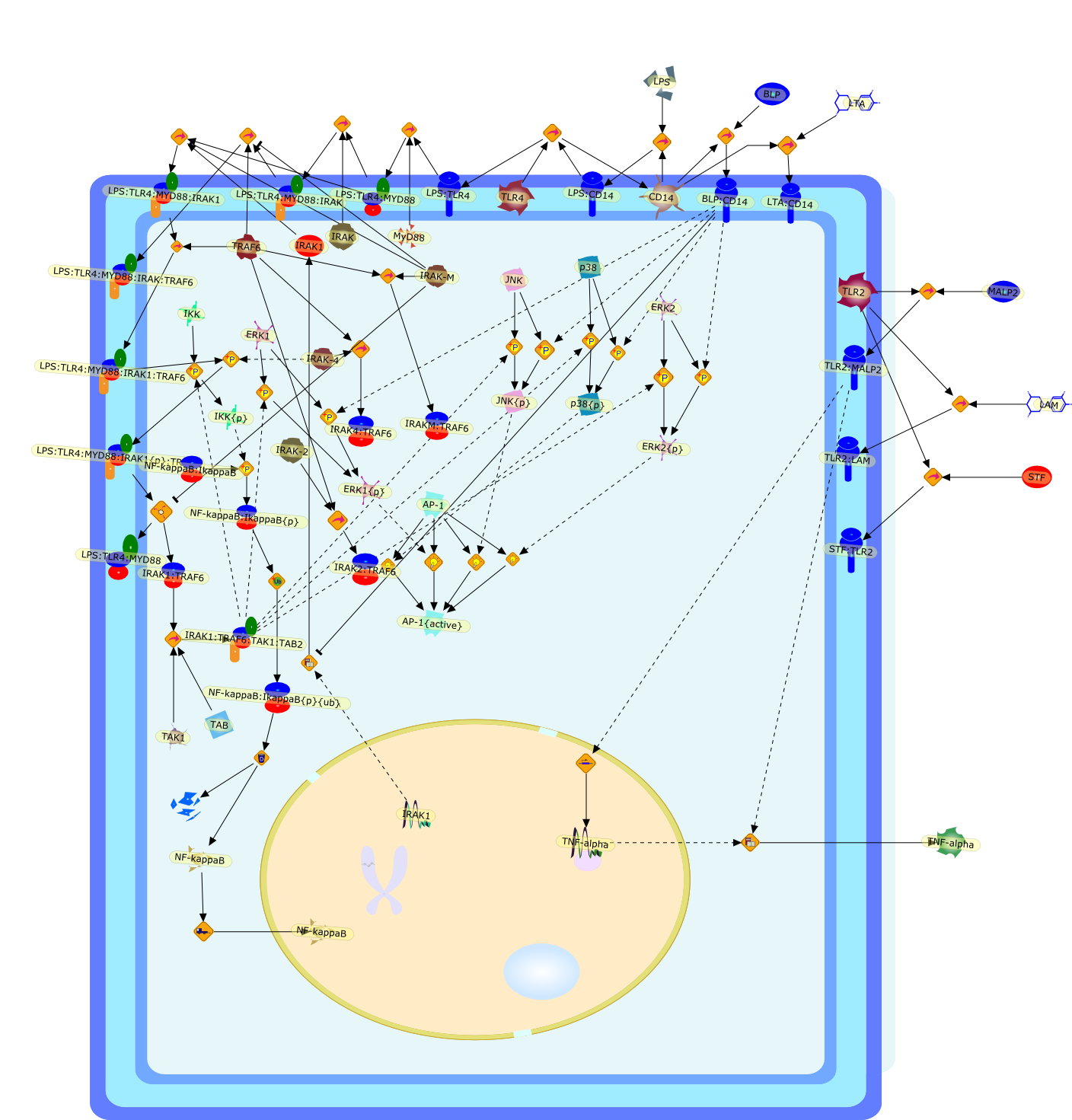{kind=link}
{kind=link}
AP-1,AP-1{active},BLP,BLP:CD14,CD14,ERK1,ERK1{p},ERK2,ERK2{p},IKK,IKK{p},IRAK,IRAK-2,IRAK-4,IRAK-M,IRAK1,IRAK1:TRAF6,IRAK1:TRAF6:TAK1:TAB2,IRAK2:TRAF6,IRAK4:TRAF6,IRAKM:TRAF6,JNK,JNK{p},LAM,LPS,LPS:CD14,LPS:TLR4,LPS:TLR4:MYD88,LPS:TLR4:MYD88:IRAK,LPS:TLR4:MYD88:IRAK1,LPS:TLR4:MYD88:IRAK1:TRAF6,LPS:TLR4:MYD88:IRAK1{p}:TRAF6,LPS:TLR4:MYD88:IRAK:TRAF6,LTA,LTA:CD14,MALP2,MyD88,NF-kappaB,NF-kappaB:IkappaB,NF-kappaB:IkappaB{p},NF-kappaB:IkappaB{p}{ub},STF,STF:TLR2,TAB,TAK1,TLR2,TLR2:LAM,TLR2:MALP2,TLR4,TNF-alpha,TRAF6,p38,p38{p},protein remnants
Macrophages are major effector cells of the innate immune system, andappropriate regulation of macrophage function requires the integration ofmultiple signalling inputs derived from the recognition of host factors (e.g.interferon-gamma/IFNgamma) and pathogen products (e.g. toll-like receptor/TLRagonists). The profound effects of IFNgamma pre-treatment ("priming") onTLR-induced macrophage activation have long been recognised, but many of themechanisms underlying the priming phenotype have only recently been identified.This review summarises the known mechanisms of integration between the IFNgammaand TLR signalling pathways. Synergy occurs at multiple levels, ranging fromsignal recognition to convergence of signals at the promoters of target genes.In particular, the cross-talk between the IFNgamma, and LPS and CpG DNAsignalling pathways is discussed.
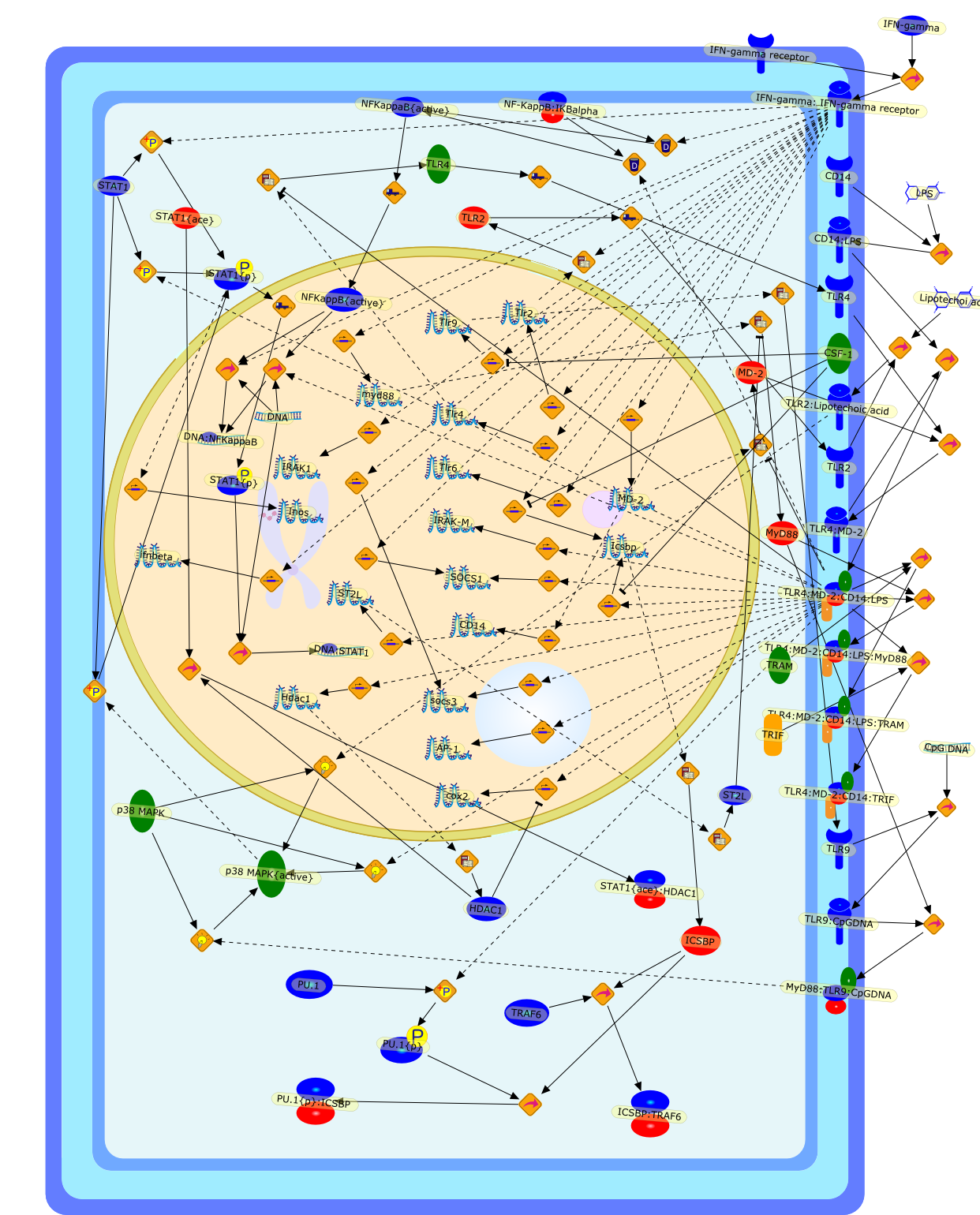{kind=link}
{kind=link}
AP-1,CD14,CD14:LPS,CSF-1,CpG DNA,DNA,DNA:NFKappaB,DNA:STAT1,HDAC1,Hdac1,ICSBP,ICSBP:TRAF6,IFN-gamma,IFN-gamma receptor,IFN-gamma: IFN-gamma receptor,IRAK-M,IRAK1,Icsbp,Inos,LPS,Lipotechoi acid,MD-2,MyD88,MyD88:TLR9:CpGDNA,NF-KappB:IKBalpha,NFKappB{active},NFKappaB{active},PU.1,PU.1{p},PU.1{p}:ICSBP,SOCS1,ST2L,STAT1,STAT1{ace},STAT1{ace}:HDAC1,STAT1{p},TLR2,TLR2:Lipotechoic acid,TLR4,TLR4:MD-2,TLR4:MD-2:CD14:LPS,TLR4:MD-2:CD14:LPS:MyD88,TLR4:MD-2:CD14:LPS:TRAM,TLR4:MD-2:CD14:TRIF,TLR9,TLR9:CpGDNA,TRAF6,TRAM,TRIF,Tlr2,Tlr4,Tlr6,Tlr9,cox2,ifnbeta,myd88,p38 MAPK,p38 MAPK{active},socs3
Toll-like receptors (TLRs) are key components of the innate immune system,functioning as pattern recognition receptors that recognise a wide range ofmicrobial pathogens. TLRs represent a primary line of defence against invadingpathogens in mammals, plants and insects. Recognition of microbial components byTLRs triggers a cascade of cellular signals that culminates in the activation ofNFkappaB which leads to inflammatory gene expression and clearance of theinfectious agent. The history of NFkappaB began with the TLR4 ligandlipopolysaccharide (LPS), a component of the cell wall of Gram-negativebacteria, since this was the stimulus first used to activate NFkappaB inpre-B-cells. However, since those early days it has been a circuitous route,made possible by drawing on information provided by many different fields, thathas led us not only to the discovery of TLRs but also to an understanding of thecomplex pathways that lead from TLR ligation to NFkappaB activation. In thisreview we will summarize the current knowledge of TLR-mediated NFkappaBactivation, and also the recent discoveries that subtle differences in kappaBbinding sequences and NFkappaB dimer formation result in specific geneexpression profiles.
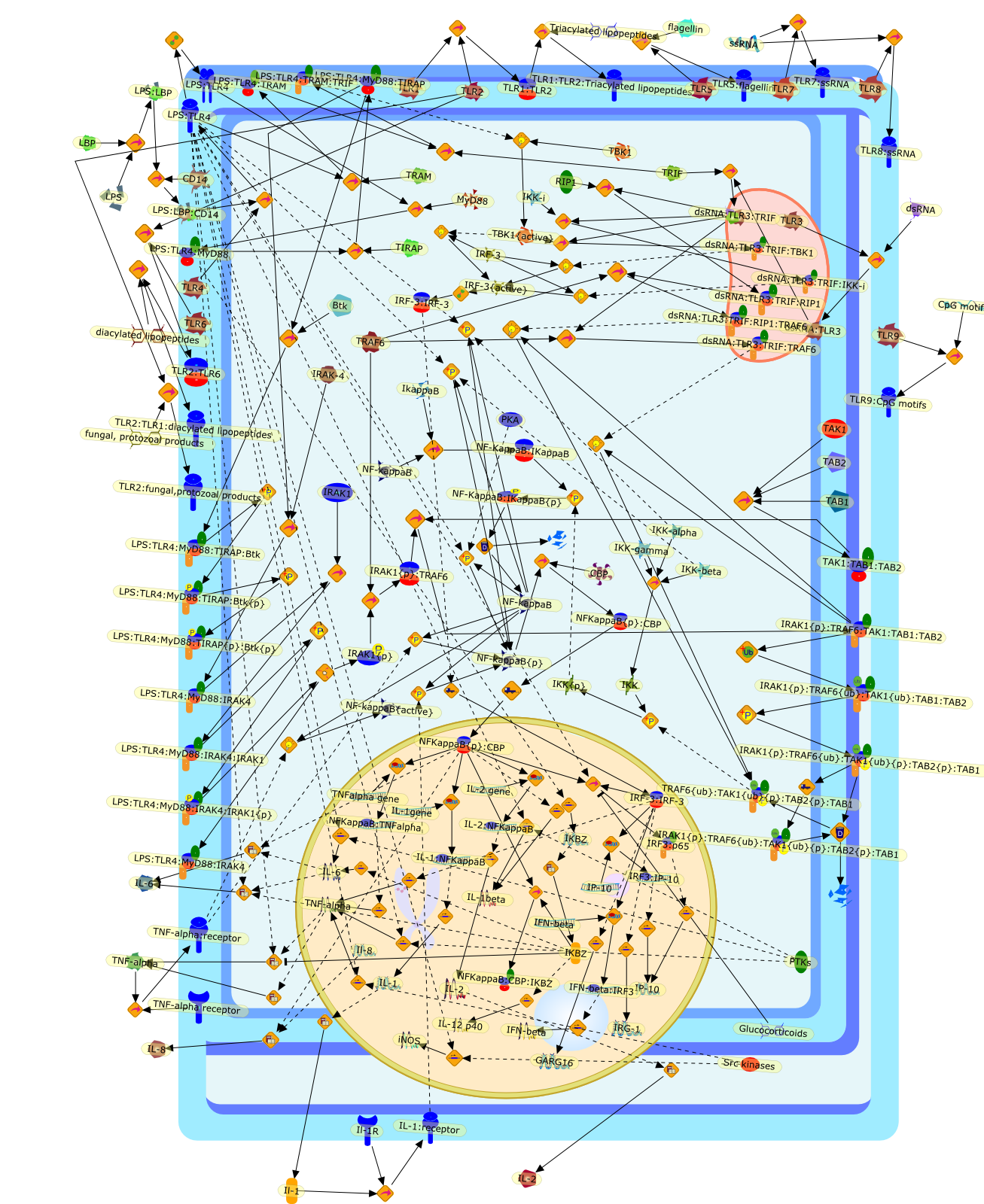{kind=link}
{kind=link}
Btk,CBP,CD14,CpG motifs,GARG16,Glucocorticoids,IFN-beta,IFN-beta:IRF3,IKBZ,IKK,IKK-alpha,IKK-beta,IKK-gamma,IKK-i,IKK{p},IL-1,IL-12 p40,IL-1:NFKappaB,IL-1:receptor,IL-1beta,IL-1gene,IL-2,IL-2 gene,IL-2:NFKappaB,IL-6,IL-8,IP-10,IRAK-4,IRAK1,IRAK1{p},IRAK1{p}:TRAF6,IRAK1{p}:TRAF6:TAK1:TAB1:TAB2,IRAK1{p}:TRAF6{ub}:TAK1{ub}:TAB1:TAB2,IRAK1{p}:TRAF6{ub}:TAK1{ub}{p}:TAB2{p}:TAB1,IRF-3,IRF-3:IRF-3,IRF-3{active},IRF3:IP-10,IRF3:p65,IRG-1,IkappaB,Il-1,Il-1R,Il-8,LBP,LPS,LPS:LBP,LPS:LBP:CD14,LPS:TLR4,LPS:TLR4:MyD88,LPS:TLR4:MyD88:IRAK4,LPS:TLR4:MyD88:IRAK4:IRAK1,LPS:TLR4:MyD88:IRAK4:IRAK1{p},LPS:TLR4:MyD88:TIRAP,LPS:TLR4:MyD88:TIRAP:Btk,LPS:TLR4:MyD88:TIRAP:Btk{p},LPS:TLR4:MyD88:TIRAP{p}:Btk{p},LPS:TLR4:TRAM,LPS:TLR4:TRAM:TRIF,MyD88,NF-KappaB:IKappaB,NF-KappaB:IKappaB{p},NF-kappaB,NF-kappaB ,NF-kappaB{active},NF-kappaB{p},NFKappaB:CBP:IKBZ,NFKappaB:TNFalpha,NFKappaB{p}:CBP,PKA,PTKs,RIP1,Src kinases,TAB1,TAB2,TAK1,TAK1:TAB1:TAB2,TBK1,TBK1{active},TIRAP,TLR1,TLR1:TLR2,TLR1:TLR2:Triacylated lipopeptides,TLR2,TLR2:TLR1:diacylated lipopeptides,TLR2:TLR6,TLR2:fungal,protozoal products,TLR3,TLR4,TLR5,TLR5:flagellin,TLR6,TLR7,TLR7:ssRNA,TLR8,TLR8:ssRNA,TLR9,TLR9:CpG motifs,TNF-alpha,TNF-alpha receptor,TNF-alpha:receptor,TNFalpha gene,TRAF6,TRAF6{ub}:TAK1{ub}{p}:TAB2{p}:TAB1,TRAM,TRIF,Triacylated lipopeptides,diacylated lipopeptides,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:IKK-i,dsRNA:TLR3:TRIF:RIP1,dsRNA:TLR3:TRIF:RIP1:TRAF6,dsRNA:TLR3:TRIF:TBK1,dsRNA:TLR3:TRIF:TRAF6,flagellin,fungal, protozoal products,iNOS,proteosome remnants,ssRNA
Exposure to pathogens induces antigen-presenting cells (APC) such as macrophagesand dendritic cells (DC) to produce various endogenous mediators, includingarachidonic acid (AA)-derived eicosanoids, cytokines, and nitric oxide (NO).Many secreted products of activated APC can act by themselves in an autocrinemanner and modulate their function. Moreover, the cross-interaction betweenendogenous bioactive molecules regulates the function of professional APC withimportant consequences for their ability to activate and sustain immune andinflammatory responses, and to regulate immune homeostasis. Although neglectedfor many years when compared to their role in cardiovascular homeostasis, cancerand inflammation, the importance of eicosanoids in immunology is becoming moredefined. The role of prostaglandin (PG) E2 (PGE2), one of the best known andmost well studied eicosanoids, is of particular interest. It modulates theactivities of professional DC by acting on their differentiation, maturation andtheir ability to secrete cytokines. Uniquely among haematopoietic cytokines,interleukin-10 (IL-10) is a pleiotropic molecule that displays bothimmunostimulatory and immunoregulatory activities. IL-10 has attached muchattention because of its anti-inflammatory properties. It modulates expressionof cytokines, soluble mediators and cell surface molecules by cells of myeloidorigin, particularly macrophages and DC. We previously reported that PGE2 is apotent inducer of IL-10 in bone marrow-derived DC (BM-DC), and PGE2-inducedIL-10 is a key regulator of the BM-DC pro-inflammatory phenotype. BM-DC may beconsidered as an important model to study complex interactions betweenendogenous mediators, and autocrine IL-10 plays a pivotal role in thecrossregulation of AA-derived lipid mediators, cytokines, and NO, with criticaleffects on immune and inflammatory responses.
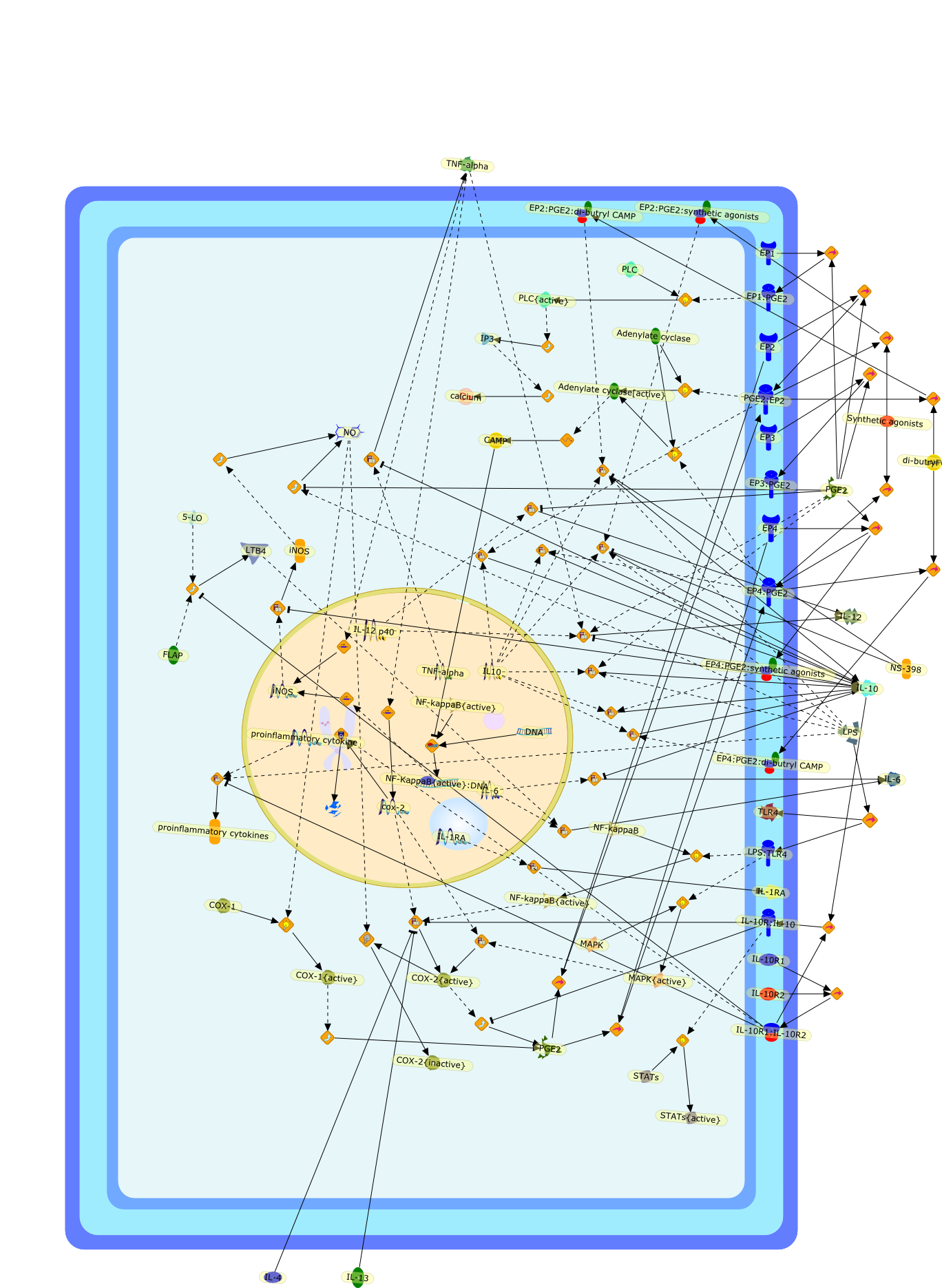{kind=link}
{kind=link}
5-LO,Adenylate cyclase,Adenylate cyclase[active},CAMP,COX-1,COX-1{active},COX-2{active},COX-2{inactive},DNA,EP1,EP1:PGE2,EP2,EP2:PGE2:di-butryl CAMP,EP2:PGE2:synthetic agonists,EP3,EP3:PGE2,EP4,EP4:PGE2,EP4:PGE2:di-butryl CAMP,EP4:PGE2:synthetic agonists,FLAP,IL-10,IL-10R1,IL-10R1:IL-10R2,IL-10R2,IL-10R:IL-10,IL-12,IL-12 p40,IL-13,IL-1RA,IL-4,IL-6,IL10,IP3,LPS,LPS:TLR4,LTB4,MAPK,MAPK{active},NF-KappaB{active}:DNA,NF-kappaB,NF-kappaB{active},NO,NS-398,PGE2,PGE2:EP2,PLC,PLC{active},STATs,STATs{active},Synthetic agonists,TLR4,TNF-alpha,calcium,cox-2,degradants,di-butryl CAMP,iNOS,proinflammatory cytokine,proinflammatory cytokines
Macrophages play a critical role in mediating the host response to biomaterials,perhaps most notably by guiding the host inflammatory response through therelease of inflammatory molecules such as the cytokine interleukin-1 (IL-1). Theextent of the macrophage response following interaction with the biomaterialsurface contributes greatly to device efficacy, yet the molecular mechanisms ofthis interaction are still unclear. The extracellular matrix (ECM) proteinfibronectin (FN) is recognized by macrophages and frequently used in biomaterialmodification to elicit greater cellular adhesion and tissue integration.Macrophage interaction with FN and other ECM molecules on the biomaterialsurface has been shown to induce a variety of inflammatory responses, thus bothFN and IL-1 can be utilized as model molecules to better understand themechanisms of material-mediated macrophage responses. This literature reviewpresents a comprehensive survey of past and current research on the interrelatedrole of IL-1, FN, and FN-derivatives in determining biomaterial-modulatedmacrophage function.
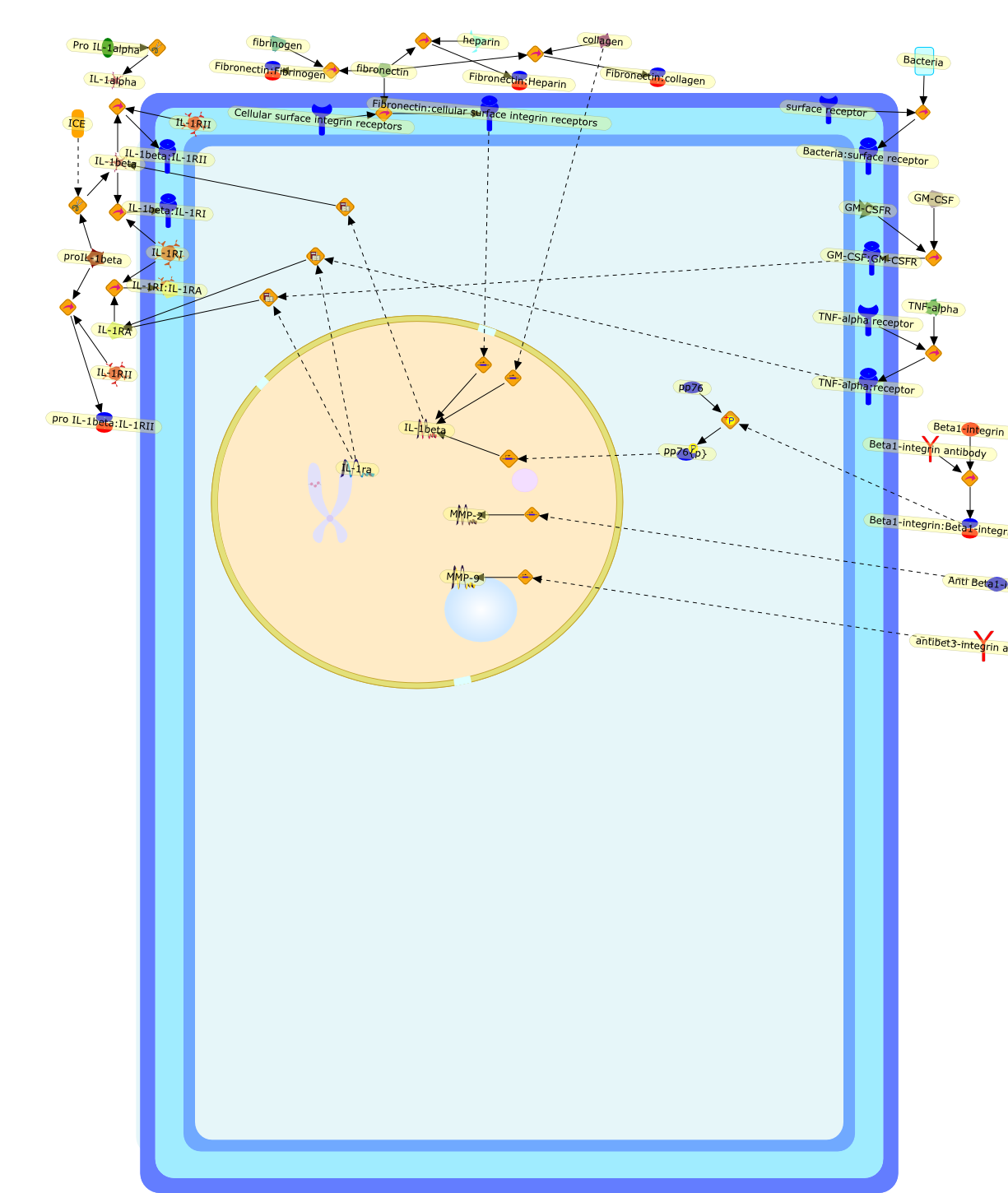{kind=link}
{kind=link}
Anti Beta1-integrin,Bacteria,Bacteria:surface receptor,Beta1-integrin,Beta1-integrin antibody,Beta1-integrin:Beta1-integrin antibody,Cellular surface integrin receptors,Fibronectin:Fibrinogen,Fibronectin:Heparin,Fibronectin:cellular surface integrin receptors,Fibronectin:collagen,GM-CSF,GM-CSF:GM-CSFR,GM-CSFR,ICE,IL-1RA,IL-1RI,IL-1RI:IL-1RA,IL-1RII,IL-1alpha,IL-1beta,IL-1beta:IL-1RI,IL-1beta:IL-1RII,IL-1ra,MMP-2,MMP-9,Pro IL-1alpha,TNF-alpha,TNF-alpha receptor,TNF-alpha:receptor,antibet3-integrin antibody,collagen,fibrinogen,fibronectin,heparin,pp76,pp76{p},pro IL-1beta:IL-1RII,proIL-1beta,surface receptor
Nearly half a century has passed since the first published description ofinterferons (IFNs). This commentary introduces the four accompanying reviewarticles on type I IFN research and attempts to relate how the field of IFNresearch has been changing during its history.
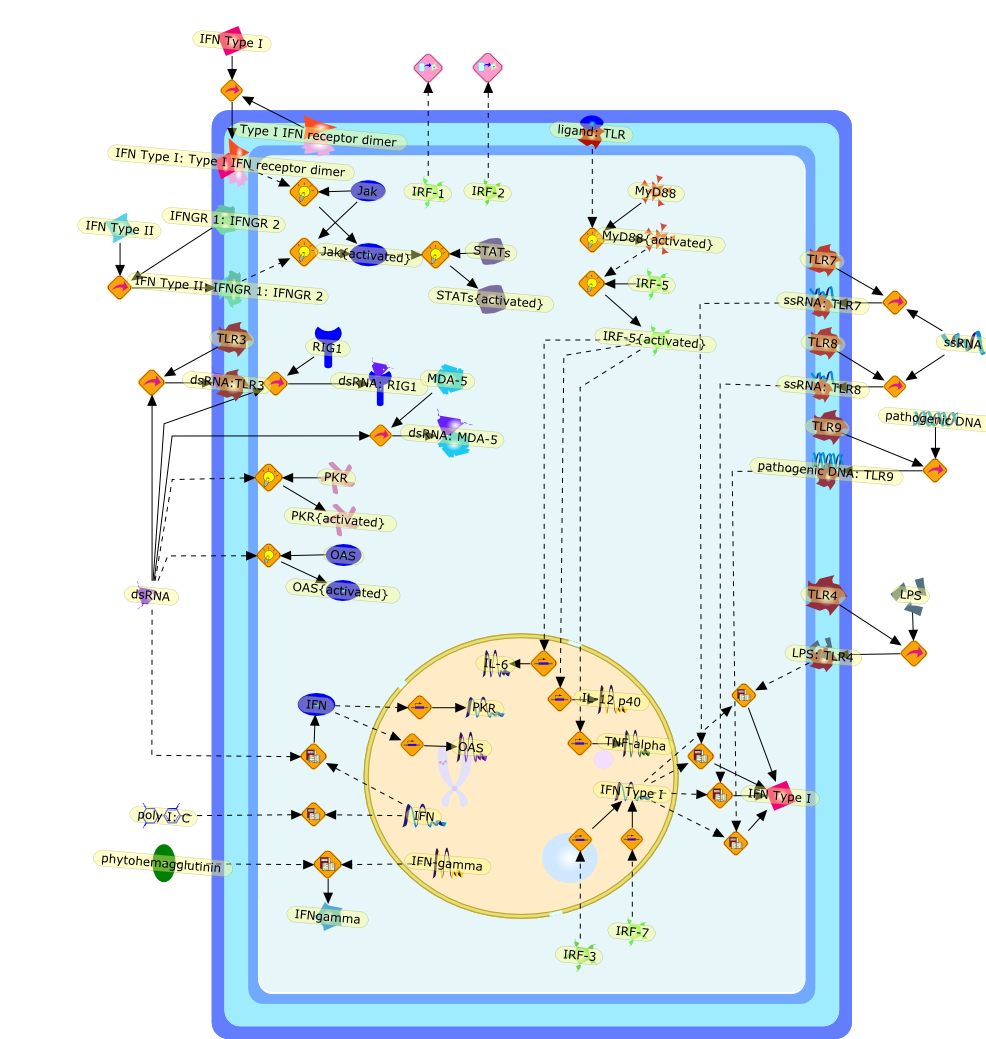{kind=link}
{kind=link}
IFN,IFN Type I,IFN Type I: Type I IFN receptor dimer,IFN Type II,IFN Type II: IFNGR 1: IFNGR 2,IFN-gamma,IFNGR 1: IFNGR 2,IFNgamma,IL-12 p40,IL-6,IRF-1,IRF-2,IRF-3,IRF-5,IRF-5{activated},IRF-7,Jak,Jak{activated},LPS,LPS: TLR4,MDA-5,MyD88,MyD88{activated},OAS,OAS{activated},PKR,PKR{activated},RIG1,STATs,STATs{activated},TLR3,TLR4,TLR7,TLR8,TLR9,TNF-alpha,Type I IFN receptor dimer,dsRNA,dsRNA: MDA-5,dsRNA: RIG1 ,dsRNA:TLR3,ligand: TLR,pathogenic DNA,pathogenic DNA: TLR9,phytohemagglutinin,poly I: C,ssRNA,ssRNA: TLR7,ssRNA: TLR8
Induction of type I interferons (IFNs) by viruses and other pathogens is crucialfor innate immunity, and it is mediated by the activation of pattern-recognitionreceptors, such as Toll-like receptors and cytosolic receptors such as RIG-I andMDA5. The type I IFN induction is primarily controlled at the genetranscriptional level, wherein a family of transcription factors, interferonregulatory factors (IRFs), plays central roles. Here, we summarize the recentstudies on IRFs, providing a paradigm of how genes are ingeniously regulatedduring immune responses. We also consider some evolutional aspects on theIFN-IRF system.
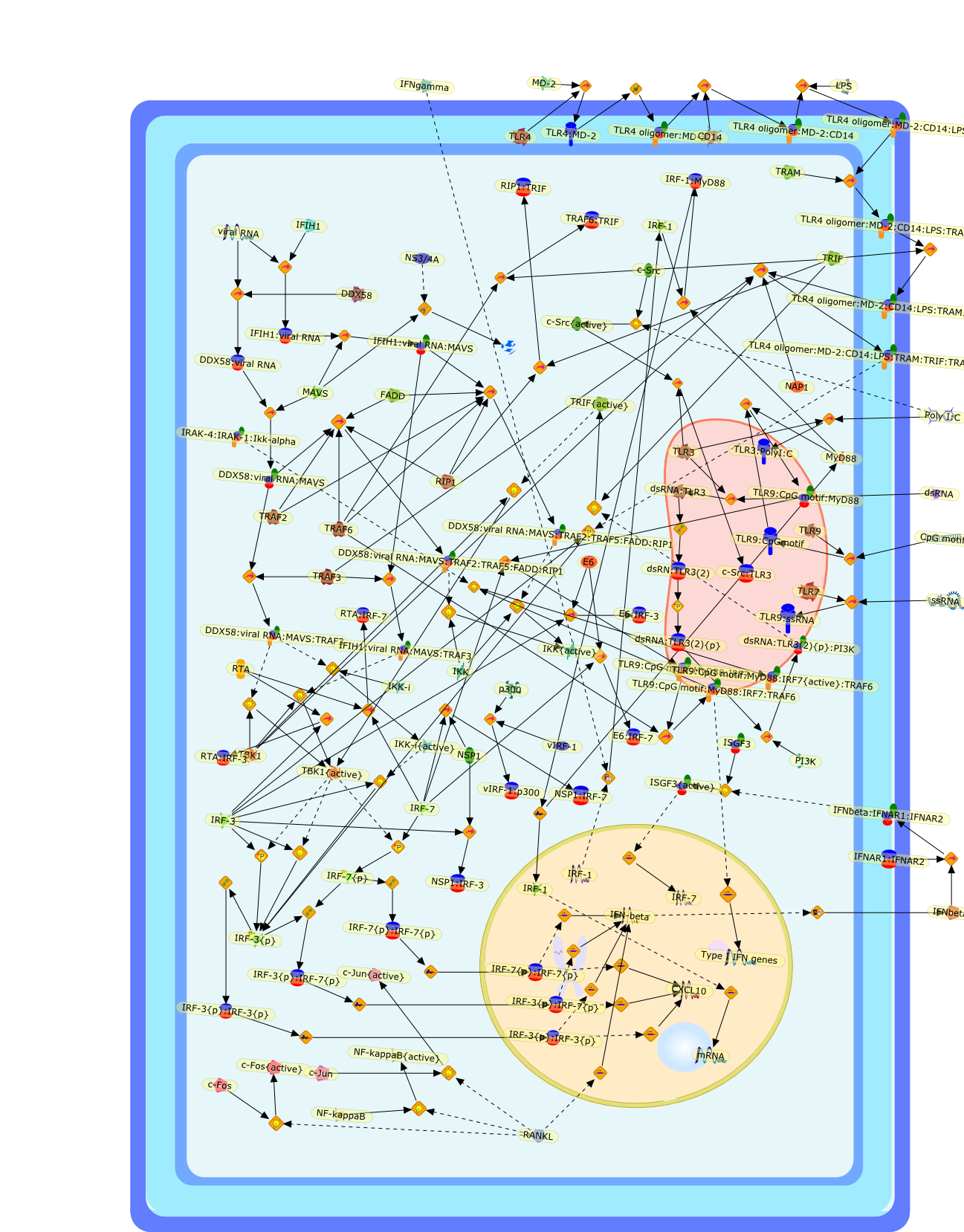{kind=link}
{kind=link}
CD14,CXCL10,CpG motif,DDX58,DDX58:viral RNA,DDX58:viral RNA:MAVS,DDX58:viral RNA:MAVS:TRAF2:TRAF5:FADD:RIP1,DDX58:viral RNA:MAVS:TRAF3,E6,E6:IRF-3,E6:IRF-7,FADD,IFIH1,IFIH1:viral RNA,IFIH1:viral RNA:MAVS,IFIH1:viral RNA:MAVS:TRAF3,IFN-beta,IFNAR1:IFNAR2,IFNbeta,IFNbeta:IFNAR1:IFNAR2,IFNgamma,IKK,IKK-i,IKK-i{active},IKK{active},IRAK-4:IRAK-1:Ikk-alpha,IRF-1,IRF-1:MyD88,IRF-3,IRF-3{p},IRF-3{p}:IRF-3{p},IRF-3{p}:IRF-7{p},IRF-7,IRF-7{p},IRF-7{p}:IRF-7{p},ISGF3,ISGF3{active},LPS,MAVS,MD-2,MyD88,NAP1,NF-kappaB,NF-kappaB{active},NS3/4A,NSP1,NSP1:IRF-3,NSP1:IRF-7,PI3K,Poly I:C,RANKL,RIP1,RIP1:TRIF,RTA,RTA:IRF-3,RTA:IRF-7,TBK1,TBK1{active},TLR3,TLR3:PolyI:C,TLR4,TLR4 oligomer:MD-2,TLR4 oligomer:MD-2:CD14,TLR4 oligomer:MD-2:CD14:LPS,TLR4 oligomer:MD-2:CD14:LPS:TRAM,TLR4 oligomer:MD-2:CD14:LPS:TRAM:TRIF,TLR4 oligomer:MD-2:CD14:LPS:TRAM:TRIF:TRAF3:NAP1:TBK1,TLR4:MD-2,TLR7,TLR9,TLR9:CpG motif:MyD88,TLR9:CpG motif:MyD88:IRF7,TLR9:CpG motif:MyD88:IRF7:TRAF6,TLR9:CpG motif:MyD88:IRF7{active}:TRAF6,TLR9:CpGmotif,TLR9:ssRNA,TRAF2,TRAF3,TRAF6,TRAF6:TRIF,TRAM,TRIF,TRIF{active},Type I IFN genes,c-Fos,c-Fos{active},c-Jun,c-Jun{active},c-Src,c-Src:TLR3,c-Src{active},dsRN:TLR3(2),dsRNA,dsRNA:TLR3,dsRNA:TLR3(2){p},dsRNA:TLR3(2){p}:PI3K,mRNA,p300,protein remnants,ssRNA,vIRF-1,vIRF-1:p300,viral RNA
DNA in microbes or host cells is normally sequestered from the immune system,and therefore inert, but becomes an active immunostimulatory molecule duringinfection or tissue damage. Recent evidence suggests that Toll-like receptor(TLR)9, currently the only known immune sensor for DNA, recognizes more diverseelements in its ligand than initially thought, and must cooperate withadditional host factors to provoke an optimal innate immune response in thephysiological environment. Moreover, the innate immune system possesses aTLR9-independent, as-yet-undefined intracellular recognition machinery ofdouble-stranded DNA that induces type I interferons through distinct signalingpathways. TLR9-dependent and TLR9-independent immune recognition of DNA mightplay crucial roles in DNA-associated protective immunity and in pathologicalautoimmunity.
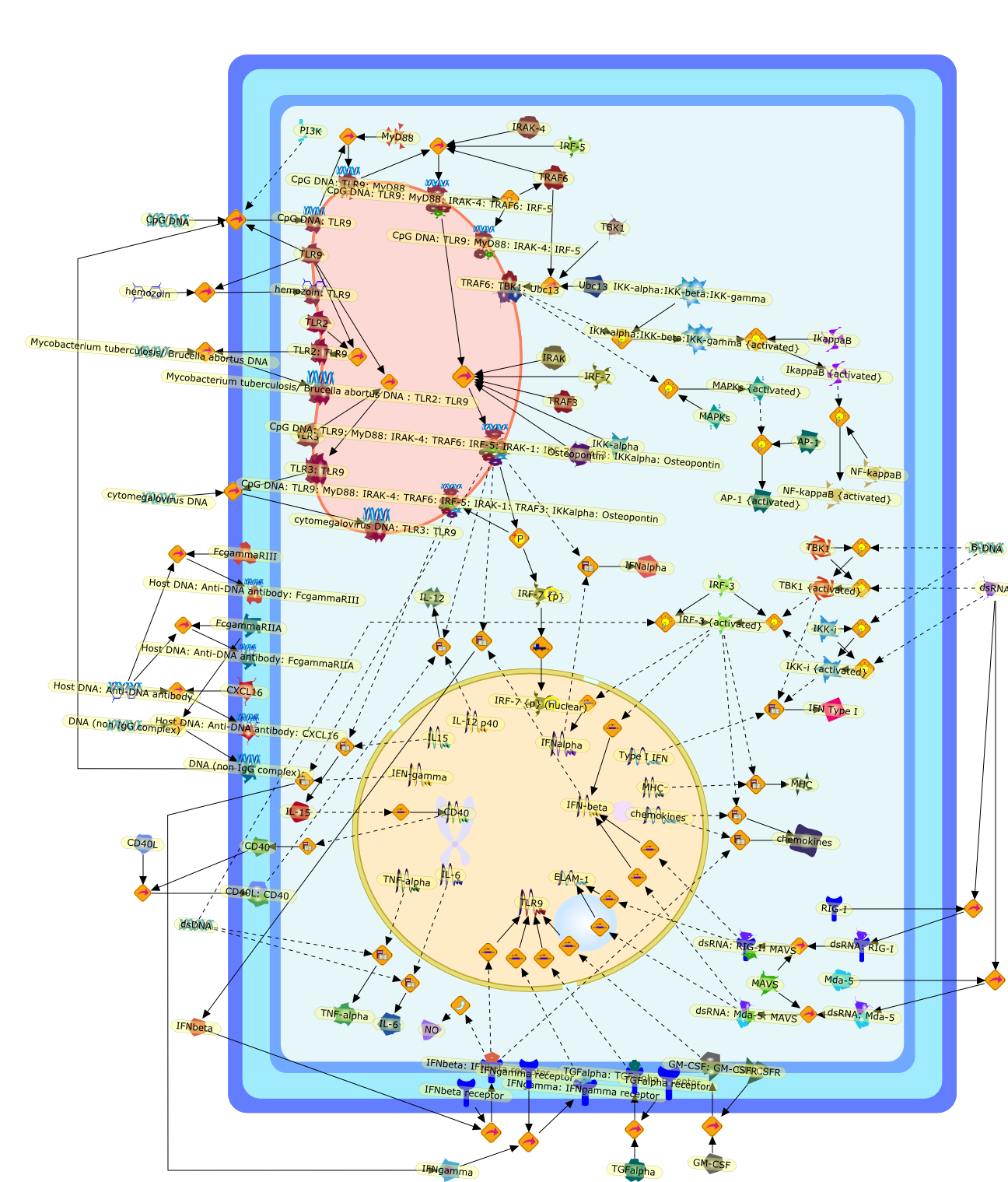{kind=link}
{kind=link}
AP-1,AP-1 {activated},B-DNA,CD40,CD40L,CD40L: CD40,CXCL16,CpG DNA,CpG DNA: TLR9,CpG DNA: TLR9: MyD88,CpG DNA: TLR9: MyD88: IRAK-4: IRF-5,CpG DNA: TLR9: MyD88: IRAK-4: TRAF6: IRF-5,CpG DNA: TLR9: MyD88: IRAK-4: TRAF6: IRF-5: IRAK-1: TRAF3: IKKalpha: Osteopontin,CpG DNA: TLR9: MyD88: IRAK-4: TRAF6: IRF-5: IRAK-1: IRF-7: TRAF3: IKKalpha: Osteopontin,DNA (non IgG complex),DNA (non IgG complex): ,ELAM-1,FcgammaRIIA,FcgammaRIII,GM-CSF,GM-CSF: GM-CSFR,GM-CSFR,Host DNA: Anti-DNA antibody,Host DNA: Anti-DNA antibody: CXCL16,Host DNA: Anti-DNA antibody: FcgammaRIIA,Host DNA: Anti-DNA antibody: FcgammaRIII,IFN Type I,IFN-beta,IFN-gamma,IFNalpha,IFNbeta,IFNbeta receptor,IFNbeta: IFNbeta receptor,IFNgamma,IFNgamma receptor,IFNgamma: IFNgamma receptor,IKK-alpha,IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma {activated},IKK-i,IKK-i {activated},IL-12,IL-12 p40,IL-15,IL-6,IL15,IRAK,IRAK-4,IRF-3,IRF-3 {activated},IRF-5,IRF-7,IRF-7 {p},IRF-7 {p} (nuclear),IkappaB,IkappaB {activated},MAPKs,MAPKs {activated},MAVS,MHC,Mda-5,MyD88,Mycobacterium tuberculosis/ Brucella abortus DNA,Mycobacterium tuberculosis/ Brucella abortus DNA : TLR2: TLR9,NF-kappaB,NF-kappaB {activated},NO,Osteopontin,PI3K,RIG-I,TBK1,TBK1 {activated},TGFalpha,TGFalpha receptor,TGFalpha: TGFalpha receptor,TLR2,TLR2: TLR9,TLR3,TLR3: TLR9,TLR9,TNF-alpha,TRAF3,TRAF6,TRAF6: TBK1: Ubc13,Type I IFN,Ubc13,chemokines,cytomegalovirus DNA,cytomegalovirus DNA: TLR3: TLR9,dsDNA,dsRNA,dsRNA: Mda-5,dsRNA: Mda-5: MAVS,dsRNA: RIG-I,dsRNA: RIG-I: MAVS,hemozoin,hemozoin: TLR9
The transcription factor nuclear factor-kappaB (NF-kappaB) has a pivotal role ininitiating inflammation and raising an effective immune response. BecauseNF-kappaB activation depends on ubiquitination, cells have developed ubiquitin(Ub)-mediated strategies for inhibiting NF-kappaB activation and preventingexcessive inflammation. Recent findings concerning tumor necrosis factor (TNF)receptor and toll-like receptor (TLR)-interleukin-1 (IL-1) receptor signallingpathways show that Ub can be a tool as well as a target for NF-kappaB inhibitoryproteins, either by labelling specific signalling proteins forproteasome-dependent degradation or by serving as a target for specificde-ubiquitinating enzymes that prevent the formation of pertinent signallingcomplexes. Interfering with ubiquitination therefore seems to be a versatilemeans for regulating NF-kappaB activity, indicating that studies of Ub-mediatedsignalling might hold the key for developing new therapeutic strategies forinflammatory disease.
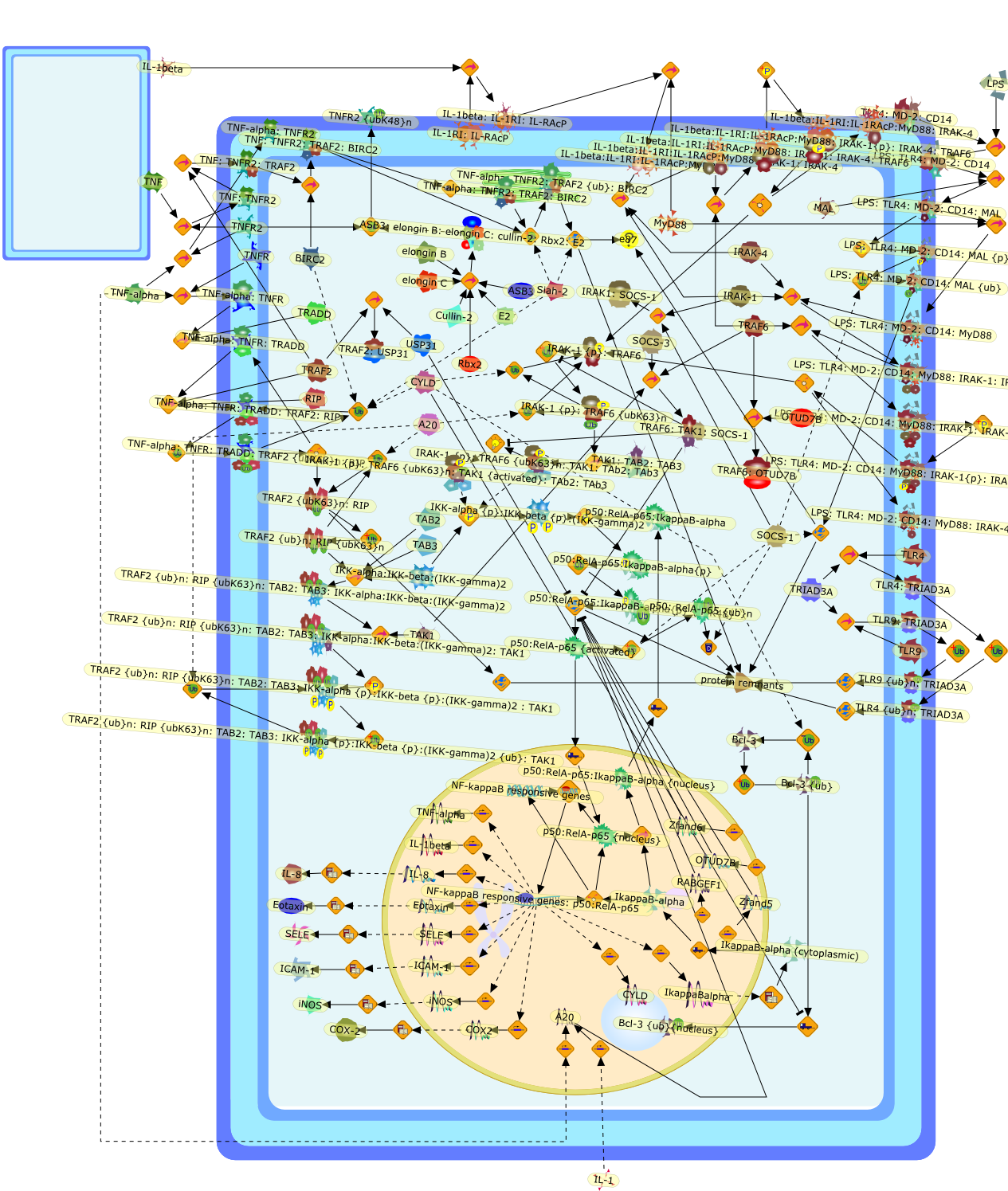{kind=link}
{kind=link}
A20,ASB3,ASB3: elongin B: elongin C: cullin-2: Rbx2: E2,BIRC2,Bcl-3,Bcl-3 {ub},Bcl-3 {ub}{nucleus},COX-2,COX2,CYLD,Cullin-2,E2,Eotaxin,ICAM-1,IKK-alpha {p}:IKK-beta {p}:(IKK-gamma)2,IKK-alpha:IKK-beta:(IKK-gamma)2,IL-1,IL-1RI: IL-RAcP,IL-1beta,IL-1beta: IL-1RI: IL-RAcP,IL-1beta:IL-1RI:IL-1RAcP:MyD88,IL-1beta:IL-1RI:IL-1RAcP:MyD88: IRAK-1: IRAK-4,IL-1beta:IL-1RI:IL-1RAcP:MyD88: IRAK-1: IRAK-4: TRAF6,IL-1beta:IL-1RI:IL-1RAcP:MyD88: IRAK-1{p}: IRAK-4: TRAF6,IL-1beta:IL-1RI:IL-1RAcP:MyD88: IRAK-4,IL-8,IRAK-1,IRAK-1 {p}: TRAF6,IRAK-1 {p}: TRAF6 {ubK63}n,IRAK-1 {p}: TRAF6 {ubK63}n: TAK1 {activated}: TAb2: TAb3,IRAK-1 {p}: TRAF6 {ubK63}n: TAK1: TAb2: TAb3,IRAK-4,IRAK1: SOCS-1,IkappaB-alpha,IkappaB-alpha (cytoplasmic),IkappaBalpha,LPS,LPS: TLR4: MD-2: CD14,LPS: TLR4: MD-2: CD14: MAL,LPS: TLR4: MD-2: CD14: MAL {p},LPS: TLR4: MD-2: CD14: MAL {ub},LPS: TLR4: MD-2: CD14: MyD88,LPS: TLR4: MD-2: CD14: MyD88: IRAK-1: IRAK-4,LPS: TLR4: MD-2: CD14: MyD88: IRAK-1: IRAK-4: TRAF6,LPS: TLR4: MD-2: CD14: MyD88: IRAK-1{p}: IRAK-4: TRAF6,LPS: TLR4: MD-2: CD14: MyD88: IRAK-4,MAL,MyD88,NF-kappaB responsive genes,NF-kappaB responsive genes: p50:RelA-p65,OTUD7B,RABGEF1,RIP,Rbx2,SELE,SOCS-1,SOCS-3,Siah-2,TAB2,TAB3,TAK1,TAK1: TAB2: TAB3,TLR4,TLR4 {ub}n: TRIAD3A,TLR4: MD-2: CD14,TLR4: TRIAD3A,TLR9,TLR9 {ub}n: TRIAD3A,TLR9: TRIAD3A,TNF,TNF-alpha,TNF-alpha: TNFR,TNF-alpha: TNFR2,TNF-alpha: TNFR2: TRAF2 {ub}: BIRC2,TNF-alpha: TNFR2: TRAF2: BIRC2,TNF-alpha: TNFR: TRADD,TNF-alpha: TNFR: TRADD: TRAF2 {ubK63}n: RIP,TNF-alpha: TNFR: TRADD: TRAF2: RIP,TNF: TNFR2,TNF: TNFR2: TRAF2,TNF: TNFR2: TRAF2: BIRC2,TNFR,TNFR2,TNFR2 {ubK48}n,TRADD,TRAF2,TRAF2 {ubK63}n: RIP,TRAF2 {ub}n: RIP {ubK63}n,TRAF2 {ub}n: RIP {ubK63}n: TAB2: TAB3: IKK-alpha {p}:IKK-beta {p}:(IKK-gamma)2 : TAK1,TRAF2 {ub}n: RIP {ubK63}n: TAB2: TAB3: IKK-alpha {p}:IKK-beta {p}:(IKK-gamma)2 {ub}: TAK1,TRAF2 {ub}n: RIP {ubK63}n: TAB2: TAB3: IKK-alpha:IKK-beta:(IKK-gamma)2,TRAF2 {ub}n: RIP {ubK63}n: TAB2: TAB3: IKK-alpha:IKK-beta:(IKK-gamma)2: TAK1,TRAF2: USP31,TRAF6,TRAF6: OTUD7B,TRAF6: TAK1: SOCS-1,TRIAD3A,USP31,Zfand5,Zfand6,e87,elongin B,elongin C,iNOS,p50: RelA-p65 {ub}n,p50:RelA-p65 {activated},p50:RelA-p65 {nucleus},p50:RelA-p65:IkappaB-alpha,p50:RelA-p65:IkappaB-alpha {nucleus},p50:RelA-p65:IkappaB-alpha{p},p50:RelA-p65:IkappaB-alpha{p} {ubK48},protein remnants
The human body is protected against external pathogens by two immune systems:innate and acquired immunities. Whereas innate immunity exhibits immediateresponses to external pathogens by recognizing pathogen-associated molecularpatterns (PAMPs), adaptive immunity uses T cells to recognize and defend againstpathogens by developing effector cells, antibodies and memory cells. Althougheach system seems to possess distinct activation mechanisms, interleukin-1receptor-associated kinase (IRAK)-4 is essential for NF-kappaB activation inToll-like receptor (TLR) and T-cell receptor (TCR) signaling pathways. Thisimplies possible crosstalk between innate and acquired immunities, andevolutionary development that resulted in the use of innate signaling moleculesby the acquired immune system. Here, we discuss the impact of theseevolutionarily conserved molecules on innate and acquired immunity, and theirpotential as drug targets for the simultaneous modulation of both immunities.
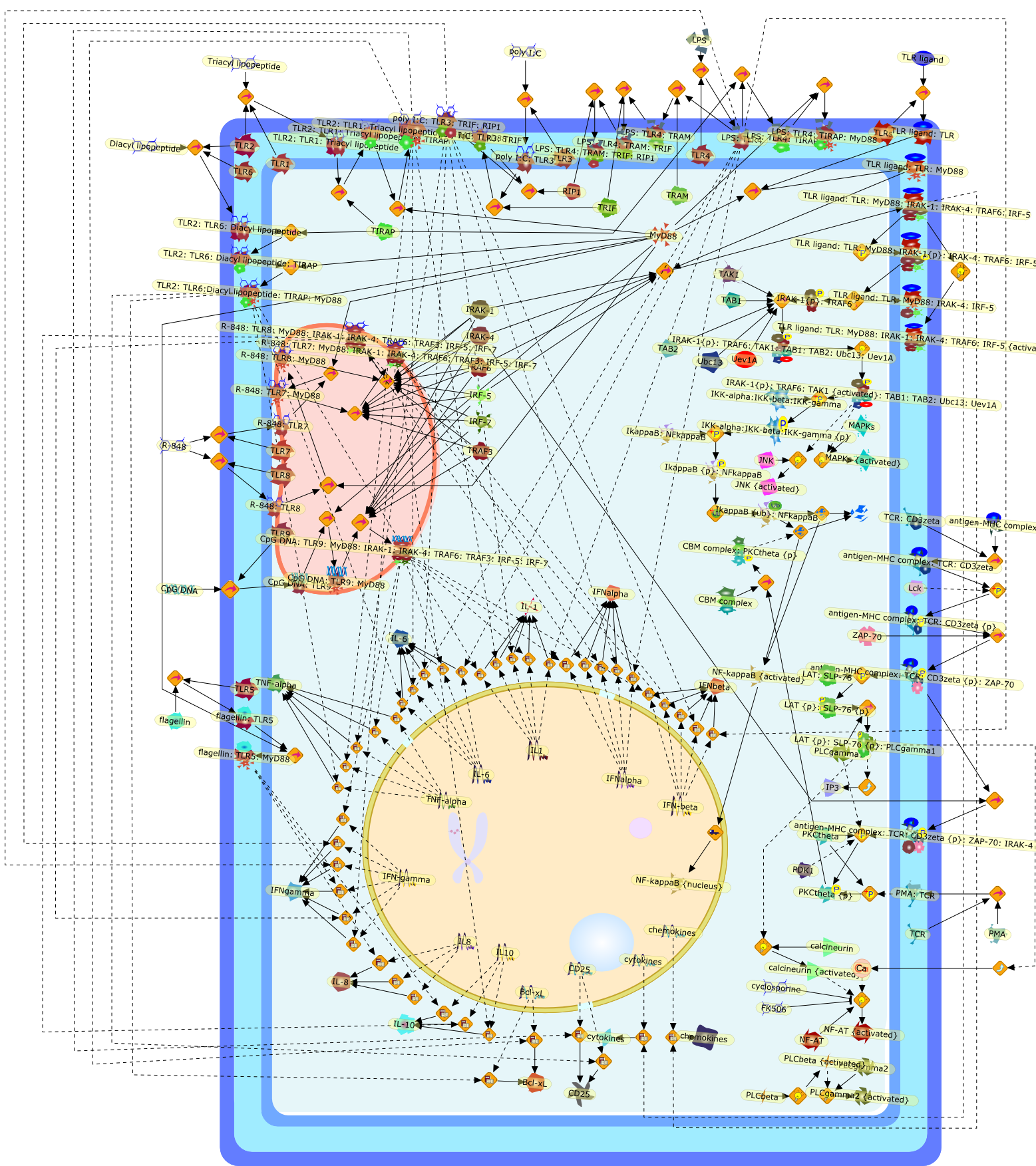{kind=link}
{kind=link}
Bcl-xL,CBM complex,CBM complex: PKCtheta {p},CD25,Ca,CpG DNA,CpG DNA: TLR9,CpG DNA: TLR9: MyD88,CpG DNA: TLR9: MyD88: IRAK-1: IRAK-4: TRAF6: TRAF3: IRF-5: IRF-7,Diacyl lipopeptide,FK506,IFN-beta,IFN-gamma,IFNalpha,IFNbeta,IFNgamma,IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma {p},IL-1,IL-10,IL-6,IL-8,IL1,IL10,IL8,IP3,IRAK-1,IRAK-1{p}: TRAF6,IRAK-1{p}: TRAF6: TAK1 {activated}: TAB1: TAB2: Ubc13: Uev1A,IRAK-1{p}: TRAF6: TAK1: TAB1: TAB2: Ubc13: Uev1A,IRAK-4,IRF-5,IRF-7,IkappaB {p}: NFkappaB,IkappaB {ub}: NFkappaB,IkappaB: NFkappaB,JNK,JNK {activated},LAT {p}: SLP-76 {p},LAT {p}: SLP-76 {p}: PLCgamma1,LAT: SLP-76,LPS,LPS: TLR4,LPS: TLR4: TIRAP,LPS: TLR4: TIRAP: MyD88,LPS: TLR4: TRAM,LPS: TLR4: TRAM: TRIF,LPS: TLR4: TRAM: TRIF: RIP1,Lck,MAPKs,MAPKs {activated},MyD88,NF-AT,NF-AT {activated},NF-kappaB {activated},NF-kappaB {nucleus},PDK1,PKCtheta,PKCtheta {p},PLCbeta,PLCbeta {activated},PLCgamma1,PLCgamma2,PLCgamma2 {activated},PMA,PMA: TCR,R-848,R-848: TLR7,R-848: TLR7: MyD88,R-848: TLR7: MyD88: IRAK-1: IRAK-4: TRAF6: TRAF3: IRF-5: IRF-7,R-848: TLR8,R-848: TLR8: MyD88,R-848: TLR8: MyD88: IRAK-1: IRAK-4: TRAF6: TRAF3: IRF-5: IRF-7,RIP1,TAB1,TAB2,TAK1,TCR,TCR: CD3zeta,TIRAP,TLR ligand,TLR ligand: TLR,TLR ligand: TLR: MyD88,TLR ligand: TLR: MyD88: IRAK-1: IRAK-4: TRAF6: IRF-5,TLR ligand: TLR: MyD88: IRAK-1: IRAK-4: TRAF6: IRF-5 {activated},TLR ligand: TLR: MyD88: IRAK-1{p}: IRAK-4: TRAF6: IRF-5,TLR ligand: TLR: MyD88: IRAK-4: IRF-5,TLR1,TLR2,TLR2: TLR1: Triacyl lipopeptide,TLR2: TLR1: Triacyl lipopeptide: TIRAP,TLR2: TLR1: Triacyl lipopeptide: TIRAP: MyD88,TLR2: TLR6: Diacyl lipopeptide,TLR2: TLR6: Diacyl lipopeptide: TIRAP,TLR2: TLR6:Diacyl lipopeptide: TIRAP: MyD88,TLR3,TLR4,TLR5,TLR6,TLR7,TLR8,TLR9,TLRs,TNF-alpha,TRAF3,TRAF6,TRAM,TRIF,Triacyl lipopeptide,Ubc13,Uev1A,ZAP-70,antigen-MHC complex,antigen-MHC complex: TCR: CD3zeta,antigen-MHC complex: TCR: CD3zeta {p},antigen-MHC complex: TCR: CD3zeta {p}: ZAP-70,antigen-MHC complex: TCR: CD3zeta {p}: ZAP-70: IRAK-4,calcineurin,calcineurin {activated},chemokines,cyclosporine,cytokines,flagellin,flagellin: TLR5,flagellin: TLR5: MyD88,poly I:C,poly I:C: TLR3,poly I:C: TLR3: TRIF,poly I:C: TLR3: TRIF: RIP1,protein ramnents
Adenosine is an endogenous purine nucleoside that, following its release intothe extracellular space, binds to specific adenosine receptors expressed on thecell surface. Adenosine appears in the extracellular space under metabolicallystressful conditions, which are associated with ischemia, inflammation, and celldamage. There are 4 types of adenosine receptors (A(1), A(2A), A(2B) and A(3))and all adenosine receptors are members of the G protein-coupled family ofreceptors. Adenosine receptors are expressed on monocytes and macrophages andthrough these receptors adenosine modulates monocyte and macrophage function.Since monocytes and macrophages are activated by the same danger signals thatcause accumulation of extracellular adenosine, adenosine receptors expressed onmacrophages represent a sensor system that provide monocytes and macrophageswith information about the stressful environment. Adenosine receptors, thus,allow monocytes and macrophages to fine-tune their responses to stressfulstimuli. Here, we review the consequences of adenosine receptor activation onmonocyte/macrophage function. We will detail the effect of stimulating thevarious adenosine receptor subtypes on macrophage differentiation/proliferation,phagocytosis, and tissue factor (TF) expression. We will also summarize ourknowledge of how adenosine impacts the production of extracellular mediatorssecreted by monocytes and macrophages in response to toll-like receptor (TLR)ligands and other inflammatory stimuli. Specifically, we will delineate howadenosine affects the production of superoxide, nitric oxide (NO), tumornecrosis factor-alpha, interleukin (IL)-12, IL-10, and vascular endothelialgrowth factor (VEGF). A deeper insight into the regulation of monocyte andmacrophage function by adenosine receptors should assist in developing newtherapies for inflammatory diseases.
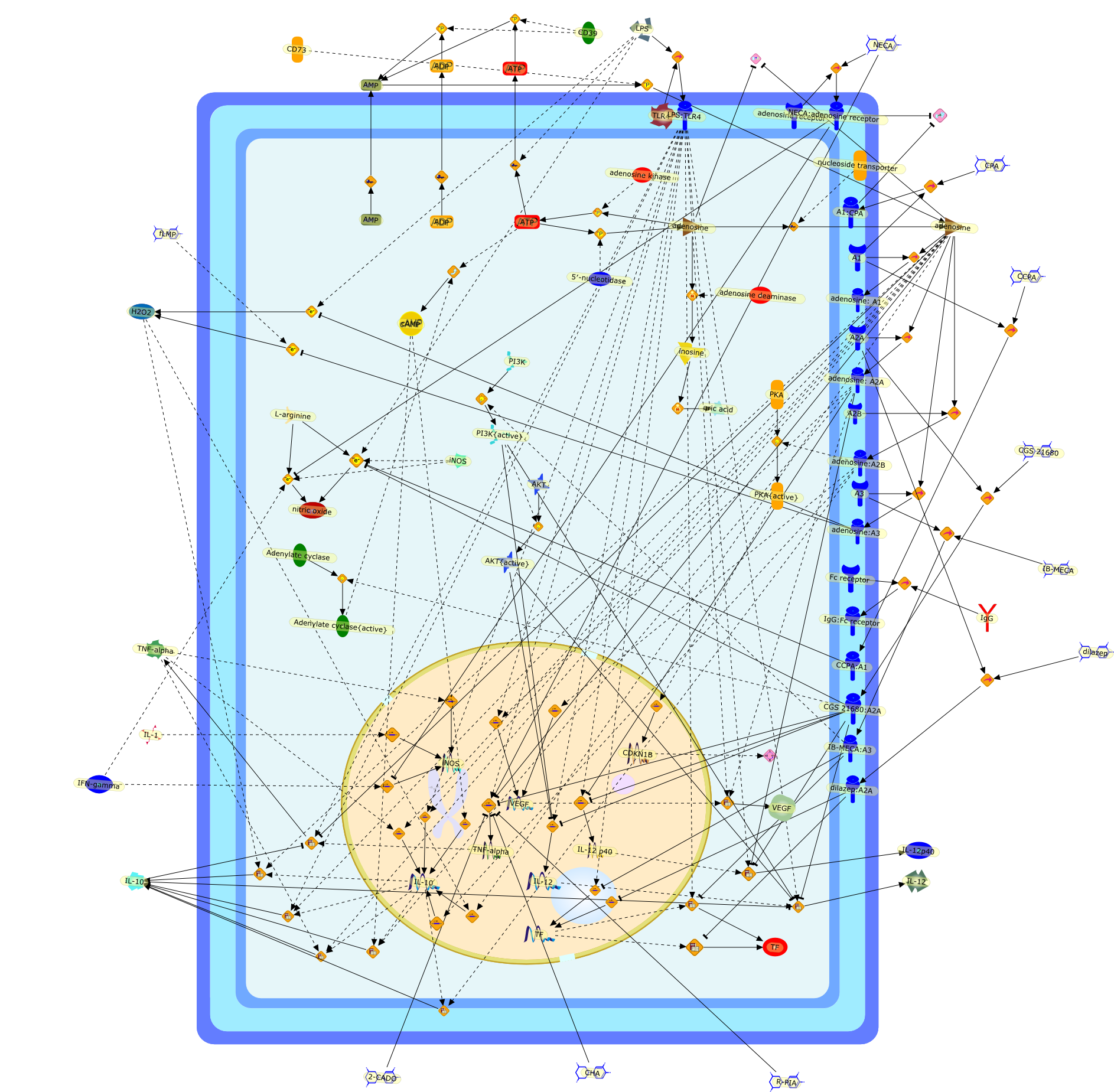{kind=link}
{kind=link}
2-CADO,5′-nucleotidase,A1,A1:CPA,A2A,A2B,A3,ADP,AKT,AKT{active},AMP,ATP,Adenylate cyclase,Adenylate cyclase{active},CCPA,CCPA:A1,CD39,CD73,CDKN1B,CGS 21680,CGS 21680:A2A,CHA,CPA,Fc receptor,H2O2,IB-MECA,IB-MECA:A3,IFN-gamma,IL-1,IL-10,IL-12,IL-12 p40,IL-12p40,IgG,IgG:Fc receptor,L-arginine,LPS,LPS:TLR4,NECA,NECA:adenosine receptor,PI3K,PI3K{active},PKA,PKA{active},R-PIA,TF,TLR4,TNF-alpha,VEGF,adenosine,adenosine deaminase,adenosine kinase,adenosine receptor,adenosine: A1,adenosine: A2A,adenosine:A2B,adenosine:A3,cAMP,dilazep,dilazep:A2A,fLMP,iNOS,inosine,nitric oxide,nucleoside transporter,uric acid
Cytokine receptors act through a complex signaling network, involving Januskinases (JAKs) and the signal transducers and activators of transcription(STATs), to regulate diverse biological processes which control growth,development, homeostasis and immune function, among others. The JAK/STATsignaling pathway is attenuated via three mechanisms controlling the initiation,magnitude, and duration of the signal: the PIAS proteins, which prevent STATdimerization or DNA interaction, the SHP phosphatases, which dephosphorylateactivating tyrosine phosphorylations, and the suppressors of cytokine signaling(SOCS), which are transcribed in response to cytokine stimulation and useseveral interconnected mechanisms to downregulate the signal. Specific studiestargeting the SOCS genes in vivo have unveiled SOCS2 as the main regulator ofsomatic growth through regulation of GH/IGF-1 signaling. In addition, severalstudies indicate that SOCS2 also has important actions in the central nervoussystem, the regulation of metabolism, the immune response, the mammary glanddevelopment, cancer, and other cytokine-dependent signaling pathways. Consistentwith the role of cytokines in human physiology, any SOCS2 imbalance could resultin a broad range of pathologies such as cardiovascular diseases, insulinresistance, cancer, and severe infections, among others. Thus, determining theimportance of SOCS2 in health and disease will no doubt aid in the developmentof novel therapeutic strategies. In this review, we attempt to summarize theavailable information, including our results, regarding the role of SOCS2 inseveral biological processes.
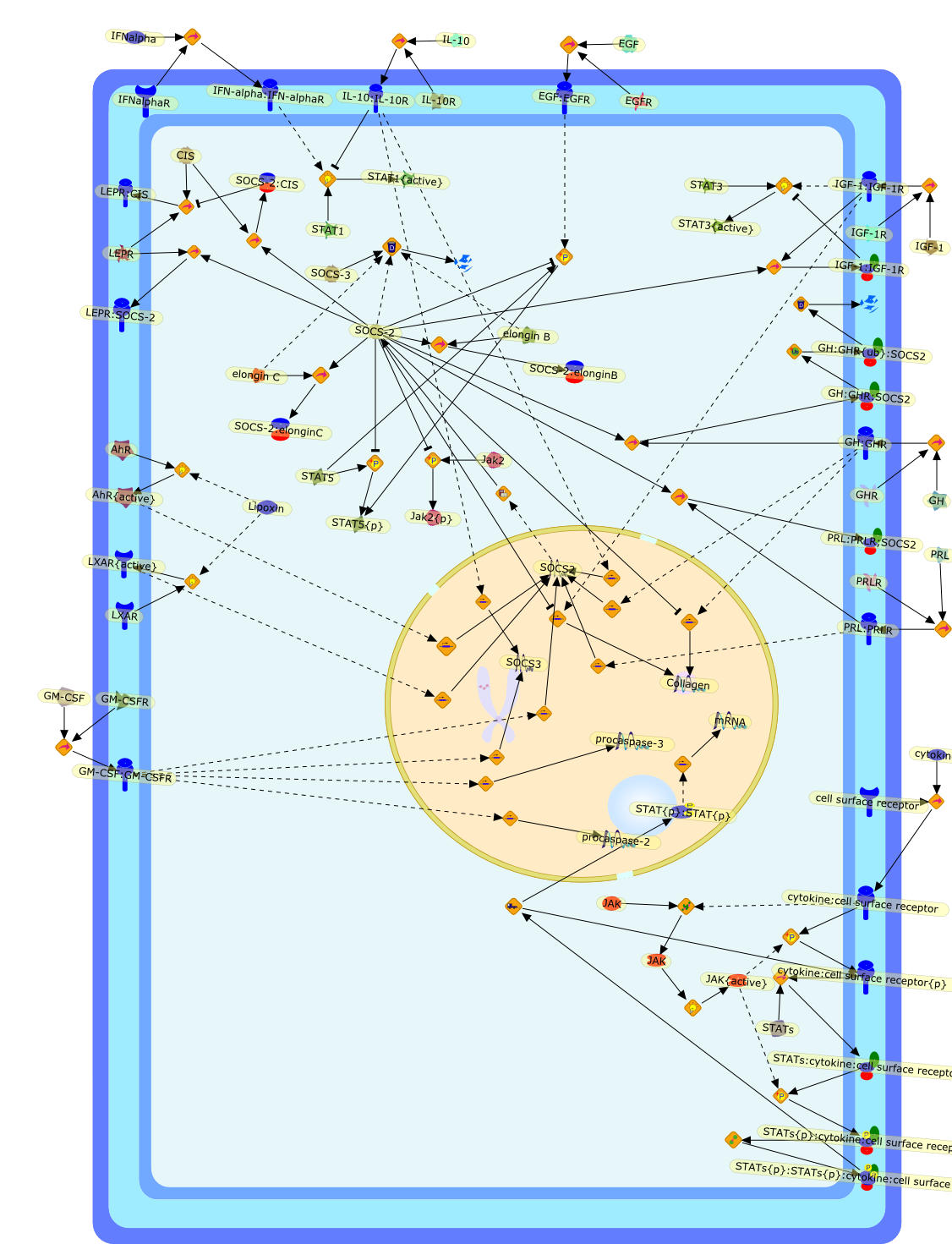{kind=link}
{kind=link}
AhR,AhR{active},CIS,Collagen,Degradant,Degradants,EGF,EGF:EGFR,EGFR,GH,GH:GHR,GH:GHR:SOCS2,GH:GHR{ub}:SOCS2,GHR,GM-CSF,GM-CSF:GM-CSFR,GM-CSFR,IFN-alpha:IFN-alphaR,IFNalpha,IFNalphaR,IGF-1,IGF-1:IGF-1R,IGF-1R,IL-10,IL-10:IL-10R,IL-10R,JAK,JAK{active},Jak2,Jak2{p},LEPR,LEPR:CIS,LEPR:SOCS-2,LXAR,LXAR{active},Lipoxin,PRL,PRL:PRLR,PRL:PRLR;SOCS2,PRLR,SOCS-2,SOCS-2:CIS,SOCS-2:elonginB,SOCS-2:elonginC,SOCS-3,SOCS2,SOCS3,STAT1,STAT1{active},STAT3,STAT3{active},STAT5,STAT5{p},STATs,STATs:cytokine:cell surface receptor,STATs{p}:STATs{p}:cytokine:cell surface receptor,STATs{p}:cytokine:cell surface receptor,STAT{p}:STAT{p},cell surface receptor,cytokine,cytokine:cell surface receptor,cytokine:cell surface receptor{p},elongin B,elongin C,mRNA,procaspase-2,procaspase-3
The immune system requires precise regulation of activating and inhibitorysignals so that it can mount effective responses against pathogens whileensuring tolerance to self-components. Some of the most potent activationsignals are triggered by innate immune molecules, particularly those in the TLRfamily. Recent studies have shown that engagement of TLRs plays a significantrole in both innate and adaptive immunity. This review focuses on the ways thatTLR function might contribute to the etiology of lupus-like syndromes in thecontext of an autoimmune-prone environment. By considering the sources,localization, and expression of both nucleic acids and the molecules that bindthem, we discuss several ways that innate immunity can play a role in thedevelopment of systemic autoimmunity.
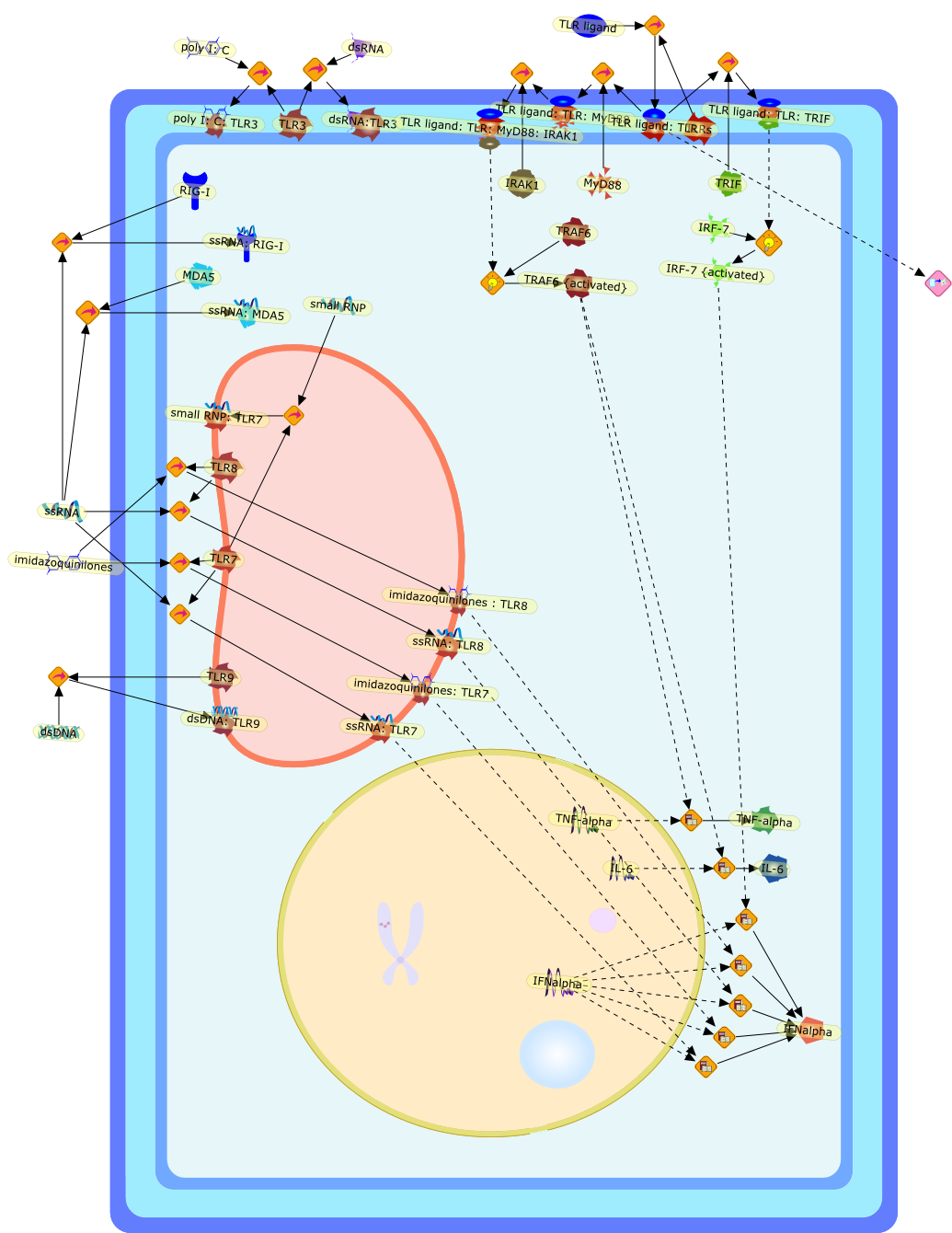{kind=link}
{kind=link}
IFNalpha,IL-6,IRAK1,IRF-7,IRF-7 {activated},MDA5,MyD88,RIG-I,TLR ligand,TLR ligand: TLR,TLR ligand: TLR: MyD88,TLR ligand: TLR: MyD88: IRAK1,TLR ligand: TLR: TRIF,TLR3,TLR7,TLR8,TLR9,TLRs,TNF-alpha,TRAF6,TRAF6 {activated},TRIF,dsDNA,dsDNA: TLR9,dsRNA,dsRNA:TLR3,imidazoquinilones,imidazoquinilones : TLR8,imidazoquinilones: TLR7,poly I: C,poly I: C: TLR3,small RNP,small RNP: TLR7,ssRNA,ssRNA: MDA5,ssRNA: RIG-I,ssRNA: TLR7,ssRNA: TLR8
The MAPK family members p38, JNK, and ERK are all activated downstream of innateimmunity's TLR to induce the production of cytokines and inflammatory mediators.However, the relative intensity and duration of the activation of different MAPKappears to determine the type of immune response. The mammalian genome encodes alarge number of dual specificity phosphatases (DUSP), many of which act as MAPKphosphatases. In this study, we review the emergence of several DUSP as genesthat are differentially expressed and regulated in immune cells. Recently, aseries of investigations in mice deficient in DUSP1, DUSP2, or DUSP10 revealedspecificity in the regulation of the different MAPK proteins, and definedessential roles in models of local and systemic inflammation. The DUSP family isproposed as a set of molecular control devices specifying and modulating MAPKsignaling, which may be targeted to unleash or attenuate innate and adaptiveimmune effector functions.
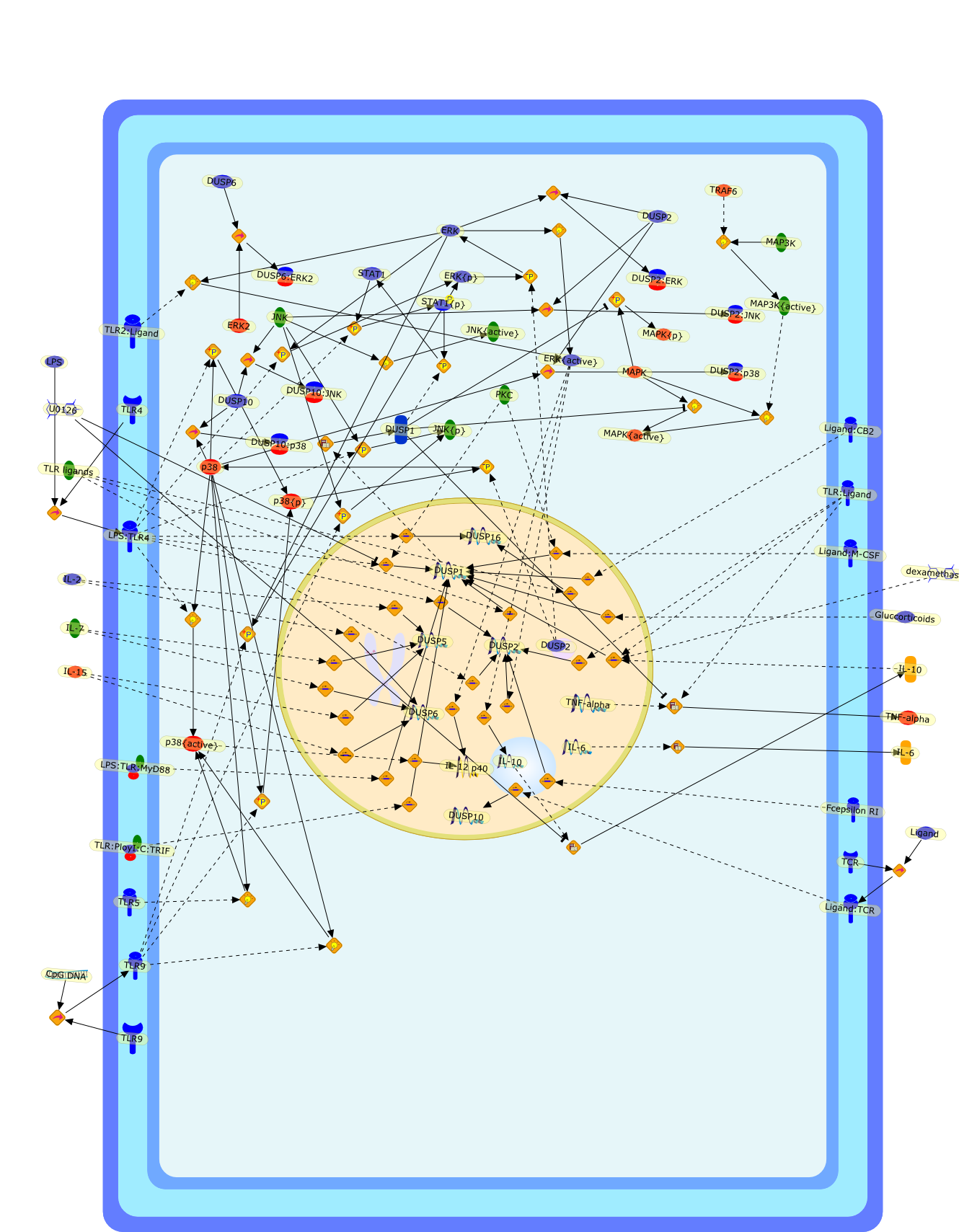{kind=link}
{kind=link}
CpG DNA,DUSP1,DUSP10,DUSP10:JNK,DUSP10:p38,DUSP16,DUSP2,DUSP2:ERK,DUSP2:JNK,DUSP2:p38,DUSP5,DUSP6,DUSP6:ERK2,ERK,ERK2,ERK{active},ERK{p},Fcepsilon RI,Gluccorticoids,IL-10,IL-12 p40,IL-15,IL-2,IL-6,IL-7,JNK,JNK{active},JNK{p},LPS,LPS:TLR4,LPS:TLR:MyD88,Ligand,Ligand:CB2,Ligand:M-CSF,Ligand:TCR,MAP3K,MAP3K{active},MAPK,MAPK{active},MAPK{p},PKC,STAT1,STAT1{p},TCR,TLR ligands,TLR2:Ligand,TLR4,TLR5,TLR9,TLR:Ligand,TLR:PloyI:C:TRIF,TNF-alpha,TRAF6,U0126,dexamethasone,p38,p38{active},p38{p}
There have been new developments in the elucidation of the biological functionsof the inter-alpha-inhibitor (IalphaI) family. The anti-proteolytic activity ofthe IalphaI family originates from bikunin (also known as urinary trypsininhibitor). Growing evidence indicates that bikunin is not just ananti-proteolytic agent, but can also be considered an anti-inflammatory agentthat suppresses lipopolysaccharide (LPS)-induced cytokine synthesis. Bikuninfunctions to inhibit calcium influx and extracellular signal-regulated kinase(ERK) signaling via LPS receptors and/or as yet unidentified bikunin signalingreceptors. By signaling via the LPS receptor, LPS increases calcium influx andyields phosphorylated ERK, which activates multiple transcription factors, suchas nuclear factor kappaB (NF-kappaB) or early growth response-1 (Egr-1), whichin turn promote cytokine expression. Deficits in the signaling cascades causedby free or cell-bound bikunin are predicted to down-regulate cytokineexpression, render macrophages/neutrophils more inactive, and impairinflammatory processes. This brief review largely focuses on our currentunderstanding of the apparent functions of bikunin, its ligands, the effectormolecules with which it interacts, and its regulation.
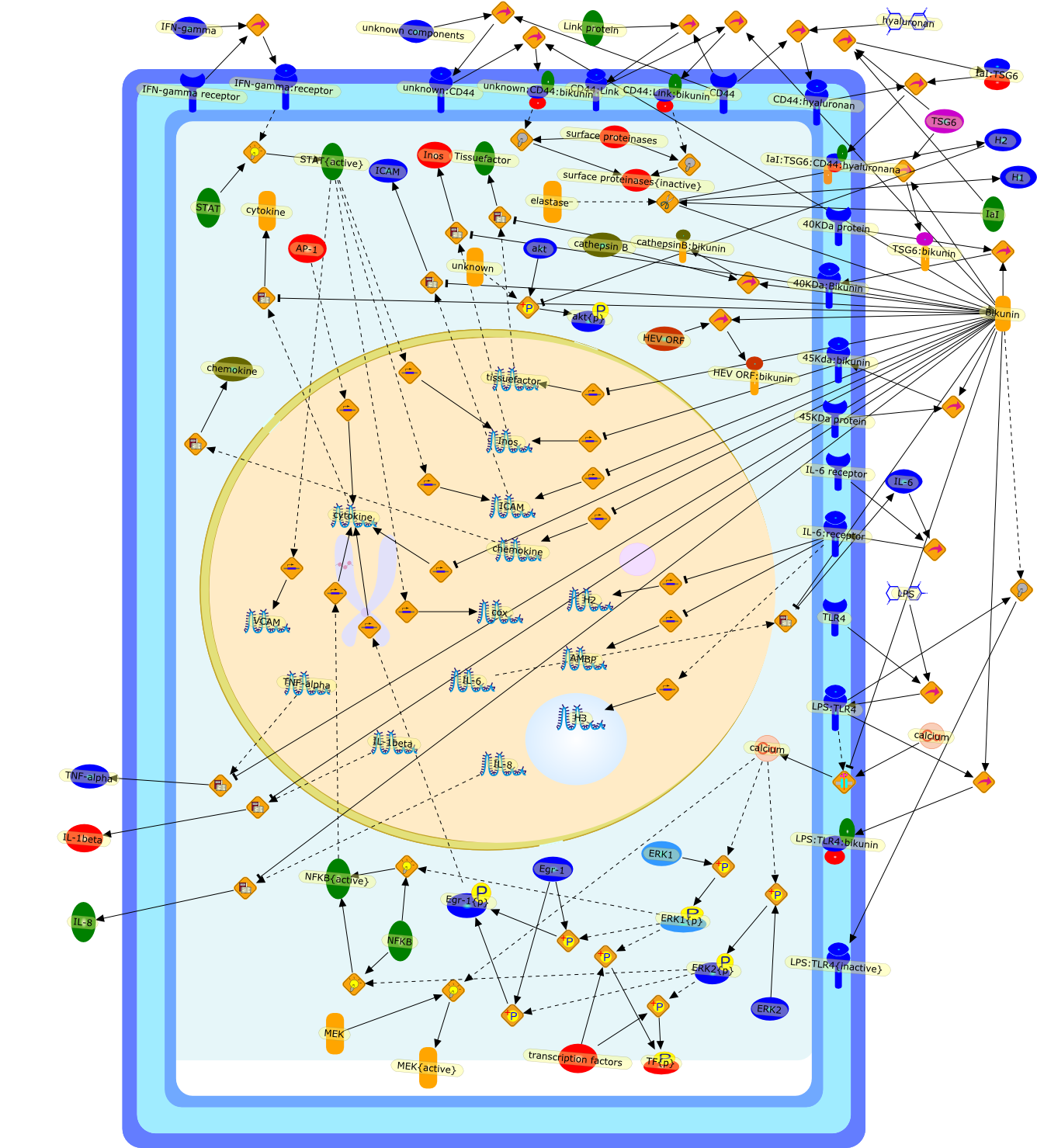{kind=link}
{kind=link}
H2,40KDa protein,40KDa:Bikunin,45KDa protein,45Kda:bikunin,AMBP,AP-1,Bikunin,CD44,CD44:Link,CD44:Link:bikunin,CD44:hyaluronan,ERK1,ERK1{p},ERK2,ERK2{p},Egr-1,Egr-1{p},H1,H2,H3,HEV ORF,HEV ORF:bikunin,ICAM,IFN-gamma,IFN-gamma receptor,IFN-gamma:receptor,IL-1beta,IL-6,IL-6 receptor,IL-6:receptor,IL-8,IaI,IaI:TSG6,IaI:TSG6:CD44:hyaluronana,Inos,LPS,LPS:TLR4,LPS:TLR4:bikunin,LPS:TLR4{inactive},Link protein,MEK,MEK{active},NFKB,NFKB{active},STAT,STAT{active},TF{p},TLR4,TNF-alpha,TSG6,TSG6:bikunin,Tissuefactor,VCAM,akt,akt{p},calcium,cathepsin B,cathepsinB:bikunin,chemokine,cox,cytokine,elastase,hyaluronan,surface proteinases,surface proteinases{inactive},tissuefactor,transcription factors,unknown,unknown components,unknown:CD44,unknown:CD44:bikunin
Toll-like receptor (TLR) signals induce dendritic cell (DC) differentiation andinfluence the immunological outcome of their interactions with T cells. Recentin vitro studies demonstrate that TLR signals also trigger strikingreorganisation of the DC vacuolar compartments, the cytoskeleton and themachinery of protein translation and turnover. Moreover, TLR ligation withinendosomes and phagosomes appears to establish organelle autonomous signals.These changes, which mostly occur within minutes to a few hours after TLRengagement, are adaptations relevant to the antigen capture, processing andmigratory phases of the DC life history.
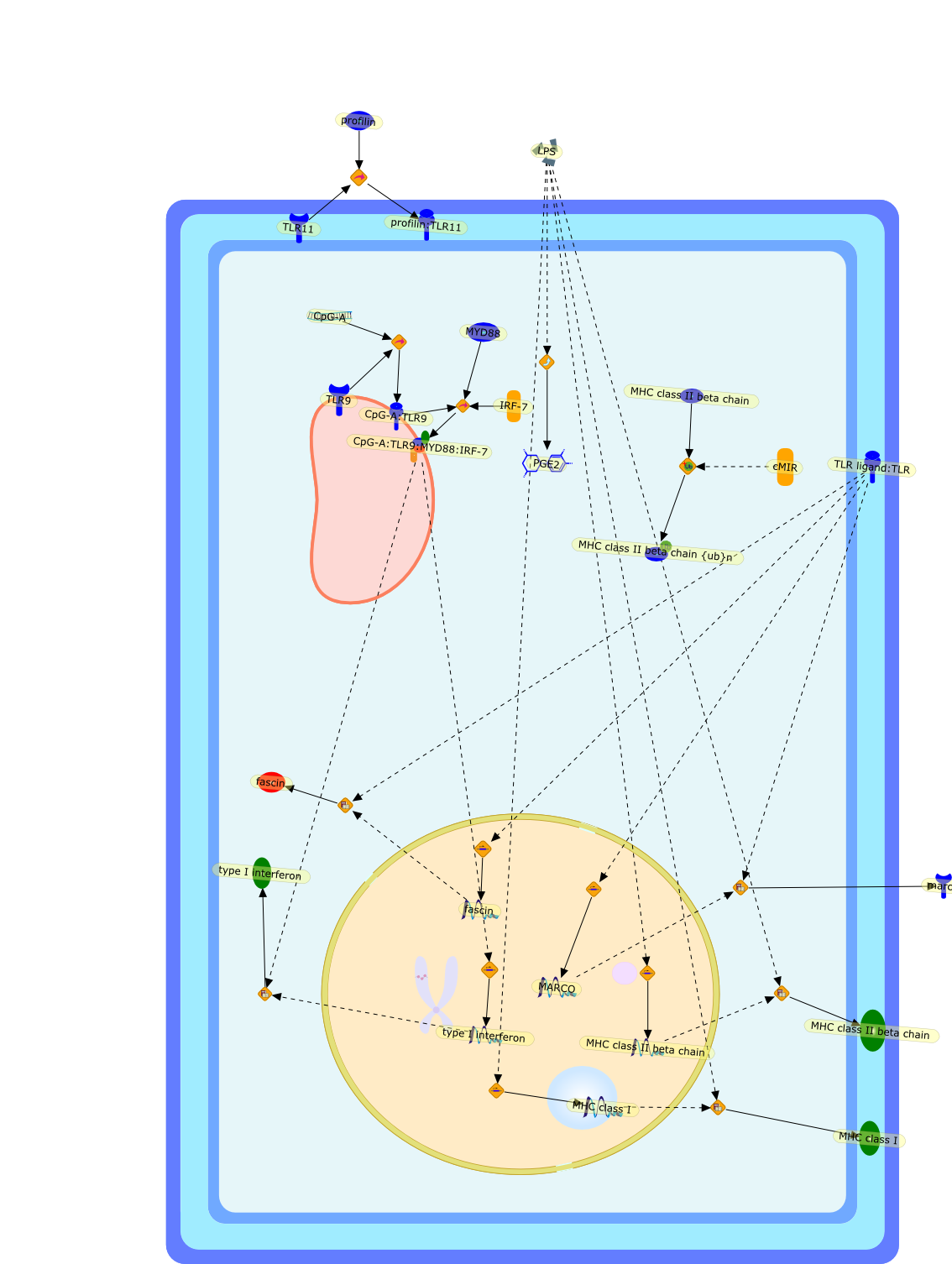{kind=link}
{kind=link}
CpG-A,CpG-A:TLR9,CpG-A:TLR9:MYD88:IRF-7,IRF-7,LPS,MARCO,MHC class I,MHC class II beta chain,MHC class II beta chain {ub}n,MYD88,PGE2,TLR ligand:TLR,TLR11,TLR9,cMIR,fascin,marco,profilin,profilin:TLR11,type I interferon
Recognition of bacteria by the vertebrate innate immune system relies ondetection of invariant molecules by specialized receptors. The view is emergingthat activation of both Toll-like receptors (TLRs) and Nod-like receptors (NLRs)by different bacterial agonists is important in order to mount an inflammatoryresponse in the host. Priming of cells with peptidoglycan and products that aresensed by cytosolic-localized members of the NLR family have a synergisticeffect on TLR signalling and vice versa. Currently, the underlying molecularmechanisms of this cross-talk between NLR and TLR signalling are beginning toemerge. These reveal that the two sensing-systems are non-redundant in bacterialrecognition and that their cross-talk plays an important role in immunologicalhomeostasis.
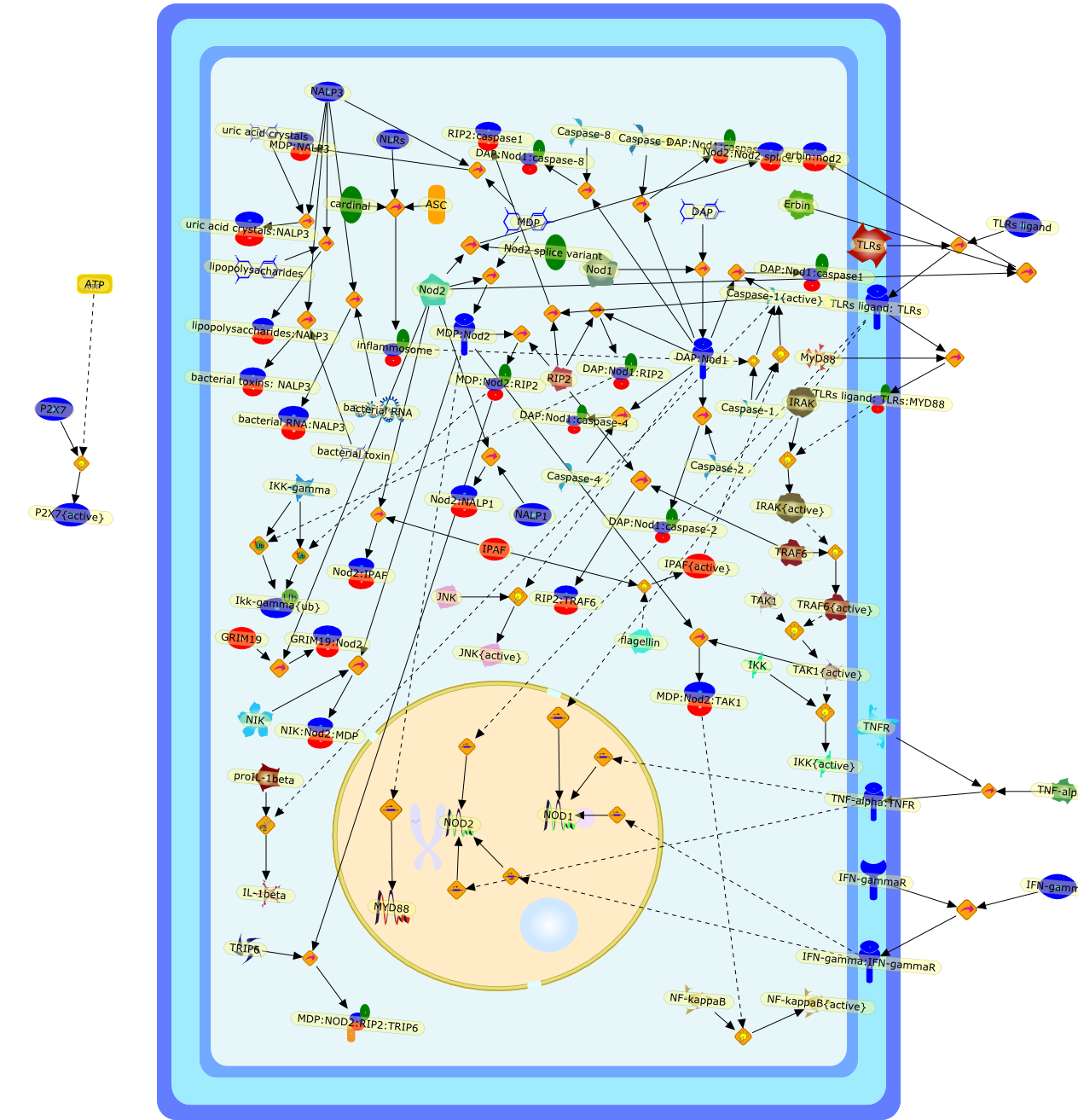{kind=link}
{kind=link}
ASC,ATP,Caspase-1,Caspase-1{active},Caspase-2,Caspase-4,Caspase-8,Caspase-9,DAP,DAP:Nod1,DAP:Nod1:RIP2,DAP:Nod1:caspase-2,DAP:Nod1:caspase-4,DAP:Nod1:caspase-8,DAP:Nod1:caspase-9,DAP:Nod1:caspase1,Erbin,GRIM19,GRIM19:Nod2,IFN-gamma,IFN-gamma:IFN-gammaR,IFN-gammaR,IKK,IKK-gamma,IKK{active},IL-1beta,IPAF,IPAF{active},IRAK,IRAK{active},Ikk-gamma{ub},JNK,JNK{active},MDP,MDP:NALP3,MDP:NOD2:RIP2:TRIP6,MDP:Nod2,MDP:Nod2:RIP2,MDP:Nod2:TAK1,MYD88,MyD88,NALP1,NALP3,NF-kappaB,NF-kappaB{active},NIK,NIK:Nod2:MDP,NLRs,NOD1,NOD2,Nod1,Nod2,Nod2 splice variant,Nod2:IPAF,Nod2:NALP1,Nod2:Nod2 splice variant,P2X7,P2X7{active},RIP2,RIP2:TRAF6,RIP2:caspase1,TAK1,TAK1{active},TLRs,TLRs ligand,TLRs ligand: TLRs,TLRs ligand: TLRs:MYD88,TNF-alpha,TNF-alpha:TNFR,TNFR,TRAF6,TRAF6{active},TRIP6,bacterial RNA,bacterial RNA:NALP3,bacterial toxin,bacterial toxins: NALP3,cardinal,erbin:nod2,flagellin,inflammosome,lipopolysaccharides:NALP3,lipopolysacharides,proIL-1beta,uric acid crystals,uric acid crystals:NALP3
The Toll-like receptor (TLR) family provide key components of mammalian immunityand are part of the earliest surveillance mechanisms responding to infection.Their activation triggers the innate immune response, and is crucial to thesuccessful induction of Th1/Th2-phenotyped adaptive immunity. Innate immunitywas long considered to be non-specific and somewhat simple compared to adaptiveimmunity, mediated via the engulfment and lysis of microbial pathogens byphagocytic cells such as macrophages and neutrophils, and involving no complexprotein-protein interactions. The emergence of the TLR field has contributed toa revision of our understanding, and innate immunity is now viewed as a highlycomplex process, in line with adaptive immunity. This review will give a briefoverview of our current knowledge of TLR biology, and will focus on TLRs as keycomponents in complex networks that activate, integrate and select theappropriate innate and adaptive immune responses in the face of immunologicaldanger.
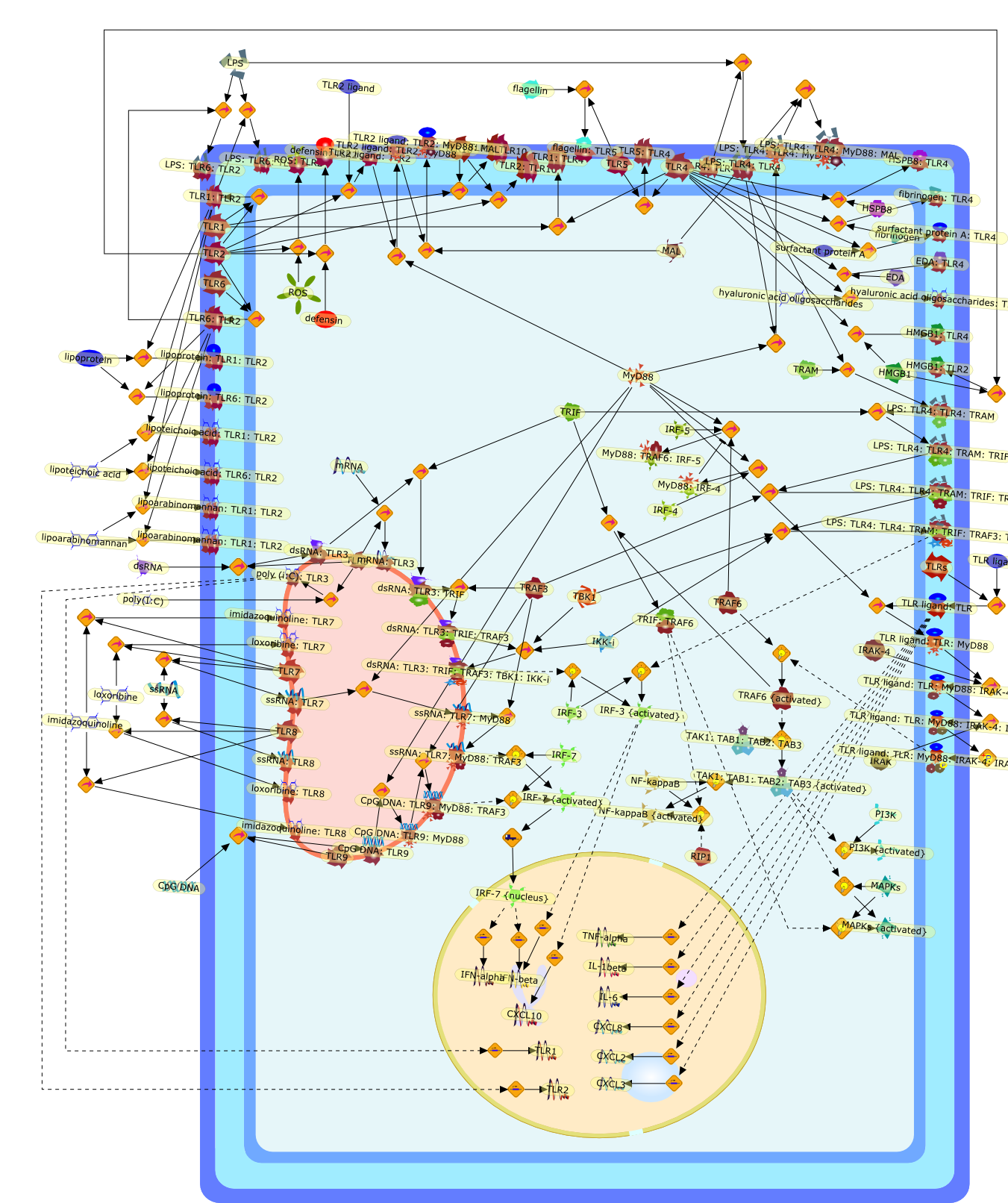{kind=link}
{kind=link}
CXCL10,CXCL2,CXCL3,CXCL8,CpG DNA,CpG DNA: TLR9,CpG DNA: TLR9: MyD88,CpG DNA: TLR9: MyD88: TRAF3,EDA,EDA: TLR4,HMGB1,HMGB1: TLR2,HMGB1: TLR4,HSPB8,HSPB8: TLR4,IFN-alpha,IFN-beta,IKK-i,IL-1beta,IL-6,IRAK,IRAK-4,IRF-3,IRF-3 {activated},IRF-4,IRF-5,IRF-7,IRF-7 {activated},IRF-7 {nucleus},LPS,LPS: TLR4: TLR4,LPS: TLR4: TLR4: MyD88,LPS: TLR4: TLR4: MyD88: MAL,LPS: TLR4: TLR4: TRAM,LPS: TLR4: TLR4: TRAM: TRIF,LPS: TLR4: TLR4: TRAM: TRIF: TRAF3,LPS: TLR4: TLR4: TRAM: TRIF: TRAF3: TBK1: IKK-i,LPS: TLR6: TLR2,MAL,MAPKs,MAPKs {activated},MyD88,MyD88: IRF-4,MyD88: TRAF6: IRF-5,NF-kappaB,NF-kappaB {activated},PI3K,PI3K {activated},RIP1,ROS,ROS: TLR2,TAK1: TAB1: TAB2: TAB3,TAK1: TAB1: TAB2: TAB3 {activated},TBK1,TLR ligand,TLR ligand: TLR,TLR ligand: TLR: MyD88,TLR ligand: TLR: MyD88: IRAK-4,TLR ligand: TLR: MyD88: IRAK-4: IRAK,TLR ligand: TLR: MyD88: IRAK-4: IRAK {p},TLR1,TLR10,TLR1: TLR10,TLR1: TLR2,TLR1: TLR4,TLR2,TLR2 ligand,TLR2 ligand: TLR2,TLR2 ligand: TLR2: MyD88,TLR2 ligand: TLR2: MyD88: MAL,TLR2: TLR10,TLR3,TLR4,TLR4: TLR4,TLR5,TLR5: TLR4,TLR6,TLR6: TLR2,TLR7,TLR8,TLR9,TLRs,TNF-alpha,TRAF3,TRAF6,TRAF6 {activated},TRAM,TRIF,TRIF: TRAF6,defensin,defensin: TLR2,dsRNA,dsRNA: TLR3,dsRNA: TLR3: TRIF,dsRNA: TLR3: TRIF: TRAF3,dsRNA: TLR3: TRIF: TRAF3: TBK1: IKK-i,fibrinogen,fibrinogen: TLR4,flagellin,flagellin: TLR5,hyaluronic acid oligosaccharides,hyaluronic acid oligosaccharides: TLR4,imidazoquinoline,imidazoquinoline: TLR7,imidazoquinoline: TLR8,lipoarabinomannan,lipoarabinomannan: TLR1: TLR2,lipoprotein,lipoprotein: TLR1: TLR2,lipoprotein: TLR6: TLR2,lipoteichoic acid,lipoteichoic acid: TLR1: TLR2,lipoteichoic acid: TLR6: TLR2,loxoribine,loxoribine: TLR7,loxoribine: TLR8,mRNA,mRNA: TLR3,poly (i:C): TLR3,poly(I:C),ssRNA,ssRNA: TLR7,ssRNA: TLR7: MyD88,ssRNA: TLR7: MyD88: TRAF3,ssRNA: TLR8,surfactant protein A,surfactant protein A: TLR4
(TLRs) respond to pathogens to initiate the innate immune response and directadaptive immunity, and evidence to date suggests that they have a role in thedetection of viruses. Many viral macromolecules have been shown to activateanti-viral signalling pathways via TLRs, leading to the induction of cytokinesand interferons, while viruses also have means of not only evading detection byTLRs, but also of subverting these receptors for their own purposes. This reviewdiscusses the role of TLRs in the context of other known viral detectionsystems, and examines some of the often surprising results from studies usingmice deficient in TLRs and their adaptors, in an attempt to unravel theparticular contribution of TLRs to anti-viral immunity.
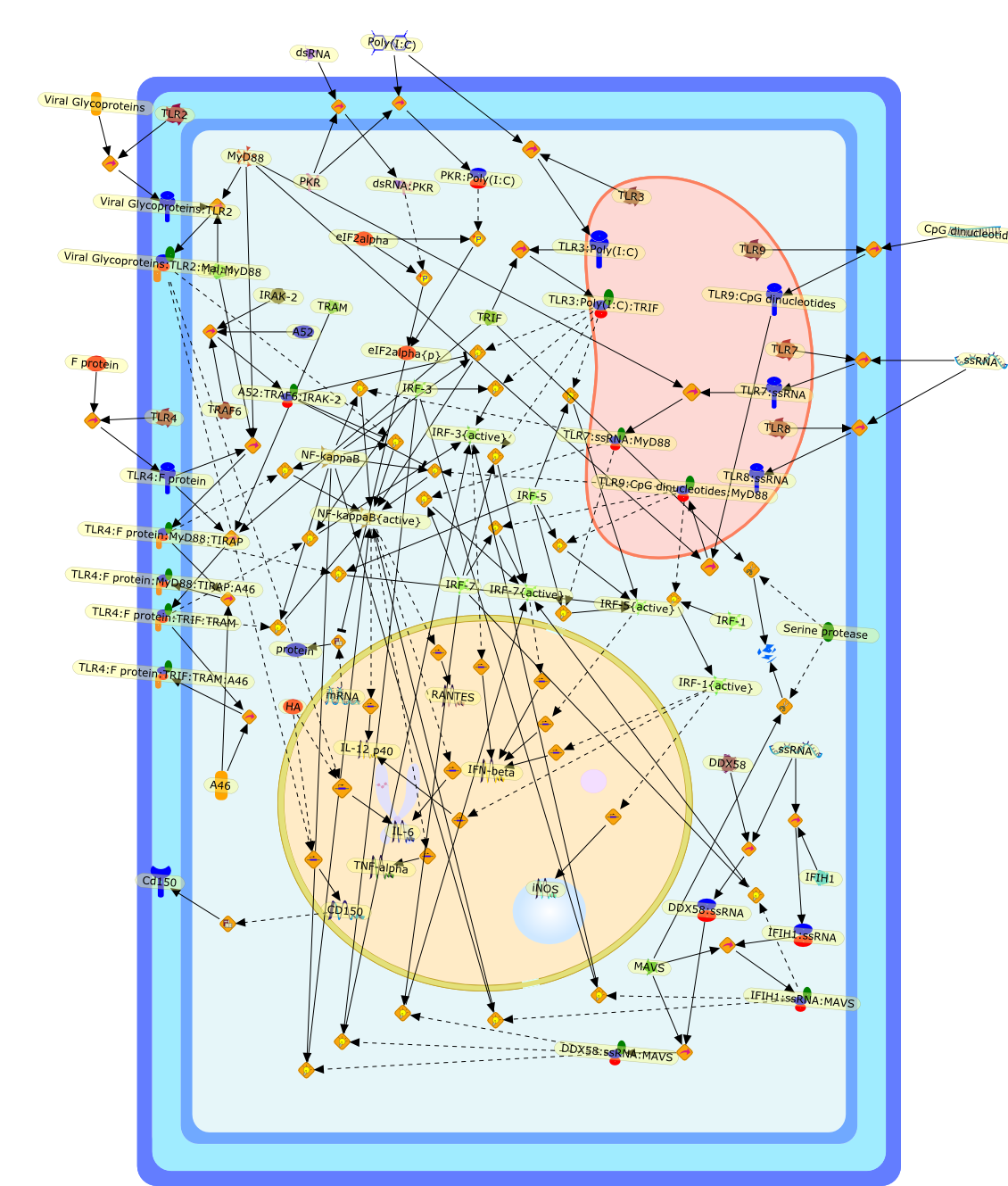{kind=link}
{kind=link}
A46,A52,A52:TRAF6:IRAK-2,CD150,Cd150,CpG dinucleotides,DDX58,DDX58:ssRNA,DDX58:ssRNA:MAVS,F protein,HA,IFIH1,IFIH1:ssRNA,IFIH1:ssRNA:MAVS,IFN-beta,IL-12 p40,IL-6,IRAK-2,IRF-1,IRF-1{active},IRF-3,IRF-3{active},IRF-5,IRF-5{active},IRF-7,IRF-7{active},MAVS,MyD88,NF-kappaB,NF-kappaB{active},PKR,PKR:Poly(I:C),Poly(I:C),RANTES,Serine protease,TIRAP,TLR2,TLR3,TLR3:Poly(I:C),TLR3:Poly(I:C):TRIF,TLR4,TLR4:F protein,TLR4:F protein:MyD88:TIRAP,TLR4:F protein:MyD88:TIRAP:A46,TLR4:F protein:TRIF:TRAM,TLR4:F protein:TRIF:TRAM:A46,TLR7,TLR7:ssRNA,TLR7:ssRNA:MyD88,TLR8,TLR8:ssRNA,TLR9,TLR9:CpG dinucleotides,TLR9:CpG dinucleotides:MyD88,TNF-alpha,TRAF6,TRAM,TRIF,Viral Glycoproteins,Viral Glycoproteins:TLR2,Viral Glycoproteins:TLR2:Mal:MyD88,dsRNA,dsRNA:PKR,eIF2alpha,eIF2alpha{p},iNOS,mRNA,protein,protein remnants,ssRNA
TLR trigger the induction of type I IFN (IFN-alpha/beta), providing a crucialmechanism of anti-viral defense. Until recently, TLR were thought to induce typeI IFN responses by activating two transcription factors which belong to theIFN-regulatory factor (IRF) family, IRF-3 and IRF-7. TLR-3 and TLR-4 induceIFN-beta by activating IRF-3; TLR-9 induces IFN-alpha and IFN-beta throughIRF-7, at least when engaged by type A CpG oligonucleotides (CpG-A) inplasmacytoid DC (pDC). In this issue of the European Journal of Immunology, itis demonstrated that TLR-9 induces IFN-beta when engaged by type B CpGoligonucleotides (CpG-B) in myeloid DC and macrophages. Remarkably, thisresponse is independent of IRF-3/7 and, in fact, requires another IRF familymember, IRF-1. IRF-1 is recruited by TLR-9 through the adaptor MyD88. Deficiencyof the TLR-9-->IRF-1-->IFN-beta pathway results in impaired anti-viral responsesnot only in vitro but also in vivo. These results demonstrate that TLR induceIFN-alpha or IFN-beta responses by activating distinct IRF, depending on the TLRligand and the cell type. These distinct TLR-IRF pathways may allow the immunesystem to tailor its responses to viral pathogens.
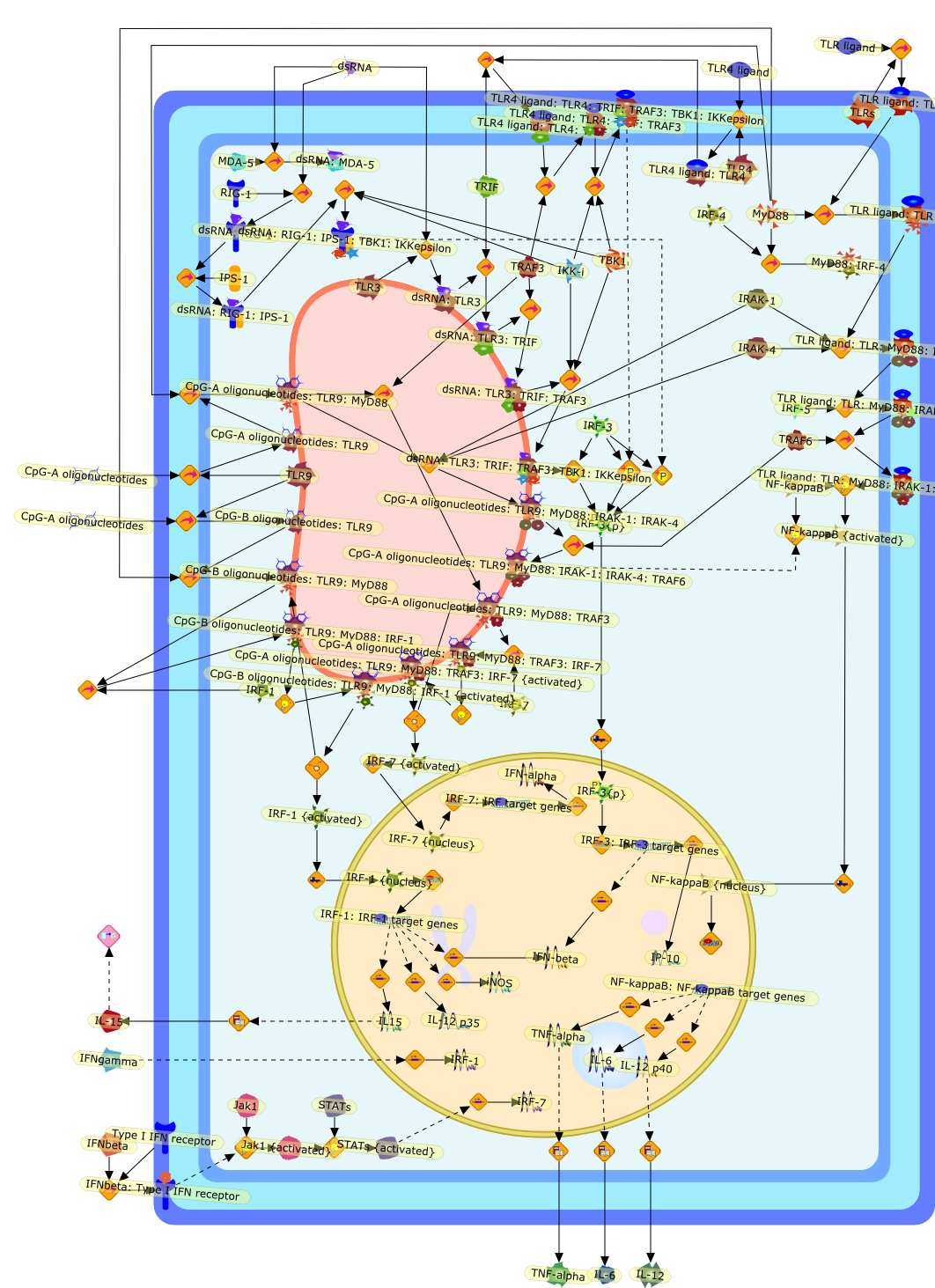{kind=link}
{kind=link}
CpG-A oligonucleotides,CpG-A oligonucleotides: TLR9,CpG-A oligonucleotides: TLR9: MyD88,CpG-A oligonucleotides: TLR9: MyD88: IRAK-1: IRAK-4,CpG-A oligonucleotides: TLR9: MyD88: IRAK-1: IRAK-4: TRAF6,CpG-A oligonucleotides: TLR9: MyD88: TRAF3,CpG-A oligonucleotides: TLR9: MyD88: TRAF3: IRF-7,CpG-A oligonucleotides: TLR9: MyD88: TRAF3: IRF-7 {activated},CpG-B oligonucleotides: TLR9,CpG-B oligonucleotides: TLR9: MyD88,CpG-B oligonucleotides: TLR9: MyD88: IRF-1,CpG-B oligonucleotides: TLR9: MyD88: IRF-1 {activated},IFN-alpha,IFN-beta,IFNbeta,IFNbeta: Type I IFN receptor,IFNgamma,IKK-i,IL-12,IL-12 p35,IL-12 p40,IL-15,IL-6,IL15,IP-10,IPS-1,IRAK-1,IRAK-4,IRF-1,IRF-1 {activated},IRF-1 {nucleus},IRF-1: IRF-1 target genes,IRF-3,IRF-3: IRF-3 target genes,IRF-3{p},IRF-4,IRF-5,IRF-7,IRF-7 {activated},IRF-7 {nucleus},IRF-7: IRF target genes,Jak1,Jak1 {activated},MDA-5,MyD88,MyD88: IRF-4,NF-kappaB,NF-kappaB {activated},NF-kappaB {nucleus},NF-kappaB: NF-kappaB target genes,RIG-1,STATs,STATs {activated},TBK1,TLR ligand,TLR ligand: TLR,TLR ligand: TLR: MyD88,TLR ligand: TLR: MyD88: IRAK-1: IRAK-4,TLR ligand: TLR: MyD88: IRAK-1: IRAK-4: IRF-5,TLR ligand: TLR: MyD88: IRAK-1: IRAK-4: IRF-5: TRAF6,TLR3,TLR4,TLR4 ligand,TLR4 ligand: TLR4,TLR4 ligand: TLR4: TRIF,TLR4 ligand: TLR4: TRIF: TRAF3,TLR4 ligand: TLR4: TRIF: TRAF3: TBK1: IKKepsilon,TLR9,TLRs,TNF-alpha,TRAF3,TRAF6,TRIF,Type I IFN receptor,dsRNA,dsRNA: MDA-5,dsRNA: RIG-1,dsRNA: RIG-1: IPS-1,dsRNA: RIG-1: IPS-1: TBK1: IKKepsilon,dsRNA: TLR3,dsRNA: TLR3: TRIF,dsRNA: TLR3: TRIF: TRAF3,dsRNA: TLR3: TRIF: TRAF3: TBK1: IKKepsilon,iNOS
The TLR family senses the molecular signatures of microbial pathogens, and playsa fundamental role in innate immune responses. TLRs signal via a common pathwaythat leads to the expression of diverse inflammatory genes. In addition, eachTLR elicits specific cellular responses to pathogens owing to differential usageof intracellular adapter proteins. Recent studies have revealed the importanceof the subcellular localization of TLRs in pathogen recognition and signaling.TLR signaling pathways is negatively regulated by a number of cellular proteinsto attenuate inflammation. Here, we describe recent advances in ourunderstanding of the regulation of TLR-mediated signaling.
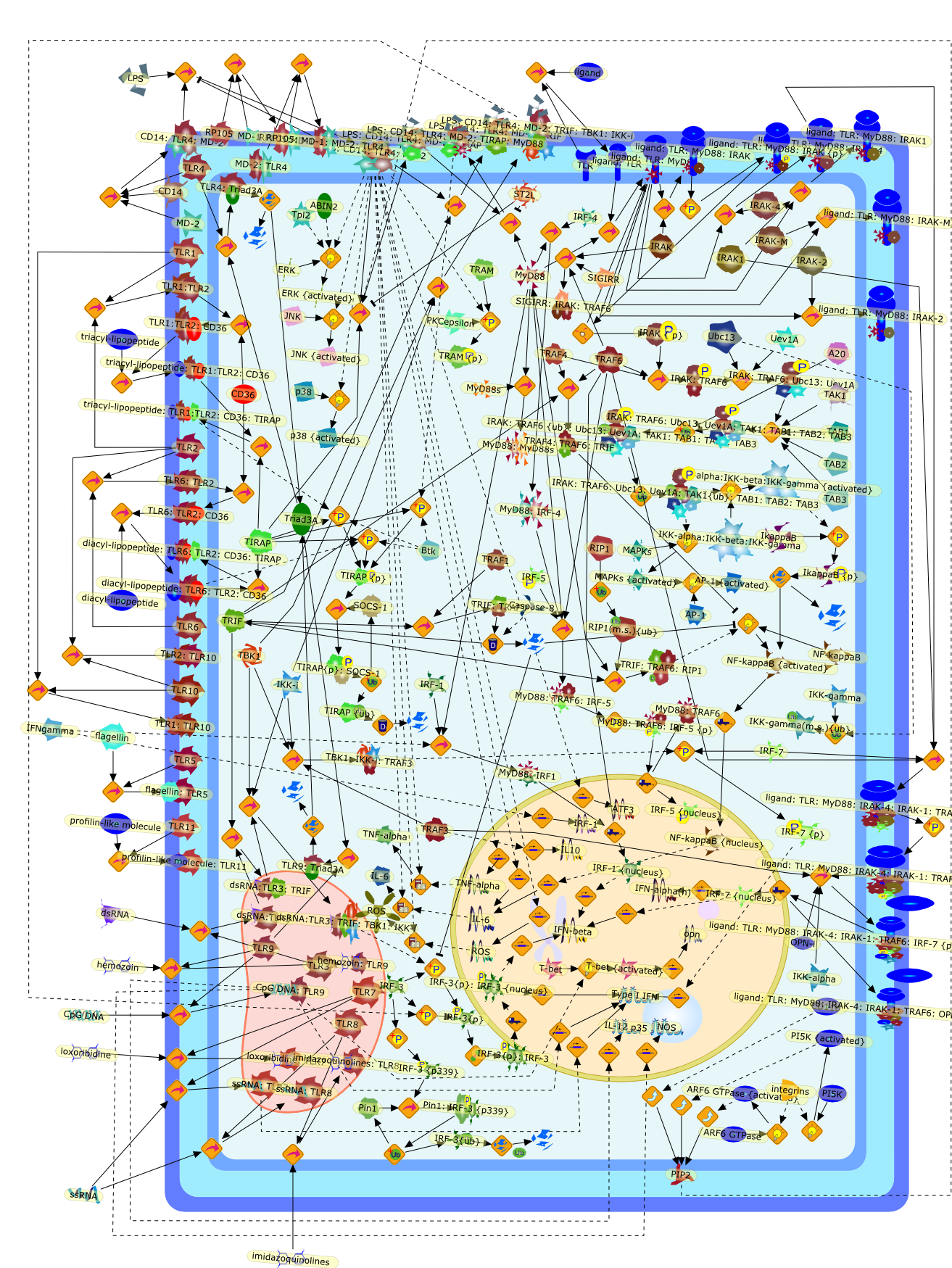{kind=link}
{kind=link}
A20,ABIN2,AP-1,AP-1 {activated},ARF6 GTPase,ARF6 GTPase {activated},ATF3,Btk,CD11b,CD14,CD14: TLR4: MD-2,CD36,Caspase-8,CpG DNA,CpG DNA: TLR9,ERK,ERK {activated},IFN-alpha(h),IFN-beta,IFNgamma,IKK-alpha,IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma {activated},IKK-gamma,IKK-gamma(m.s.){ub},IKK-i,IL-12 p35,IL-6,IL10,IRAK,IRAK { p},IRAK-2,IRAK-4,IRAK-M,IRAK1,IRAK: TRAF6,IRAK: TRAF6 {ub}: Ubc13: Uev1A: TAK1: TAB1: TAB2: TAB3,IRAK: TRAF6: Ubc13: Uev1A,IRAK: TRAF6: Ubc13: Uev1A: TAK1: TAB1: TAB2: TAB3,IRAK: TRAF6: Ubc13: Uev1A: TAK1{ub}: TAB1: TAB2: TAB3,IRF-1,IRF-1 {nucleus},IRF-3,IRF-3 {degraded},IRF-3 {p339},IRF-3{p},IRF-3{p}: IRF-3,IRF-3{p}: IRF-3 {nucleus},IRF-3{ub},IRF-4,IRF-5,IRF-5 {nucleus},IRF-7,IRF-7 {nucleus},IRF-7 {p},IkappaB,IkappaB {p},JNK,JNK {activated},LPS,LPS: CD14: TLR4: MD-2,LPS: CD14: TLR4: MD-2: TIRAP,LPS: CD14: TLR4: MD-2: TIRAP: MyD88,LPS: CD14: TLR4: MD-2: TRIF,LPS: CD14: TLR4: MD-2: TRIF: TBK1: IKK-i,MAPKs,MAPKs {activated},MD-1,MD-2,MD-2: TLR4,MyD88,MyD88: IRF-4,MyD88: IRF1,MyD88: MyD88s,MyD88: TRAF6,MyD88: TRAF6: IRF-5,MyD88: TRAF6: IRF-5 {p},MyD88s,NF-kappaB,NF-kappaB {activated},NF-kappaB {nucleus},NOS,OPN-i,PI5K,PI5K {activated},PIP2,PKCepsilon,Pin1,Pin1: IRF-3 {p339},RIP1,RIP1(m.s.){ub},ROS,RP105,RP105: MD-1: MD-2: TLR4,RP105:MD-1,SIGIRR,SIGIRR: IRAK: TRAF6,SOCS-1,ST2L,T-bet,T-bet {activated},TAB1,TAB2,TAB3,TAK1,TBK1,TBK1: IKK-i: TRAF3,TIRAP,TIRAP {p},TIRAP {ub},TIRAP{p}: SOCS-1,TIRAp {degraded},TLR,TLR1,TLR10,TLR11,TLR1: TLR10,TLR1:TLR2,TLR1:TLR2: CD36,TLR2,TLR2: TLR10,TLR3,TLR4,TLR4 {degraded},TLR4: Triad3A,TLR5,TLR6,TLR6: TLR2,TLR6: TLR2: CD36,TLR7,TLR8,TLR9,TLR9 {degraded},TLR9: Triad3A,TNF-alpha,TRAF1,TRAF1 {degraded},TRAF3,TRAF4,TRAF4: TRAF6: TRIF,TRAF6,TRAM,TRAM {p},TRIF,TRIF: TRAF1,TRIF: TRAF6: RIP1,Tpl2,Triad3A,Type I IFN,Ubc13,Uev1A,degraded IkappaB,diacyl-lipopeptide,diacyl-lipopeptide: TLR6: TLR2: CD36,diacyl-lipopeptide: TLR6: TLR2: CD36: TIRAP,dsRNA,dsRNA:TLR3,dsRNA:TLR3: TRIF,dsRNA:TLR3: TRIF: TBK1: IKK-i,flagellin,flagellin: TLR5,hemozoin,hemozoin: TLR9,imidazoquinolines,imidazoquinolines: TLR8,integrins,ligand,ligand: TLR,ligand: TLR: MyD88,ligand: TLR: MyD88: IRAK,ligand: TLR: MyD88: IRAK {p},ligand: TLR: MyD88: IRAK-2,ligand: TLR: MyD88: IRAK-4,ligand: TLR: MyD88: IRAK-4: IRAK-1: TRAF6: IRF-7,ligand: TLR: MyD88: IRAK-4: IRAK-1: TRAF6: IRF-7 {p},ligand: TLR: MyD88: IRAK-4: IRAK-1: TRAF6: IRF-7 {p}: OPN-i: IKK-alpha: TRAF3,ligand: TLR: MyD88: IRAK-4: IRAK-1: TRAF6: OPN-i: IKK-alpha: TRAF3,ligand: TLR: MyD88: IRAK-M,ligand: TLR: MyD88: IRAK1,loxoribidine,loxoribidine: TLR7,opn,p38,p38 {activated},profilin-like molecule,profilin-like molecule: TLR11,ssRNA,ssRNA: TLR7,ssRNA: TLR8,triacyl-lipopeptide,triacyl-lipopeptide: TLR1:TLR2: CD36,triacyl-lipopeptide: TLR1:TLR2: CD36: TIRAP
The Toll family of receptors consists of cell surface TLRs (TLR4/MD-2, TLR1,TLR2, and TLR6) and intracellular TLRs (TLR3, TLR7, TLR8, and TLR9). Cellsurface TLRs sense microbial membranes such as lipopolysaccharide orlipopeptides. Recognition by TLRs is the frontline where pathogens and a hosttry to take the control of immune responses. Bacteria can modulate the structureof a TLR ligand lipid A to subvert host responses. Cell surface TLRs also senseendogenous ligands which are released in tissue damages as danger signals andinduce inflammation in infectious and non-infectious condition. The availabilityof endogenous ligands and the amount of cell surface TLRs are both tightlylimited to keep TLR responses sufficient for containment of pathogens withoutdetrimental responses to the host.
{kind=link}
{kind=link}
A/D type CpG,A/D type CpG:TLR9,Actin,Actin polymer,B/K type CpG,B/K type CpG: TLR9,Biglycan,CD14,CD14:TLR4:MD-2,CD14:TLR4:MD-2:Biglycan,CD36,Cellular fibronectin,Co-stimulatory molecules,Diacylated lipopeptide,ERK,ERK{active},HMGB1,HMGB1:TLR2:TLR6,Hsp60,Hsp70,IFN-beta,IFNbeta,IL-10,IL-12,IL-12 p40,IRF-3,IRF-3{p},IRF-3{p}:IRF-3{p},LBP,LPS,LPS:LBP,LPS:LBP:CD14,LPS:LBP:CD14:TLR4:MD2,LPS:LBP:CD14:TLR4{oligomer}:MD2,LPS:LBP:CD14:TLR4{oligomer}:MD2:MyD88:TIRAP,LPS:LBP:CD14:TLR4{oligomer}:MD2:TRIF:TRAM,LPS:MD-2,Lipoteichoic acid,MD-1,MD-2,MIP-2,MMP-9,MyD88,NF-kappaB,NF-kappaB{active},NO,OxLDL,Pro-inflammtory cytokies,RP105,RP105:MD-1,RP105:MD-1:TLR4:MD-2,SP-A,TIRAP,TLR2:TLR1,TLR2:TLR1:CD14,TLR2:TLR1:CD14:Di acylated lipopeptide,TLR2:TLR1:CD14:Lipoteichoic acid,TLR2:TLR1:CD36,TLR2:TLR1:CD36:Di acylated lipopeptide,TLR2:TLR1:CD36:Lipoteichoic acid,TLR2:TLR6,TLR2:TLR6:Biglycan,TLR4,TLR4:MD-2:Biglycan,TLR4:MD-2:HMGB1,TLR4:MD2,TLR4:gp96,TLR4;MD-2:mDFbeta,TLR4{Gly},TLR9,TNF-alpha,TRAM,TRIF,Tlr4,Triacylated lipopeptide:TLR2:TLR6,gp96,iNOS,mDF2beta,p38,p38{active},pr0-inflammtory cytokines,pro-inflammatory cytokines,triacylated lipopeptide
The innate immune system provides the host with an immediate and rapid defenseagainst invading microbes. Detection of foreign invaders is mediated by a classof receptors that are known as the pattern recognition receptors, such as thefamily of Toll-like receptors (TLRs). In humans, ten functional TLRs have beenidentified and they respond to conserved pathogen-associated molecular patternsderived from bacteria, mycoplasma, fungi and viruses. TLR activation leads todirect antimicrobial activity against both intracellular and extracellularbacteria, and induces an antiviral gene program. Recently, it was reported thatTLR2 activation leads to the use of vitamin D3 as a mechanism to combatMycobacterium tuberculosis. Here, we focus on recent findings concerning theTLR-induced antimicrobial mechanisms in humans and the therapeutic implicationsof these findings. Owing to their capability to combat a wide array ofpathogens, TLRs are attractive therapeutic targets. However, additionalknowledge about their antimicrobial mechanisms is needed.
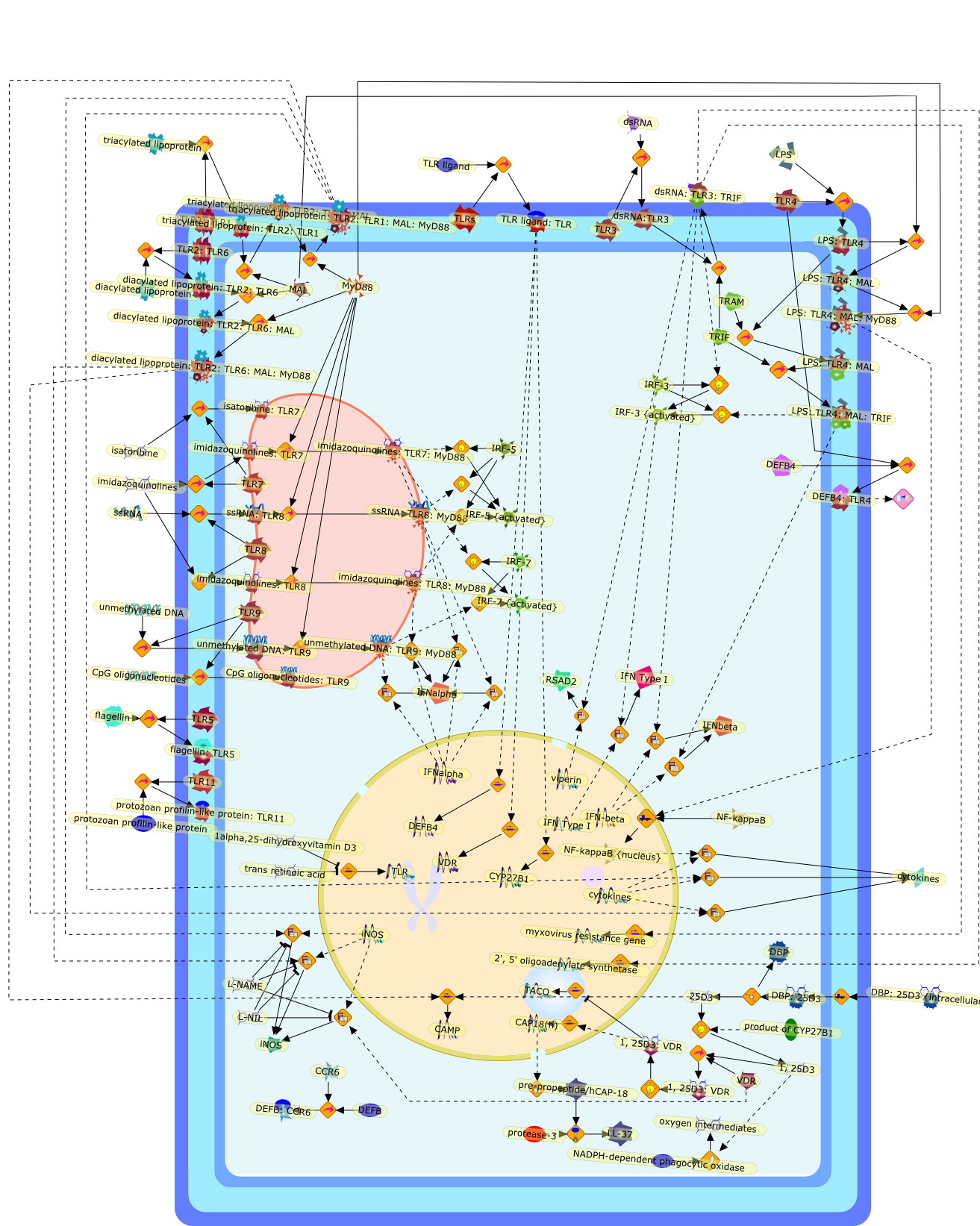{kind=link}
{kind=link}
cytokines,1, 25D3,1, 25D3: VDR,1alpha,25-dihydroxyvitamin D3,2', 5' oligoadenylate synthetase,25D3,CAMP,CAP18(h),CCR6,CYP27B1,CpG oligonucleotides,CpG oligonucleotides: TLR9,DBP,DBP: 25D3,DBP: 25D3 {intracellular},DEFB,DEFB4,DEFB4: TLR4,DEFB: CCR6,IFN Type I,IFN-beta,IFNalpha,IFNbeta,IRF-3,IRF-3 {activated},IRF-5,IRF-5 {activated},IRF-7,IRF-7 {activated},L-NAME,L-NIL,LL-37,LPS,LPS: TLR4,LPS: TLR4: MAL,LPS: TLR4: MAL: MyD88,LPS: TLR4: MAL: TRIF,MAL,MyD88,NADPH-dependent phagocytic oxidase,NF-kappaB,NF-kappaB {nucleus},RSAD2,TACO,TLR,TLR ligand,TLR ligand: TLR,TLR11,TLR2: TLR1,TLR2: TLR6,TLR3,TLR4,TLR5,TLR7,TLR8,TLR9,TLRs,TRAM,TRIF,VDR,cytokines,diacylated lipoprotein,diacylated lipoprotein: TLR2: TLR6,diacylated lipoprotein: TLR2: TLR6: MAL,diacylated lipoprotein: TLR2: TLR6: MAL: MyD88,dsRNA,dsRNA: TLR3: TRIF,dsRNA:TLR3,flagellin,flagellin: TLR5,iNOS,imidazoquinolines,imidazoquinolines: TLR7,imidazoquinolines: TLR7: MyD88,imidazoquinolines: TLR8,imidazoquinolines: TLR8: MyD88,isatoribine,isatoribine: TLR7,myxovirus resistance gene,oxygen intermediates,pre-propeptide/hCAP-18,product of CYP27B1,protease-3,protozoan profilin-like protein,protozoan profilin-like protein: TLR11,ssRNA,ssRNA: TLR8,ssRNA: TLR8: MyD88,trans retinoic acid,triacylated lipoprotein,triacylated lipoprotein: TLR2: TLR1,triacylated lipoprotein: TLR2: TLR1: MAL,triacylated lipoprotein: TLR2: TLR1: MAL: MyD88,unmethylated DNA,unmethylated DNA: TLR9,unmethylated DNA: TLR9: MyD88,viperin
Invasion of viruses and bacteria is initially sensed by the host innate immunesystem, and evokes a rapid inflammatory response. Nucleotides from RNA virusesare recognized by retinoic-acid-inducible gene I-like helicases and Toll-likereceptors, and this recognition triggers signaling cascades that induceantiviral mediators such as type I interferons. By contrast, Toll-like receptorsrecognizing bacterial components induce the expression of proinflammatorycytokines. Furthermore, recent studies suggest that viral and bacterial DNA alsoinduce interferons in a Toll-independent mechanism, possibly throughunidentified cytoplasmic receptor(s).
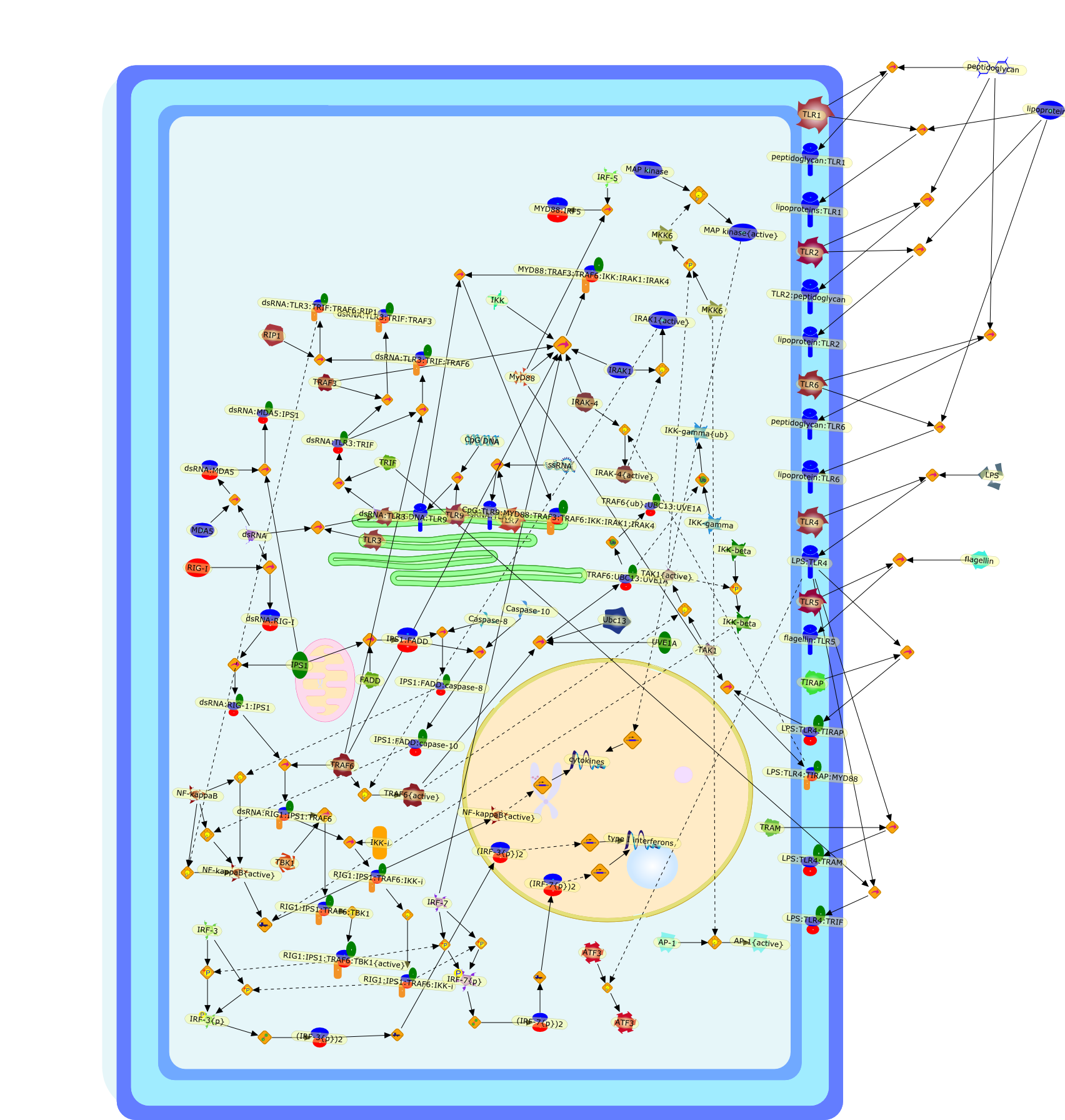{kind=link}
{kind=link}
(IRF-3{p})2,(IRF-7{p})2,AP-1,AP-1{active},ATF3,Caspase-10,Caspase-8,CpG DNA,CpG:TLR9:MYD88:TRAF3:TRAF6:IKK:IRAK1:IRAK4,CpGDNA:TLR9,FADD,IKK,IKK-beta,IKK-gamma,IKK-gamma{ub},IKK-i,IPS1,IPS1:FADD,IPS1:FADD:capase-10,IPS1:FADD:caspase-8,IRAK-4,IRAK-4{active},IRAK1,IRAK1{active},IRF-3,IRF-3{p},IRF-5,IRF-7,IRF-7{p},LPS,LPS:TLR4,LPS:TLR4:TIRAP,LPS:TLR4:TIRAP:MYD88,LPS:TLR4:TRAM,LPS:TLR4:TRIF,MAP kinase,MAP kinase{active},MDA5,MKK6,MYD88:IRF5,MYD88:TRAF3:TRAF6:IKK:IRAK1:IRAK4,MyD88,NF-kappaB,NF-kappaB{active},RIG-I,RIG1:IPS1:TRAF6:IKK-i,RIG1:IPS1:TRAF6:TBK1,RIG1:IPS1:TRAF6:TBK1{active},RIP1,TAK1,TAK1{active},TBK1,TIRAP,TLR1,TLR2,TLR2:peptidoglycan,TLR3,TLR4,TLR5,TLR6,TLR7,TLR9,TRAF3,TRAF6,TRAF6:UBC13:UVE1A,TRAF6{active},TRAF6{ub}:UBC13:UVE1A,TRAM,TRIF,UVE1A,Ubc13,cytokines,dsRNA,dsRNA:MDA5,dsRNA:MDA5:IPS1,dsRNA:RIG-1:IPS1,dsRNA:RIG-I,dsRNA:RIG1:IPS1:TRAF6,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:TRAF3,dsRNA:TLR3:TRIF:TRAF6,dsRNA:TLR3:TRIF:TRAF6:RIP1,flagellin,flagellin:TLR5,lipoprotein:TLR2,lipoprotein:TLR6,lipoproteins,lipoproteins:TLR1,peptidoglycan,peptidoglycan:TLR1,peptidoglycan:TLR6,ssRNA,ssRNA:TLR7,type I interferons
The ability to distinguish foreign nucleic acids from abundant self nucleicacids is essential to protect the host from invading pathogens. Several innateimmune surveillance systems have evolved to detect nucleic acids and triggercellular responses to eliminate foreign invaders. For RNA recognition, theseinclude double stranded (ds)RNA-dependent protein kinase, Toll-like receptor(TLR)3, TLR7, TLR8, retinoic acid-inducible gene (RIG)-I and melanomadifferentiation-associated gene 5. In the case of the nucleic-acid-sensing TLRs,endosomal localization is thought to be crucial for providing self versusnon-self discrimination. For RNA-sensing in the cytoplasm, RIG-I was recentlyshown to detect and directly bind to the 5'-end of certain viral RNA genomes,specifically, to a 5'-triphosphate group. Such 5'-triphosphates are generallyremoved from, or masked on, host RNA species, thereby remaining silent to innateimmunity and providing a structural basis for the distinction between self andnon-self RNA. The mechanisms by which MDA5 senses RNA are unclear at present butseem to involve the sensing of dsRNA structures.
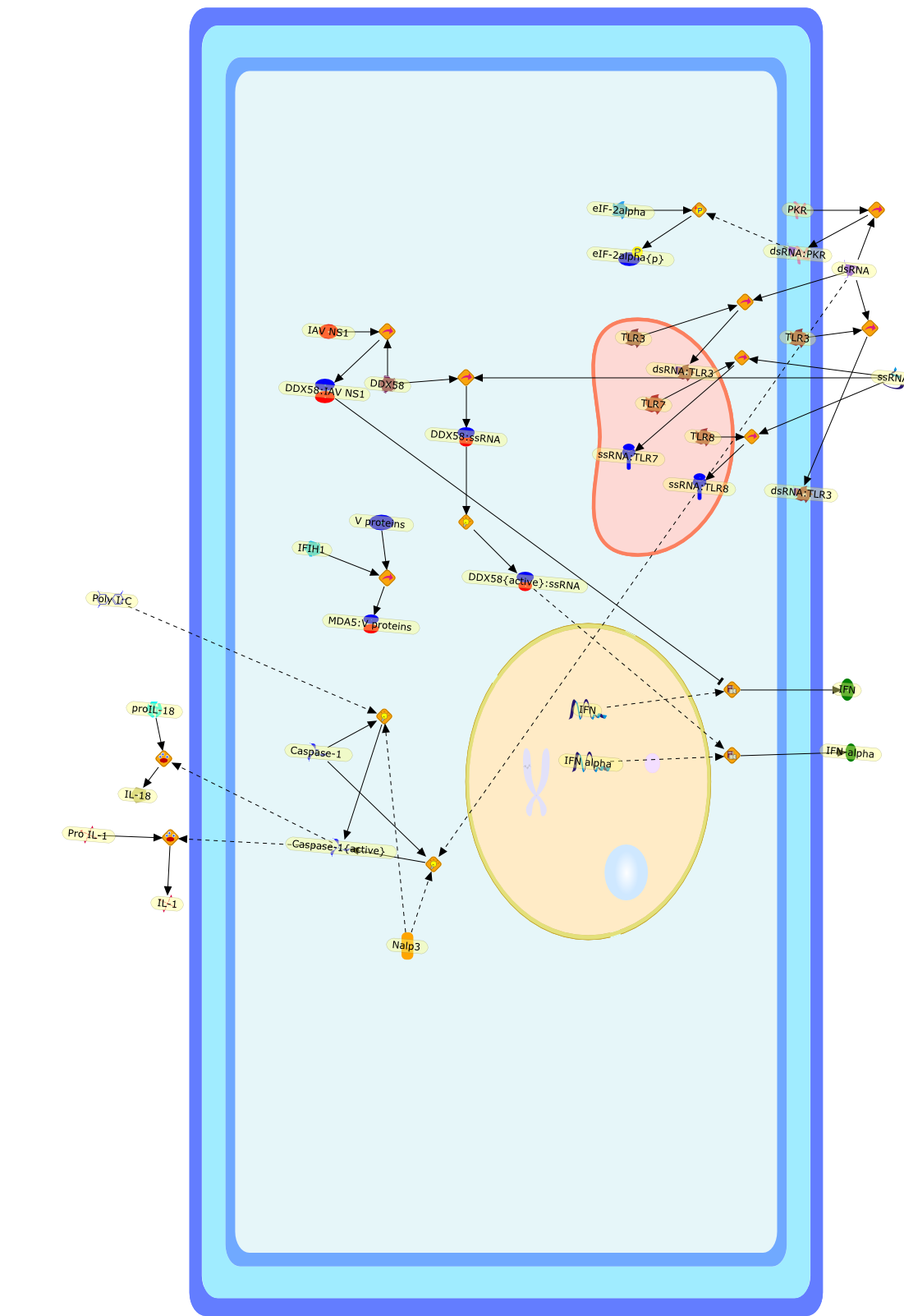{kind=link}
{kind=link}
Caspase-1,Caspase-1{active},DDX58,DDX58:IAV NS1,DDX58:ssRNA,DDX58{active}:ssRNA,IAV NS1,IFIH1,IFN,IFN alpha ,IFN-alpha,IL-1,IL-18,MDA5:V proteins,Nalp3,PKR,Poly I:C,Pro IL-1,TLR3,TLR7,TLR8,V proteins,dsRNA,dsRNA:PKR,dsRNA:TLR3,eIF-2alpha,eIF-2alpha{p},proIL-18,ssRNA,ssRNA:TLR7,ssRNA:TLR8
Toll-like receptors (TLRs) have evolved to recognize conserved features ofmicrobial pathogens, including bacteria, viruses, fungi, and protozoa. Amongthese pathogen classes, viruses are particularly vexing, as they generally lackany uniquely foreign features that easily distinguish them from the host.Consequently, a number of TLR family members have evolved to recognize variousforms of viral nucleic acid. The emergence of this specificity has interestingimplications for how we view the evolution of viral genomes. In this review, Idiscuss the various strategies used by TLRs to recognize viruses as well as theexperimental evidence implicating TLRs in viral immunity.
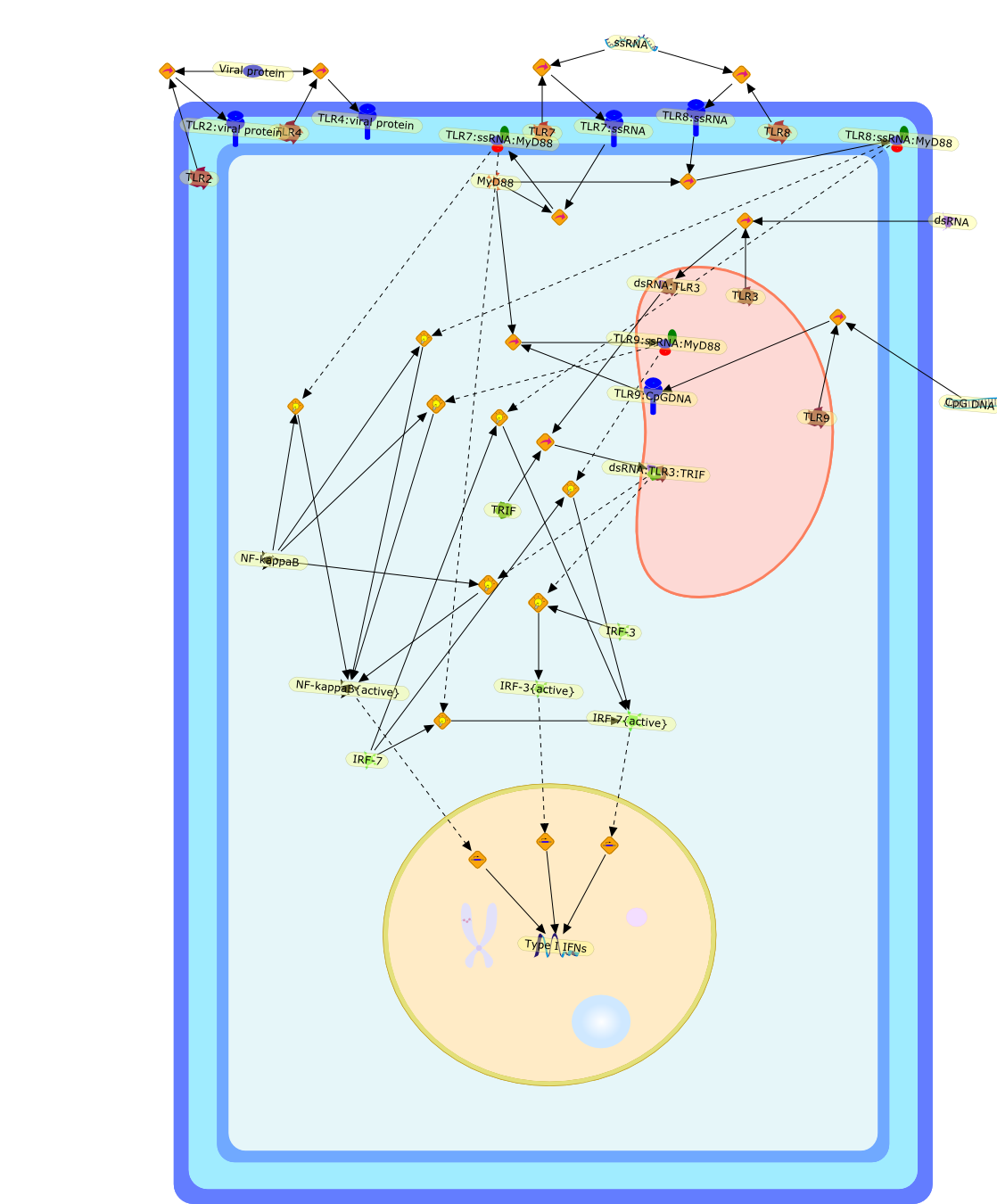{kind=link}
{kind=link}
CpG DNA,IRF-3,IRF-3{active},IRF-7,IRF-7{active},MyD88,NF-kappaB,NF-kappaB{active},TLR2,TLR2:viral protein,TLR3,TLR4,TLR4:viral protein,TLR7,TLR7:ssRNA,TLR7:ssRNA:MyD88,TLR8,TLR8:ssRNA,TLR8:ssRNA:MyD88,TLR9,TLR9:CpGDNA,TLR9:ssRNA:MyD88,TRIF,Type I IFNs,Viral protein,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,ssRNA
Toll-like receptors (TLRs) recognize distinct microbial components to initiatethe innate and adaptive immune responses. TLR activation culminates in theexpression of appropriate pro-inflammatory and immunomodulatory factors to meetpathogenic challenges. The transcription factor NF-kappaB is the masterregulator of all TLR-induced responses and its activation is the pivotal eventin TLR-mediated activation of the innate immune response. Many of the keymolecular events required for TLR-induced NF-kappaB activation have beenelucidated. However, much remains to be learned about the ability of TLRs togenerate pathogen-specific responses using a limited number of transcriptionfactors. This review will focus on our current understanding of NF-kappaBactivation by TLRs and potential mechanisms for achieving a signal-specificresponse through NF-kappaB.
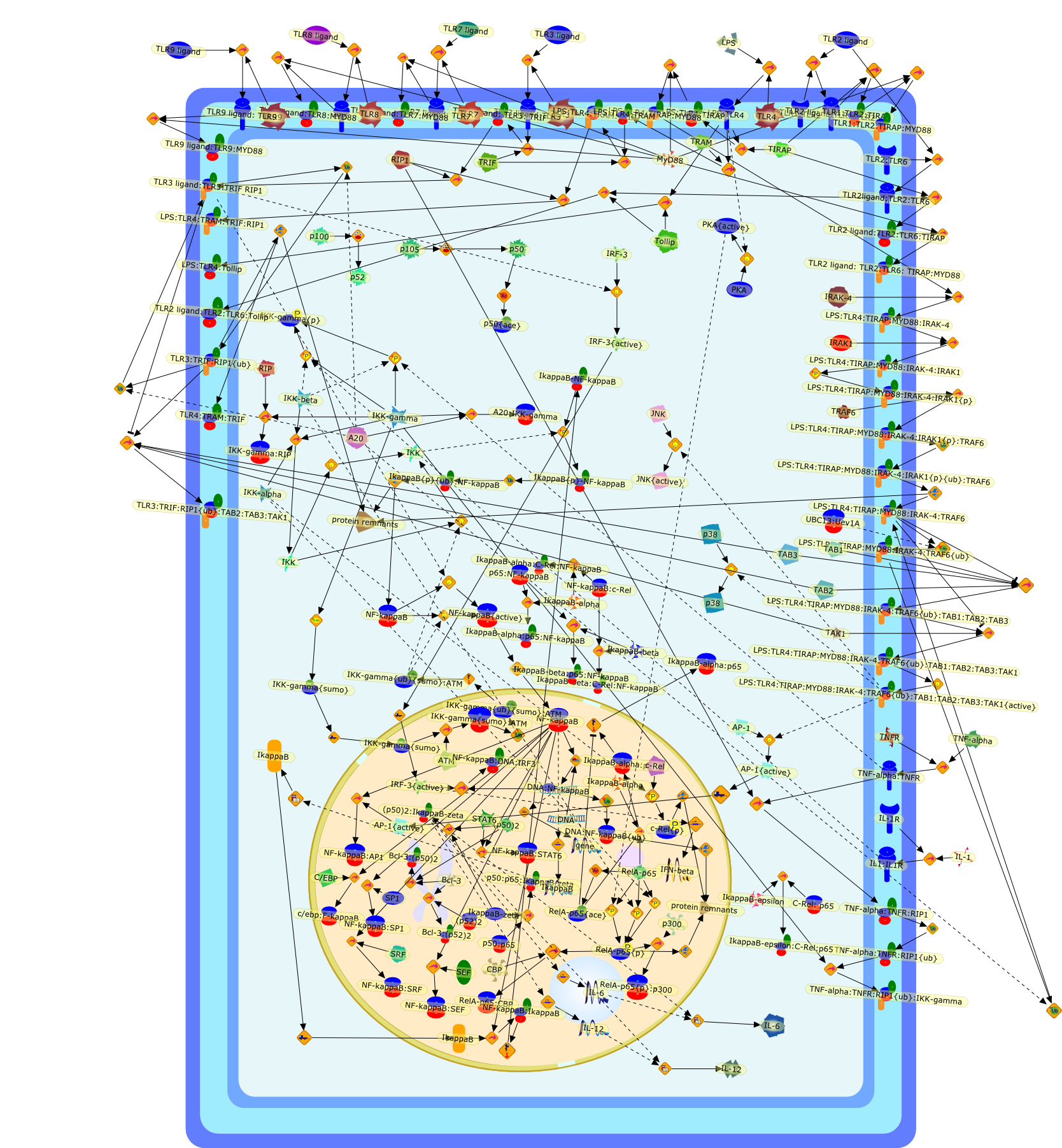{kind=link}
{kind=link}
(p50)2,(p50)2:IkappaB-zeta,(p52)2,A20,A20:IKK-gamma,AP-1,AP-1{active},ATM,Bcl-3,Bcl-3:(p50)2,Bcl-3:(p52)2,C-Rel: p65,C/EBP,CBP,DNA,DNA:NF-kappaB,DNA:NF-kappaB{ub},IFN-beta,IKK,IKK-alpha,IKK-beta,IKK-gamma,IKK-gamma:RIP,IKK-gamma{p},IKK-gamma{sumo},IKK-gamma{sumo}:ATM,IKK-gamma{ub}{sumo}:ATM,IL-1,IL-12,IL-1R,IL-6,IL1:IL1R,IRAK-4,IRAK1,IRF-3,IRF-3{active},IkappaB,IkappaB-NF-kappaB,IkappaB-alpha,IkappaB-alpha:C-Rel:NF-kappaB,IkappaB-alpha:p65,IkappaB-alpha:p65:NF-kappaB,IkappaB-beta,IkappaB-beta:C-Rel:NF-kappaB,IkappaB-beta:p65:NF-kappaB,IkappaB-epsilon,IkappaB-epsilon:C-Rel:p65,IkappaB-zeta,IkappaB{p}-NF-kappaB,IkappaB{p}{ub}:NF-kappaB,JNK,JNK{active},LPS,LPS:TLR4,LPS:TLR4:TIRAP,LPS:TLR4:TIRAP:MYD88,LPS:TLR4:TIRAP:MYD88:IRAK-4,LPS:TLR4:TIRAP:MYD88:IRAK-4:IRAK1,LPS:TLR4:TIRAP:MYD88:IRAK-4:IRAK1{p},LPS:TLR4:TIRAP:MYD88:IRAK-4:IRAK1{p}:TRAF6,LPS:TLR4:TIRAP:MYD88:IRAK-4:IRAK1{p}{ub}:TRAF6,LPS:TLR4:TIRAP:MYD88:IRAK-4:TRAF6,LPS:TLR4:TIRAP:MYD88:IRAK-4:TRAF6{ub},LPS:TLR4:TIRAP:MYD88:IRAK-4:TRAF6{ub}:TAB1:TAB2:TAB3,LPS:TLR4:TIRAP:MYD88:IRAK-4:TRAF6{ub}:TAB1:TAB2:TAB3:TAK1,LPS:TLR4:TIRAP:MYD88:IRAK-4:TRAF6{ub}:TAB1:TAB2:TAB3:TAK1{active},LPS:TLR4:TRAM,LPS:TLR4:TRAM:TRIF,LPS:TLR4:TRAM:TRIF:RIP1,LPS:TLR4:Tollip,MyD88,NF-kappaB,NF-kappaB:AP1,NF-kappaB:DNA:IRF3,NF-kappaB:IkappaB,NF-kappaB:SEF,NF-kappaB:SP1,NF-kappaB:SRF,NF-kappaB:STAT6,NF-kappaB:c-Rel,NF-kappaB{active},PKA,PKA{active},RIP,RIP1,RelA-p65,RelA-p65:CBP,RelA-p65{ace},RelA-p65{p},RelA-p65{p}:p300,SEF,SP1,SRF,STAT6,TAB1,TAB2,TAB3,TAK1,TIRAP,TLR1:TLR2,TLR1:TLR2:TIRAP,TLR1:TLR2:TIRAP:MYD88,TLR2 ligand,TLR2 ligand: TLR2:TLR6: TIRAP:MYD88,TLR2 ligand:TLR1:TLR2,TLR2 ligand:TLR2:TLR6:TIRAP,TLR2 ligand:TLR2:TLR6:Tollip,TLR2:TLR6,TLR2ligand:TLR2:TLR6,TLR3,TLR3 ligand,TLR3 ligand: TLR3: TRIF,TLR3 ligand:TLR3,TLR3 ligand:TLR3:TRIF RIP1,TLR3:TRIF:RIP1{ub},TLR3:TRIF:RIP1{ub}:TAB2:TAB3:TAK1,TLR4,TLR4:TRAM:TRIF,TLR7,TLR7 ligand,TLR7 ligand: TLR7,TLR7 ligand:TLR7:MYD88,TLR8,TLR8 ligand,TLR8 ligand:TLR8:MYD88,TLR9,TLR9 ligand,TLR9 ligand: TLR9,TLR9 ligand:TLR9:MYD88,TNF-alpha,TNF-alpha:TNFR,TNF-alpha:TNFR:RIP1,TNF-alpha:TNFR:RIP1{ub},TNF-alpha:TNFR:RIP1{ub}:IKK-gamma,TNFR,TRAF6,TRAM,TRIF,Tollip,UBC13:Uev1A,c-Rel,c-Rel{p},c/ebp:F-kappaB,e76,gene,p100,p105,p300,p38,p50,p50:p65,p50:p65:IkappaB-zeta,p50{ace},p52,p65:NF-kappaB,protein remnants
Research into intracellular sensing of microbial products is an up and comingfield in innate immunity. Nod1 and Nod2 are members of the rapidly expandingfamily of NACHT domain-containing proteins involved in intracellular recognitionof bacterial products. Nods proteins are involved in the cytosolic detection ofpeptidoglycan motifs of bacteria, recognized through the LRR domain. The role ofthe NACHT-LRR system of detection in innate immune responses is highlighted atthe mucosal barrier, where most of the membranous Toll like receptors (TLRs) arenot expressed, or with pathogens that have devised ways to escape TLR sensing.For a given pathogen, the sum of the pathways induced by the recognition of thedifferent "pathogen associated molecular patterns" (PAMPs) by the differentpattern recognition receptors (PRRs) trigger and shape the subsequent innate andadaptive immune responses. Knowledge gathered during the last decade on PRR andtheir agonists, and recent studies on bacterial infections provide new insightsinto the immune response and the pathogenesis of human infectious diseases.
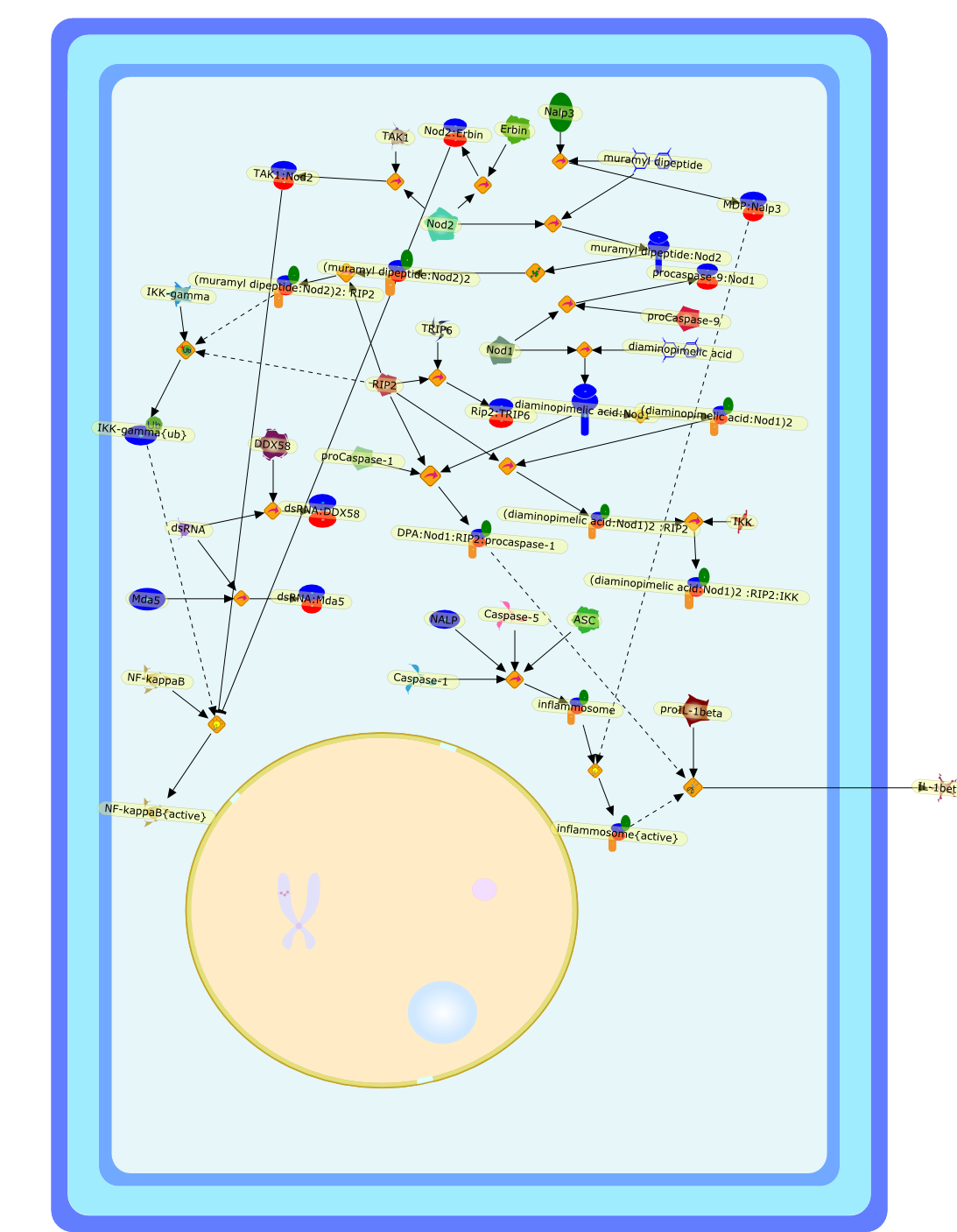{kind=link}
{kind=link}
(diaminopimelic acid:Nod1)2 , (diaminopimelic acid:Nod1)2 :RIP2, (diaminopimelic acid:Nod1)2 :RIP2:IKK,(muramyl dipeptide:Nod2)2,(muramyl dipeptide:Nod2)2: RIP2,ASC,Caspase-1,Caspase-5,DDX58,DPA:Nod1:RIP2:procaspase-1,Erbin,IKK,IKK-gamma,IKK-gamma{ub},IL-1beta,MDP:Nalp3,Mda5,NALP,NF-kappaB,NF-kappaB{active},Nalp3,Nod1,Nod2,Nod2:Erbin,RIP2,Rip2:TRIP6,TAK1,TAK1:Nod2,TRIP6,diaminopimelic acid ,diaminopimelic acid:Nod1,dsRNA,dsRNA:DDX58,dsRNA:Mda5,inflammosome,inflammosome{active},muramyl dipeptide,muramyl dipeptide:Nod2,proCaspase-1,proCaspase-9,proIL-1beta,procaspase-9:Nod1
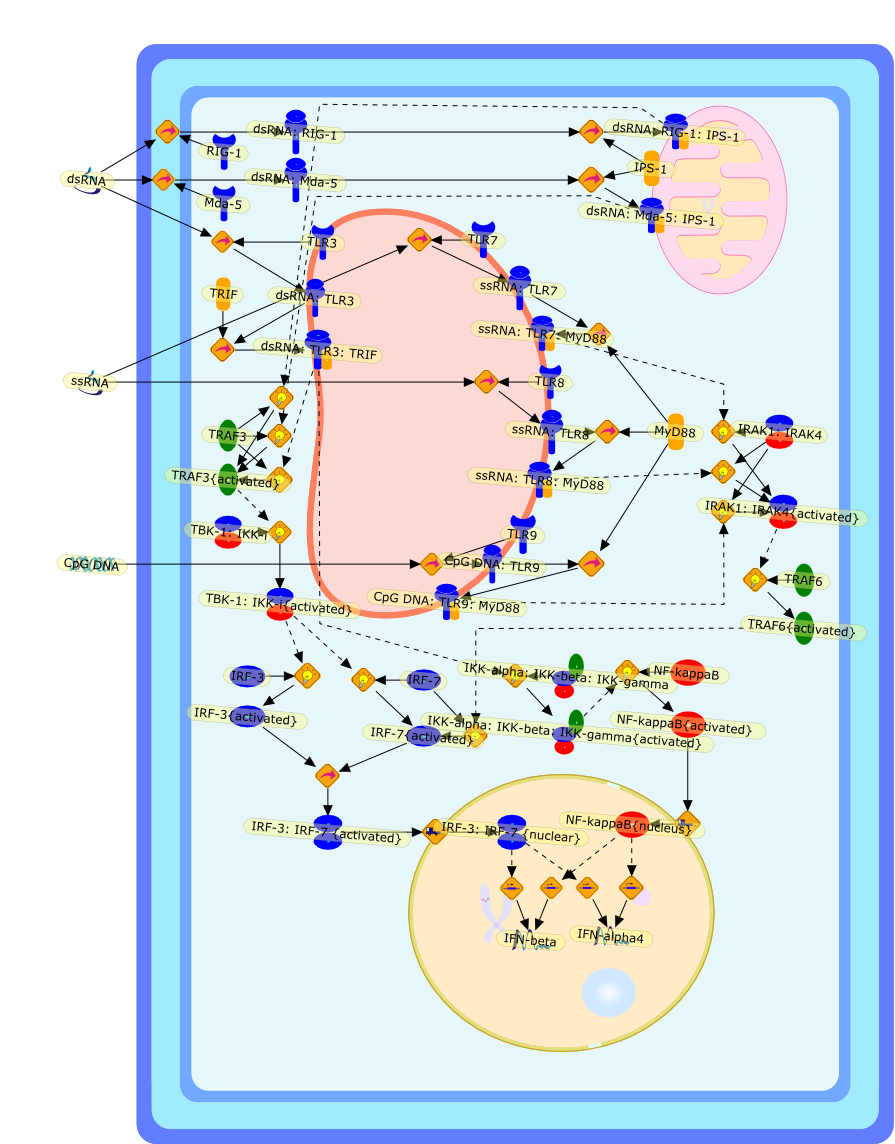{kind=link}
{kind=link}
CpG DNA,CpG DNA: TLR9,CpG DNA: TLR9: MyD88,IFN-alpha4,IFN-beta,IKK-alpha: IKK-beta: IKK-gamma,IKK-alpha: IKK-beta: IKK-gamma{activated},IPS-1,IRAK1: IRAK4,IRAK1: IRAK4{activated},IRF-3,IRF-3: IRF-7 {activated},IRF-3: IRF-7 {nuclear},IRF-3{activated},IRF-7,IRF-7{activated},Mda-5,MyD88,NF-kappaB,NF-kappaB{activated},NF-kappaB{nucleus},RIG-1,TBK-1: IKK-i,TBK-1: IKK-i{activated},TLR3,TLR7,TLR8,TLR9,TRAF3,TRAF3{activated},TRAF6,TRAF6{activated},TRIF,dsRNA,dsRNA: Mda-5,dsRNA: Mda-5: IPS-1,dsRNA: RIG-1,dsRNA: RIG-1: IPS-1,dsRNA: TLR3,dsRNA: TLR3: TRIF,ssRNA,ssRNA: TLR7,ssRNA: TLR7: MyD88,ssRNA: TLR8,ssRNA: TLR8: MyD88
Toll-like receptors (TLRs) are key molecules of the innate immune systems, whichdetect conserved structures found in a broad range of pathogens and triggerinnate immune responses. A subset of TLRs recognizes viral components andinduces antiviral responses. Whereas TLR4 recognizes viral components at thecell surface, TLR3, TLR7, TLR8, and TLR9 recognize viral nucleic acids onendosomal membrane. After ligand recognition, these members activate theirintrinsic signaling pathways and induce type I interferon. In this review, wediscuss the recent findings of the viral recognition by TLRs and their signalingpathways.
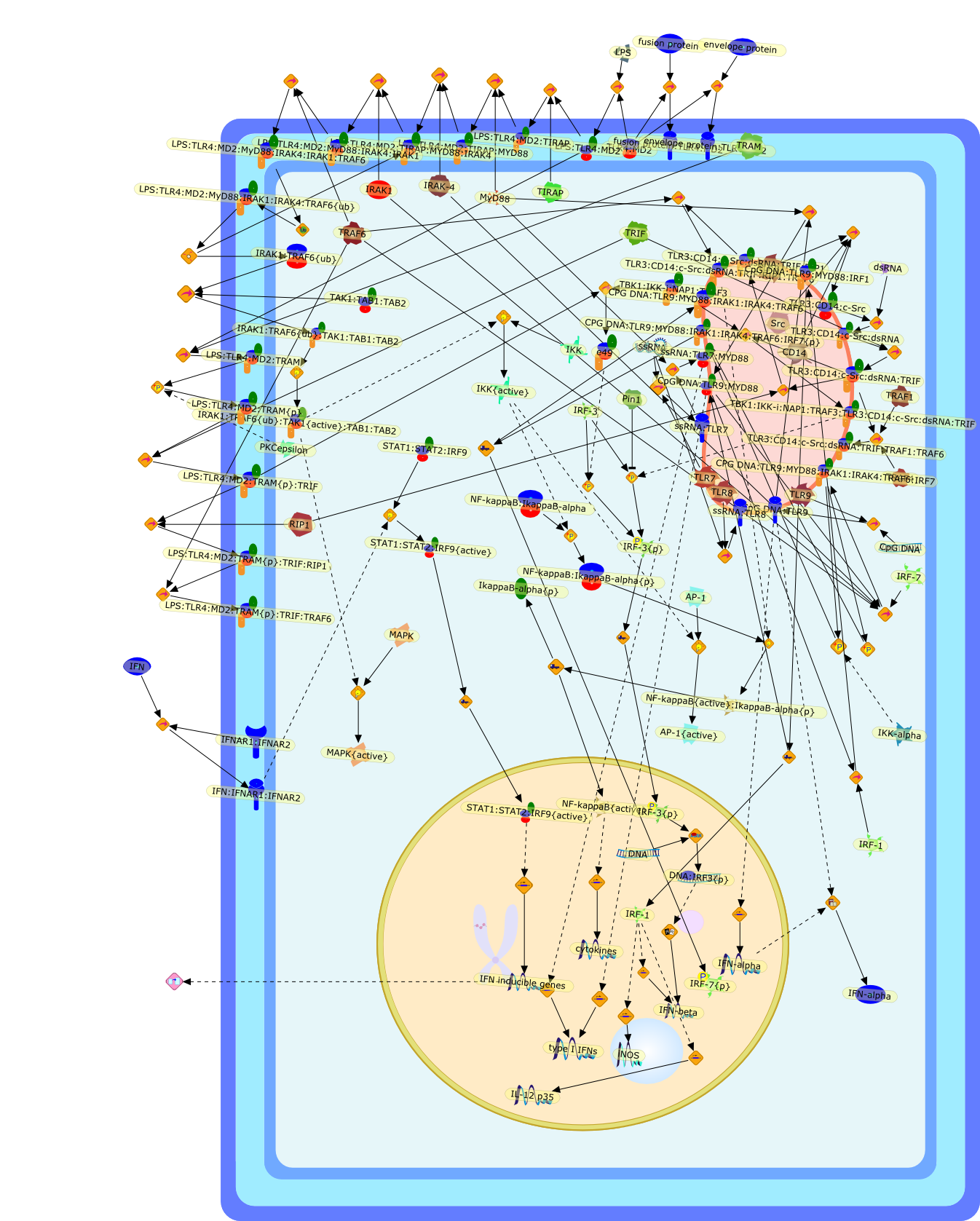{kind=link}
{kind=link}
AP-1,AP-1{active},CD14,CPG DNA:TLR9:MYD88:IRAK1:IRAK4:TRAF6,CPG DNA:TLR9:MYD88:IRAK1:IRAK4:TRAF6:IRF7,CPG DNA:TLR9:MYD88:IRAK1:IRAK4:TRAF6:IRF7{p},CpG DNA,CpG DNA:TLR9,CpG DNA:TLR9:MYD88,CpG DNA:TLR9:MYD88:IRF1,DNA,DNA:IRF3{p},IFN,IFN inducible genes,IFN-alpha,IFN-beta,IFN:IFNAR1:IFNAR2,IFNAR1:IFNAR2,IKK,IKK-alpha,IKK{active},IL-12 p35,IRAK-4,IRAK1,IRAK1:TRAF6{ub},IRAK1:TRAF6{ub}:TAK1:TAB1:TAB2,IRAK1:TRAF6{ub}:TAK1{active}:TAB1:TAB2,IRF-1,IRF-3,IRF-3{p},IRF-7,IRF-7{p},IkappaB-alpha{p},LPS,LPS:TLR4:MD2,LPS:TLR4:MD2:MyD88:IRAK1:IRAK4:TRAF6{ub},LPS:TLR4:MD2:MyD88:IRAK4:IRAK1,LPS:TLR4:MD2:MyD88:IRAK4:IRAK1:TRAF6,LPS:TLR4:MD2:TIRAP,LPS:TLR4:MD2:TIRAP:MYD88,LPS:TLR4:MD2:TIRAP:MYD88:IRAK4,LPS:TLR4:MD2:TRAM,LPS:TLR4:MD2:TRAM{p},LPS:TLR4:MD2:TRAM{p}:TRIF,LPS:TLR4:MD2:TRAM{p}:TRIF:RIP1,LPS:TLR4:MD2:TRAM{p}:TRIF:TRAF6,MAPK,MAPK{active},MyD88,NF-kappaB:IkappaB-alpha,NF-kappaB:IkappaB-alpha{p},NF-kappaB{active},NF-kappaB{active}:IkappaB-alpha{p},PKCepsilon,Pin1,RIP1,STAT1:STAT2:IRF9,STAT1:STAT2:IRF9{active},Src,TAK1:TAB1:TAB2,TBK1:IKK-i:NAP1:TRAF3,TBK1:IKK-i:NAP1:TRAF3:TLR3:CD14:c-Src:dsRNA:TRIF,TIRAP,TLR3,TLR3:CD14:c-Src,TLR3:CD14:c-Src:dsRNA,TLR3:CD14:c-Src:dsRNA:TRIF,TLR3:CD14:c-Src:dsRNA:TRIF:RIP1,TLR3:CD14:c-Src:dsRNA:TRIF:RIP1:TRAF6,TLR3:CD14:c-Src:dsRNA:TRIF:TRAF1:TRAF6,TLR4:MD2,TLR7,TLR8,TLR9,TRAF1,TRAF4,TRAF6,TRAM,TRIF,cytokines,dsRNA,e49,envelope protein,envelope protein:TLR4:MD2,fusion protein,fusion protein:TLR4:MD2,iNOS,ssRNA,ssRNA:TLR7,ssRNA:TLR7:MYD88,ssRNA:TLR8,type I IFNs
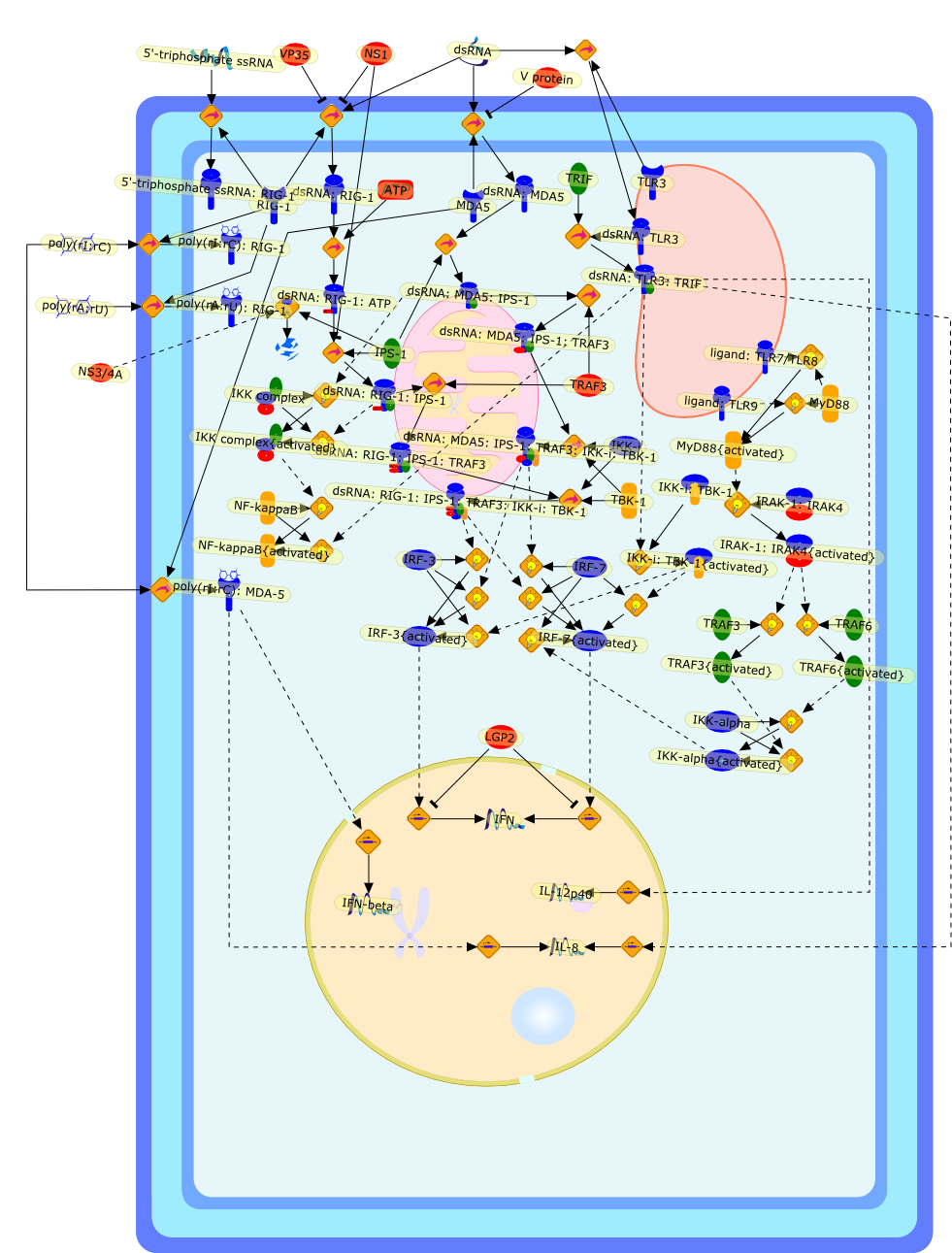{kind=link}
{kind=link}
5'-triphosphate ssRNA,5'-triphosphate ssRNA: RIG-1,ATP,IFN,IFN-beta,IKK complex,IKK complex{activated},IKK-alpha,IKK-alpha{activated},IKK-i,IKK-i: TBK-1,IKK-i: TBK-1{activated},IL-12p40,IL-8,IPS-1,IRAK-1: IRAK4,IRAK-1: IRAK4{activated},IRF-3,IRF-3{activated},IRF-7,IRF-7{activated},LGP2,MDA5,MyD88,MyD88{activated},NF-kappaB,NF-kappaB{activated},NS1,NS3/4A,RIG-1,TBK-1,TLR3,TRAF3,TRAF3{activated},TRAF6,TRAF6{activated},TRIF,V protein,VP35,dsRNA,dsRNA: MDA5,dsRNA: MDA5: IPS-1,dsRNA: MDA5: IPS-1: TRAF3,dsRNA: MDA5: IPS-1: TRAF3: IKK-i: TBK-1,dsRNA: RIG-1,dsRNA: RIG-1: ATP,dsRNA: RIG-1: IPS-1,dsRNA: RIG-1: IPS-1: TRAF3,dsRNA: RIG-1: IPS-1: TRAF3: IKK-i: TBK-1,dsRNA: TLR3,dsRNA: TLR3: TRIF,ligand: TLR7/TLR8,ligand: TLR9,poly(rA:rU),poly(rA:rU): RIG-1,poly(rI:rC),poly(rI:rC): MDA-5,poly(rI:rC): RIG-1,protein remnant
Rapid induction of type I interferon (IFN) expression is a central event in theestablishment of the innate immune response against viral infection and requiresthe activation of multiple transcriptional proteins following engagement andsignaling through Toll-like receptor-dependent and -independent pathways. Thetranscription factor interferon regulatory factor-3 (IRF-3) contributes to afirst line of defense against viral infection by inducing the production ofIFN-beta that in turn amplifies the IFN response and the development ofantiviral activity. In murine knock-out models, the absence of IRF-3 and theclosely related IRF-7 ablates IFN production and increases viral pathogenesis,thus supporting a pivotal role for IRF-3/IRF-7 in the development of the hostantiviral response.
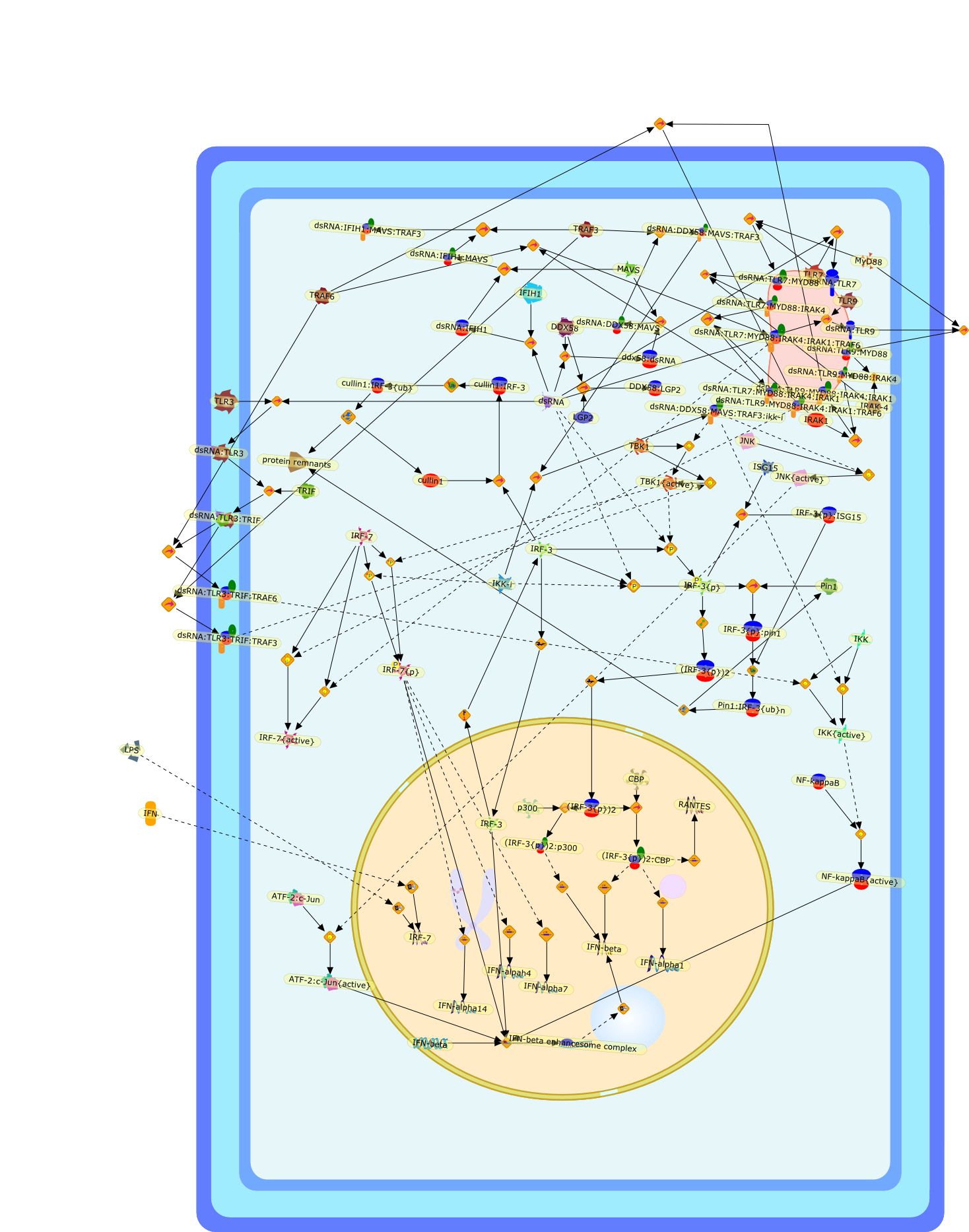{kind=link}
{kind=link}
(IRF-3{p})2,(IRF-3{p})2:CBP,(IRF-3{p})2:p300,ATF-2:c-Jun,ATF-2:c-Jun{active},CBP,DDX58,DDX58:LGP2,IFIH1,IFN,IFN-alpah4,IFN-alpha1,IFN-alpha14,IFN-alpha7,IFN-beta,IFN-beta enhancesome complex,IKK,IKK-i,IKK{active},IRAK-4,IRAK1,IRF-3,IRF-3{p},IRF-3{p}:ISG15,IRF-3{p}:pin1,IRF-7,IRF-7{active},IRF-7{p},ISG15,JNK,JNK{active},LGP2,LPS,MAVS,MyD88,NF-kappaB,NF-kappaB{active},Pin1,Pin1:IRF-3{ub}n,RANTES,TBK1,TBK1{active},TLR3,TLR7,TLR9,TRAF3,TRAF6,TRIF,cullin1,cullin1:IRF-3,cullin1:IRF-3{ub},ddx58:dsRNA,dsRNA,dsRNA:DDX58:MAVS,dsRNA:DDX58:MAVS:TRAF3,dsRNA:DDX58:MAVS:TRAF3:ikk-i,dsRNA:IFIH1,dsRNA:IFIH1:MAVS,dsRNA:IFIH1:MAVS:TRAF3,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:TRAF3,dsRNA:TLR3:TRIF:TRAF6,dsRNA:TLR7,dsRNA:TLR7:MYD88,dsRNA:TLR7:MYD88:IRAK4,dsRNA:TLR7:MYD88:IRAK4:IRAK1,dsRNA:TLR7:MYD88:IRAK4:IRAK1:TRAF6,dsRNA:TLR9,dsRNA:TLR9:MYD88,dsRNA:TLR9:MYD88:IRAK4,dsRNA:TLR9:MYD88:IRAK4:IRAK1,dsRNA:TLR9:MYD88:IRAK4:IRAK1:TRAF6,p300,protein remnants
The term sepsis describes a potentially lethal clinical condition that developsas a result of a dysregulated host response to bacterial infection. The mostcommon bacterial component implicated in initiating the septic syndrome is acell wall molecule derived from Gram-negative bacteria, known aslipopolysaccharide (LPS) or endotoxin. Like all mammals, humans are equippedwith an LPS-sensing machinery consisting, primarily, of LPS-binding protein(LBP), CD14, a glycosylphosphatidylinositol (GPI)-anchored monocytedifferentiation antigen, and toll-like receptor 4 (TLR4), a signal-transducingintegral membrane protein. Modest stimulation of TLR4 facilitates theelimination of invading microorganisms. Potent TLR4 stimulation, however,produces severe reactions in the host, often leading to multiple organ failureand death. The search for pharmaceuticals that reduce mortality in septicpatients has been a painstaking process. Thus far, only a few compounds havebeen found to significantly reduce mortality rates. Perhaps one of the morepromising therapeutic strategies currently pursued is based on theidentification of synthetic or naturally occurring substances that neutralizeLPS or inhibit LPS-mediated activation of host immune cells, such as monocytesand macrophages. Here, we describe a number of diverse molecular structures witha capacity to either enhance or blunt LPS-induced monocyte activation. Theunderlying molecular mechanisms are discussed.
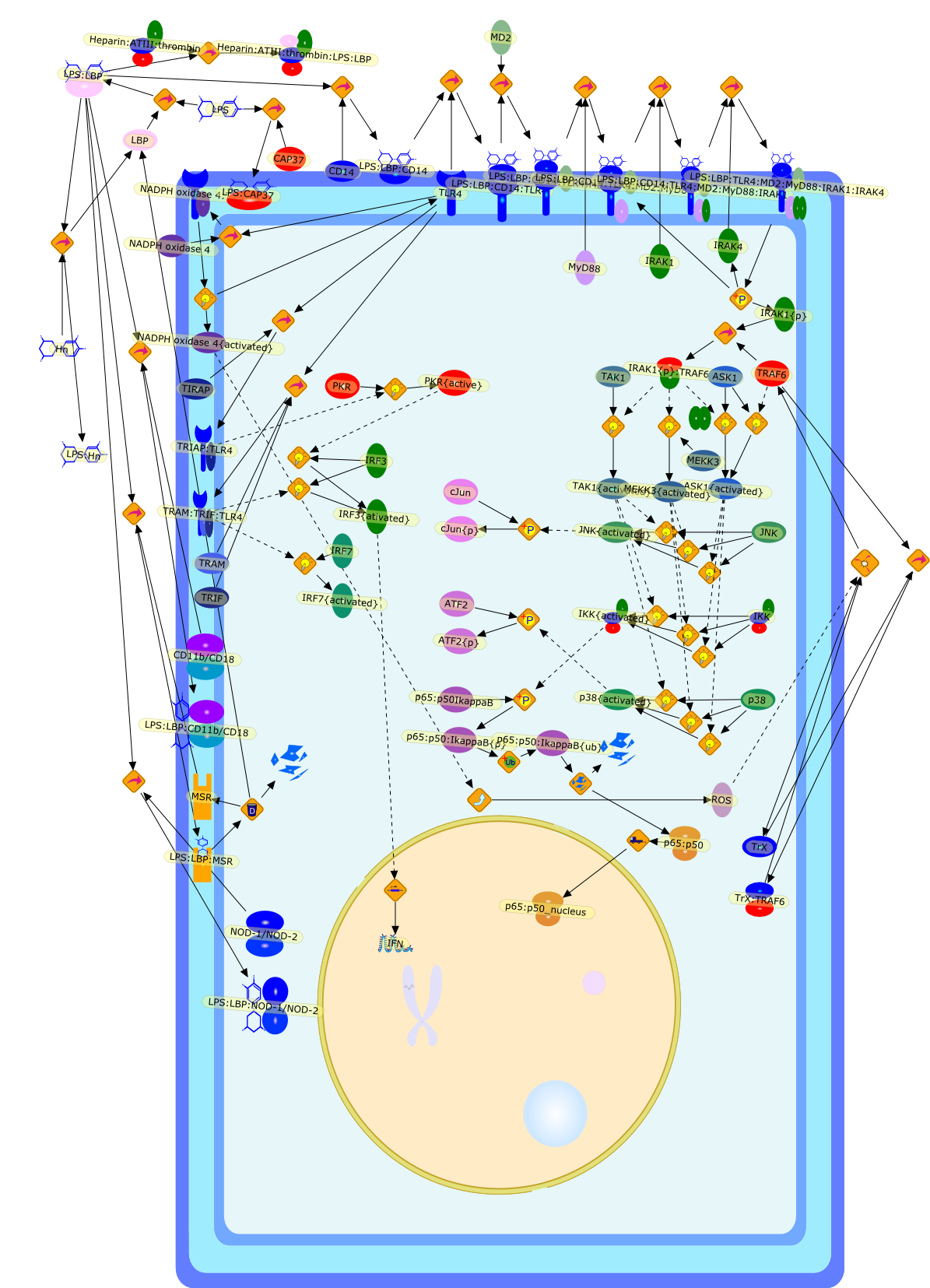{kind=link}
{kind=link}
ASK1,ASK1{activated},ATF2,ATF2{p},CAP37,CD11b/CD18,CD14,Heparin:ATIII:thrombin,Heparin:ATIII:thrombin:LPS:LBP,Hn,IFN,IKK,IKK{activated},IRAK1,IRAK1{p},IRAK1{p}:TRAF6,IRAK4,IRF3,IRF3{ativated},IRF7,IRF7{activated},IkappaB{degraded},JNK,JNK{activated},LBP,LPS,LPS:CAP37,LPS:Hn,LPS:LBP,LPS:LBP:CD11b/CD18,LPS:LBP:CD14,LPS:LBP:CD14:TLR4,LPS:LBP:CD14:TLR4:MD2,LPS:LBP:CD14:TLR4:MD2:MyD88,LPS:LBP:CD14:TLR4:MD2:MyD88:IRAK1,LPS:LBP:MSR,LPS:LBP:NOD-1/NOD-2,LPS:LBP:TLR4:MD2:MyD88:IRAK1:IRAK4,LPS{degraded},MD2,MEKK3,MEKK3{activated},MSR,MyD88,NADPH oxidase 4,NADPH oxidase 4:TLR4,NADPH oxidase 4{activated},NOD-1/NOD-2,PKR,PKR{active},ROS,TAK1,TAK1{activated},TIRAP,TLR4,TRAF6,TRAM,TRAM:TRIF:TLR4,TRIAP:TLR4,TRIF,TrX,TrX:TRAF6,cJun,cJun{p},p38,p38{activated},p65:p50,p65:p50:IkappaB{p},p65:p50:IkappaB{ub},p65:p50IkappaB,p65:p50_nucleus
Signaling by two of the most important bacteria-sensing TLRs, TLR2 and TLR4,involves two adaptor proteins, MyD88 adaptor-like (Mal) and Toll/IL-1 receptor(TIR) domain-containing adaptor-inducing IFN-beta (Trif)-related adaptormolecule (TRAM). Recently, new insights into the functioning of these twoadapters have emerged. Mal is required by both TLRs to act as a bridge torecruit the adaptor MyD88, leading ultimately to NF-kappaB activation.Similarly, TRAM acts as a bridge to recruit TRIF to the TLR4 complex, leading toactivation of the transcription factor IFN regulatory factor 3. Consistent withMal and TRAM being key points of control, recent evidence suggests that they aresubject to regulation by phosphorylation. Further, a variant in Mal in humanshas been found to protect against multiple infectious diseases. Finally, anotherTIR domain-containing adaptor, sterile alpha and HEAT/armadillo motif protein(SARM), has been shown to act as an inhibitor of TRIF-dependent signaling. Theserecent discoveries add to the complexity of TLR signaling and highlight specificcontrol mechanisms for TLR2 and TLR4 signaling.
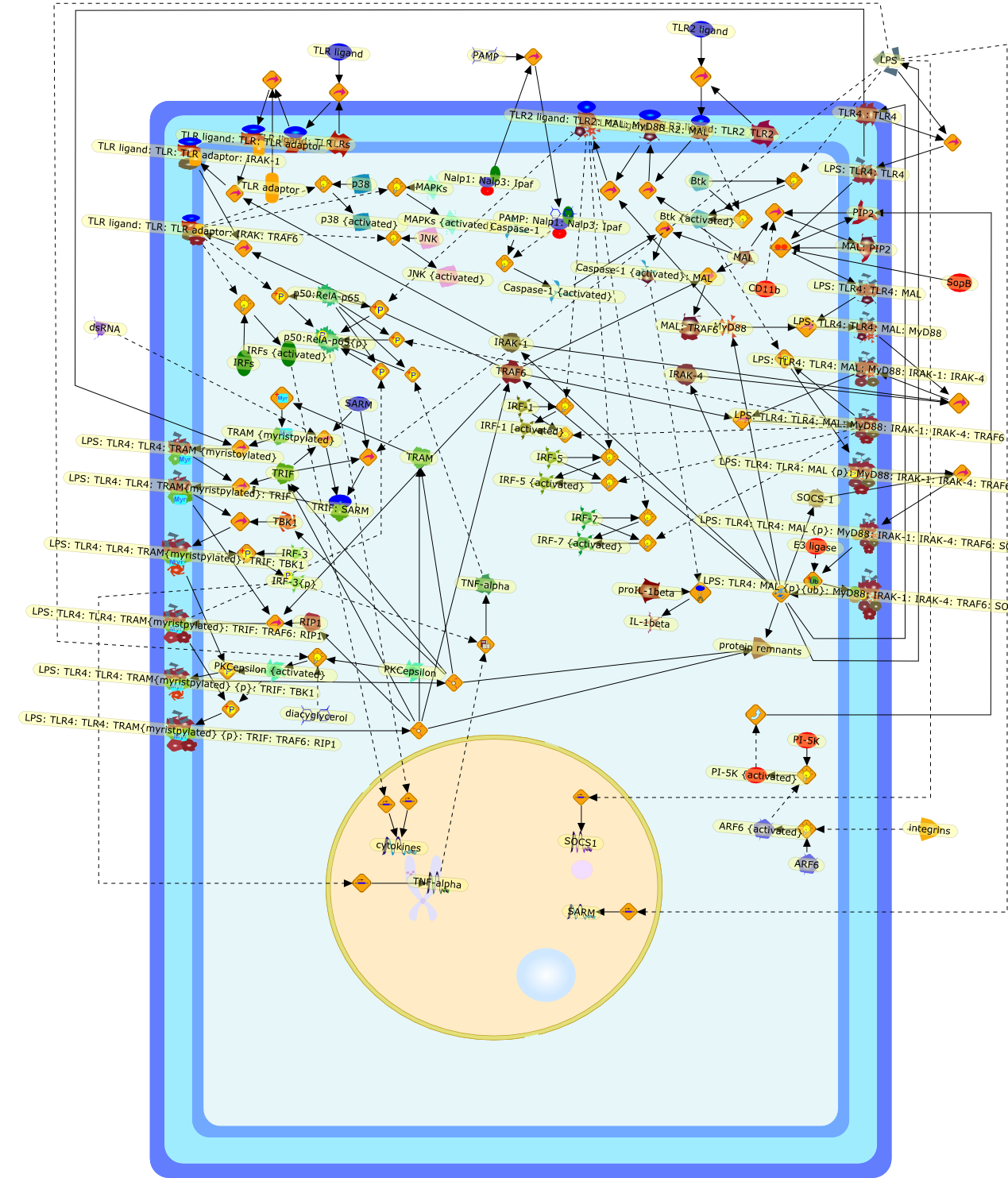{kind=link}
{kind=link}
ARF6,ARF6 {activated},Btk,Btk {activated},CD11b,Caspase-1,Caspase-1 {activated},Caspase-1 {activated}: MAL,E3 ligase,IL-1beta,IRAK-1,IRAK-4,IRF-1,IRF-1 [activated},IRF-3,IRF-3{p},IRF-5,IRF-5 {activated},IRF-7,IRF-7 {activated},IRFs,IRFs {activated},JNK,JNK {activated},LPS,LPS: TLR4: MAL {p}{ub}: MyD88: IRAK-1: IRAK-4: TRAF6: SOCS-1,LPS: TLR4: TLR4,LPS: TLR4: TLR4: MAL,LPS: TLR4: TLR4: MAL {p}: MyD88: IRAK-1: IRAK-4: TRAF6,LPS: TLR4: TLR4: MAL {p}: MyD88: IRAK-1: IRAK-4: TRAF6: SOCS-1,LPS: TLR4: TLR4: MAL: MyD88,LPS: TLR4: TLR4: MAL: MyD88: IRAK-1: IRAK-4,LPS: TLR4: TLR4: MAL: MyD88: IRAK-1: IRAK-4: TRAF6,LPS: TLR4: TLR4: TRAM {myristoylated},LPS: TLR4: TLR4: TRAM{myristpylated} {p}: TRIF: TBK1,LPS: TLR4: TLR4: TRAM{myristpylated} {p}: TRIF: TRAF6: RIP1,LPS: TLR4: TLR4: TRAM{myristpylated}: TRIF,LPS: TLR4: TLR4: TRAM{myristpylated}: TRIF: TBK1,LPS: TLR4: TLR4: TRAM{myristpylated}: TRIF: TRAF6: RIP1,MAL,MAL: PIP2,MAL: TRAF6,MAPKs,MAPKs {activated},MyD88,Nalp1: Nalp3: Ipaf,PAMP,PAMP: Nalp1: Nalp3: Ipaf,PI-5K,PI-5K {activated},PIP2,PKCepsilon,PKCepsilon {activated},RIP1,SARM,SOCS-1,SOCS1,SopB,TBK1,TLR adaptor,TLR ligand,TLR ligand: TLR,TLR ligand: TLR: TLR adaptor,TLR ligand: TLR: TLR adaptor: IRAK-1,TLR ligand: TLR: TLR adaptor: IRAK: TRAF6,TLR2,TLR2 ligand,TLR2 ligand: TLR2,TLR2 ligand: TLR2: MAL,TLR2 ligand: TLR2: MAL: MyD88,TLR4 : TLR4,TLRs,TNF-alpha,TRAF6,TRAM,TRAM {myristpylated},TRIF,TRIF: SARM,cytokines,diacyglycerol,dsRNA,integrins,p38,p38 {activated},p50:RelA-p65,p50:RelA-p65{p},proIL-1beta,protein remnants
G-protein-coupled receptors (GPCRs) are widely targeted in drug discovery. Asmacrophages are key cellular mediators of acute and chronic inflammation, wereview here the role of GPCRs in regulating macrophage function, with a focus oncontribution to disease pathology and potential therapeutic applications. Withinthis analysis, we highlight novel GPCRs with a macrophage-restricted expressionprofile, which provide avenues for further exploration. We also review anemerging literature, which documents novel roles for GPCR signaling componentsin GPCR-independent signaling in macrophages. In particular, we examine thecrosstalk between GPCR and TLR signaling pathways and highlight GPCR signalingmolecules which are likely to have uncharacterized functions in this celllineage.
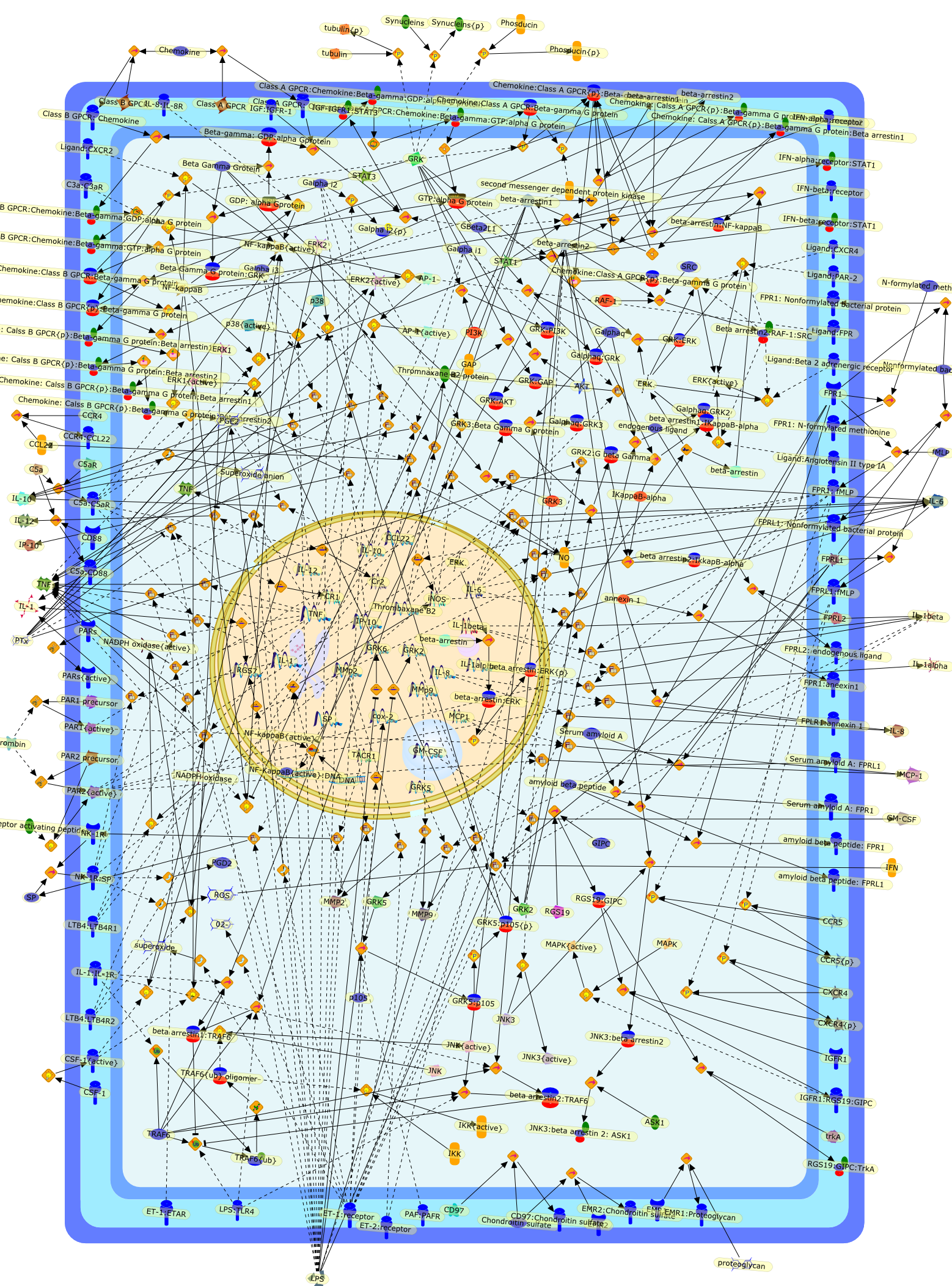{kind=link}
{kind=link}
02-,AKT,AP-1,AP-1{active},ASK1,Beta Gamma G protein:GRK,Beta Gamma Grotein,Beta arrestin2:RAF-1:SRC,Beta-gamma: GDP:alpha Gprotein,C3a:C3aR,C5a,C5a:C5aR,C5a:CD88,C5aR,CCL22,CCR4,CCR4:CCL22,CCR5,CCR5{p},CD88,CD97,CD97:Chondroitin sulfate,CR1,CSF-1,CSF-1{active},CXCR4,CXCR4{p},Chemokine,Chemokine: Calss A GPCR{p}:Beta-gamma G protein:Beta arrestin1,Chemokine: Calss A GPCR{p}:Beta-gamma G protein:Beta arrestin2,Chemokine: Calss B GPCR{p}:Beta-gamma G protein:Beta arrestin1,Chemokine: Calss B GPCR{p}:Beta-gamma G protein:Beta arrestin2,Chemokine:Class A GPCR:Beta-gamma G protein,Chemokine:Class A GPCR{p}:Beta-gamma G protein,Chemokine:Class B GPCR:Beta-gamma G protein,Chemokine:Class B GPCR{p}:Beta-gamma G protein,Chondroitin sulfate,Class A GPCR,Class A GPCR: Chemokine,Class A GPCR:Chemokine:Beta-gamma:GDP:alpha G protein,Class A GPCR:Chemokine:Beta-gamma:GTP:alpha G protein,Class B GPCR,Class B GPCR: Chemokine,Class B GPCR:Chemokine:Beta-gamma:GDP:alpha G protein,Class B GPCR:Chemokine:Beta-gamma:GTP:alpha G protein,Cr2,DNA,EMR1,EMR1:Proteoglycan,EMR2,EMR2:Chondroitin sulfate,ERK,ERK1,ERK1{active},ERK2,ERK2{active},ERK{active},ET-1:ETAR,ET-1:receptor,ET-2:receptor,FPLR1:annexin 1,FPR1,FPR1: N-formylated methionine,FPR1: Nonformylated bacterial protein,FPR1: fMLP,FPR1:aneexin1,FPRL1,FPRL1: Nonformylated bacterial protein,FPRL1:fMLP,FPRL2,FPRL2: endogenous ligand,GAP,GBeta2L1,GDP: alpha Gprotein,GIPC,GM-CSF,GRK,GRK2,GRK2:G beta Gamma,GRK3,GRK3:Beta Gamma G protein,GRK5,GRK5:p105,GRK5:p105{p},GRK6,GRK:AKT,GRK:ERK,GRK:GAP,GRK:PI3K,GTP:alpha G protein,Galpha i1,Galpha i2,Galpha i2{p},Galpha i3,Galphaq,Galphaq:GRK,Galphaq:GRK2,Galphaq:GRK3,IFN,IFN-alpha:receptor,IFN-alpha:receptor:STAT1,IFN-beta:receptor,IFN-beta:receptor:STAT1,IGF-IGFR1:STAT3,IGF:IGFR-1,IGFR1,IGFR1:RGS19:GIPC,IKK,IKK{active},IKappaB-alpha,IL-1,IL-10,IL-12,IL-1:IL-1R,IL-1alpha,IL-1beta,IL-6,IL-8,IL-8:IL-8R,IP-10,JNK,JNK3,JNK3:beta arrestin 2: ASK1,JNK3:beta arrestin2,JNK3{active},JNK{active},LPS,LPS:TLR4,LTB4:LTB4R1,LTB4:LTB4R2,Ligand:Angiotensin II type IA,Ligand:Beta 2 adrenergic receptor,Ligand:CXCR2,Ligand:CXCR4,Ligand:FPR,Ligand:PAR-2,MAPK,MAPK{active},MCP-1,MCP1,MMP2,MMP9,MMp2,MMp9,N-formylated methionine,NADPH oxidase,NADPH oxidase{active},NF-KappaB{active}:DNA,NF-kappaB,NF-kappaB{active},NK-1R,NK-1R:SP,NO,Nonformylated bacterial protein,PAF:PAFR,PAR1 precursor,PAR1{active},PAR2 precursor,PAR2 receptor activating peptides,PAR2{active},PARs,PARs{active},PGD2,PGE2,PI3K,PTx,Phosducin,Phosducin{p},RAF-1,RGS19,RGS19:GIPC,RGS19:GIPC:TrkA,RGS7,ROS,SP,SRC,STAT1,STAT3,Serum amyloid A,Serum amyloid A: FPR1,Serum amyloid A: FPRL1,Superoxide anion,Synucleins,Synucleins{p},TACR1,TNF,TRAF6,TRAF6{ub},TRAF6{ub} oligomer,Thrombaxane B2,Thromnaxane B2 protein,amyloid beta peptide,amyloid beta peptide: FPR1,amyloid beta peptide: FPRL1,annexin 1,beta arrestin1:IKappaB-alpha,beta arrestin1:TRAF6,beta arrestin2:IkkapB-alpha,beta arrestin2:TRAF6,beta arrestin:ERK{p},beta-arrestin,beta-arrestin1,beta-arrestin2,beta-arrestin:ERK,beta-arrestin:NF-kappaB,cox-2,endogenous ligand,fMLP,iNOS,p105,p38,p38{active},proteoglycan,second messenger dependent protein kinase,superoxide,thrombin,trkA,tubulin,tubulin{p}
As all immune responses have potential for damaging the host, tight regulationof such responses--in amplitude, space, time and character--is essential formaintaining health and homeostasis. It was thus inevitable that the initial waveof papers on the role of Toll-like receptors (TLRs), NOD-like receptors (NLRs)and RIG-I-like receptors (RLRs) in activating innate and adaptive immuneresponses would be followed by a second wave of reports focusing on themechanisms responsible for restraining and modulating signaling by thesereceptors. This overview outlines current knowledge and controversies about theimmunobiology of the RP105/MD-1 complex, a modulator of the most robustlysignaling TLR, TLR4.
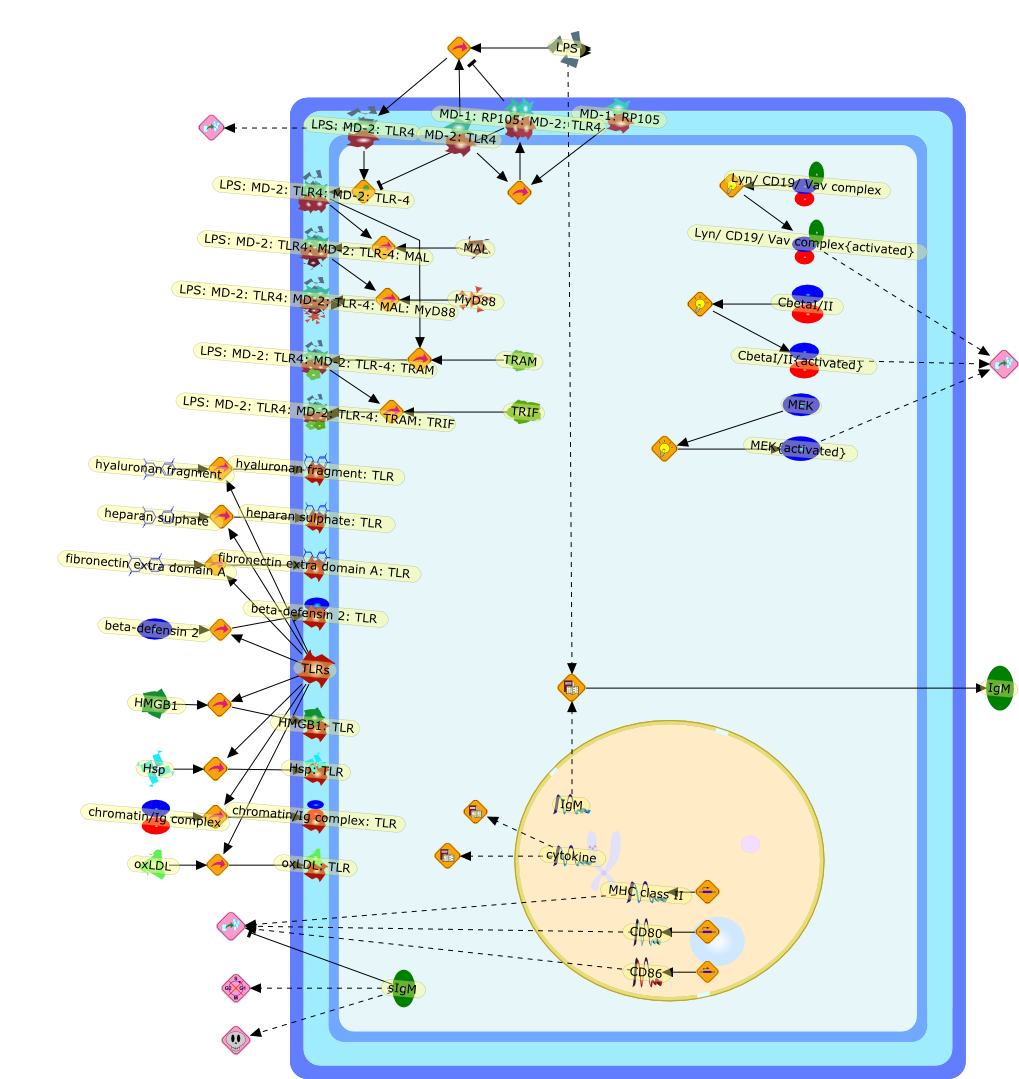{kind=link}
{kind=link}
CD80,CD86,CbetaI/II,CbetaI/II{activated},HMGB1,HMGB1: TLR,Hsp,Hsp: TLR,IgM,LPS,LPS: MD-2: TLR4,LPS: MD-2: TLR4: MD-2: TLR-4,LPS: MD-2: TLR4: MD-2: TLR-4: MAL,LPS: MD-2: TLR4: MD-2: TLR-4: MAL: MyD88,LPS: MD-2: TLR4: MD-2: TLR-4: TRAM,LPS: MD-2: TLR4: MD-2: TLR-4: TRAM: TRIF,Lyn/ CD19/ Vav complex,Lyn/ CD19/ Vav complex{activated},MAL,MD-1: RP105,MD-1: RP105: MD-2: TLR4,MD-2: TLR4,MEK,MEK{activated},MHC class II,MyD88,TLRs,TRAM,TRIF,beta-defensin 2,beta-defensin 2: TLR,chromatin/Ig complex,chromatin/Ig complex: TLR,cytokine,fibronectin extra domain A,fibronectin extra domain A: TLR,heparan sulphate,heparan sulphate: TLR,hyaluronan fragment,hyaluronan fragment: TLR,oxLDL,oxLDL: TLR,sIgM
Macrophage phenotype and activation are regulated by cytokines that use theJak-STAT signaling pathway, microbial recognition receptors that include TLRs,and immunoreceptors that signal via ITAM motifs. The amplitude and qualitativenature of macrophage activation are determined by crosstalk among thesesignaling pathways. Basal ITAM signaling restrains macrophage responses to TLRsand other activating ligands, whereas strong ITAM signals synergize with thesame ligands to activate cells strongly. Similarly, basal ITAM signalingaugments IFN signaling and function of receptor activator of NF-kappaB, butextensive ITAM activation inhibits Jak-STAT signaling. Thus, intensity andduration of ITAM signaling determine whether ITAM-coupled receptors augment orattenuate TLR and Jak-STAT responses. IFN-gamma synergizes with TLRs in part bysuppressing TLR-induced feedback inhibition, mediated by IL-10 and Stat3, by amechanism that depends on glycogen synthase kinase (GSK)3 regulation of AP-1 andCREB. IFN-gamma suppresses TLR2 and TLR4 induction/activation of AP-1 byoverlapping mechanisms that include regulation of MAPKs, GSK3-dependentsuppression of DNA binding, and decreased Fos and Jun protein expression andstability. IFN-gamma suppression of TLR-induced activation of AP-1 anddownstream target genes challenges current concepts about the inflammatory roleof AP-1 proteins in macrophage activation and is consistent with a role for AP-1in the generation of noninflammatory osteoclasts. Jak-STAT, TLR, and ITAMpathways are basally active in macrophages and strongly induced during innateresponses. Thus, signal transduction crosstalk is regulated in a dynamic manner,which differs under homeostatic and pathologic conditions, and dysregulation ofsignal transduction crosstalk may contribute to pathogenesis of chronicinflammatory diseases.
{kind=link}
{kind=link}
(stat1{p})2,AKT,AKT{active},AP-1,AP-1{p},CREB,CREB/activatingfactor,CREB/activatingfactor{act},CREB{pS},CXCL10,CXCL9,CaMKII,DAP12,DNA,DNA:(stat1{p})2,DNA:AP1,DNA:histone,DNA:histone{p},ERK,ERK{active},Ets,Ets{active},FcRgamma,FcgammaRI,GSK-3beta,IFN-alpha,IFN-gamma,IFN-gamma:IFN-gammaR:JAK,IFN-gamma:IFN-gammaR:JAK{act},IFN-gammaR:JAK,IL-10,IL-12,IL-6,IL10,IRFs,IRFs{active},ITAM,ITAM receptor,ITAM receptor lgand: ITAM receptor,ITAM receptor lgand: ITAM receptor:ITAM,ITAM receptor lgand: ITAM receptor:ITAM{p},ITAM receptor lgand: ITAM receptor:ITAM{p}:syk,ITAM receptor ligand,JNK,JNK{active},LPS,LPS:TLR4,MAPKs,MAPKs{active},MMP,MMP1,NF-kappaB,NF-kappaB{active},NFATc1,NFATc1{active},PAI-2,PKC,PKC{active},PLCgamma,PLCgamma{active},PTK,PTK{active},SIRPbeta1:DAP12,SIRPbeta1:DAP12:ligand,STAT1,STAT1{active},STAT1{p},STAT3,STAT3{active},Semaphorin 6D,Src family,Syk,TLR,TLR ligand,TLR ligand:TLR,TLR2,TLR2 ligand,TLR2 ligand:TLR2,TLR4,TNF-alpha,TREM2 ligand,TREM2 ligand:TREM2,TREM2:DAP12,c-Fos,c-fos,c-jun,ca2+,e65,gsk-3beta{p},inflammatory genes,p38,p38{active},plexin A1-TREM2-DAP12,plexin A1-TREM2-DAP12-Semaphorin 6D,type I IFN
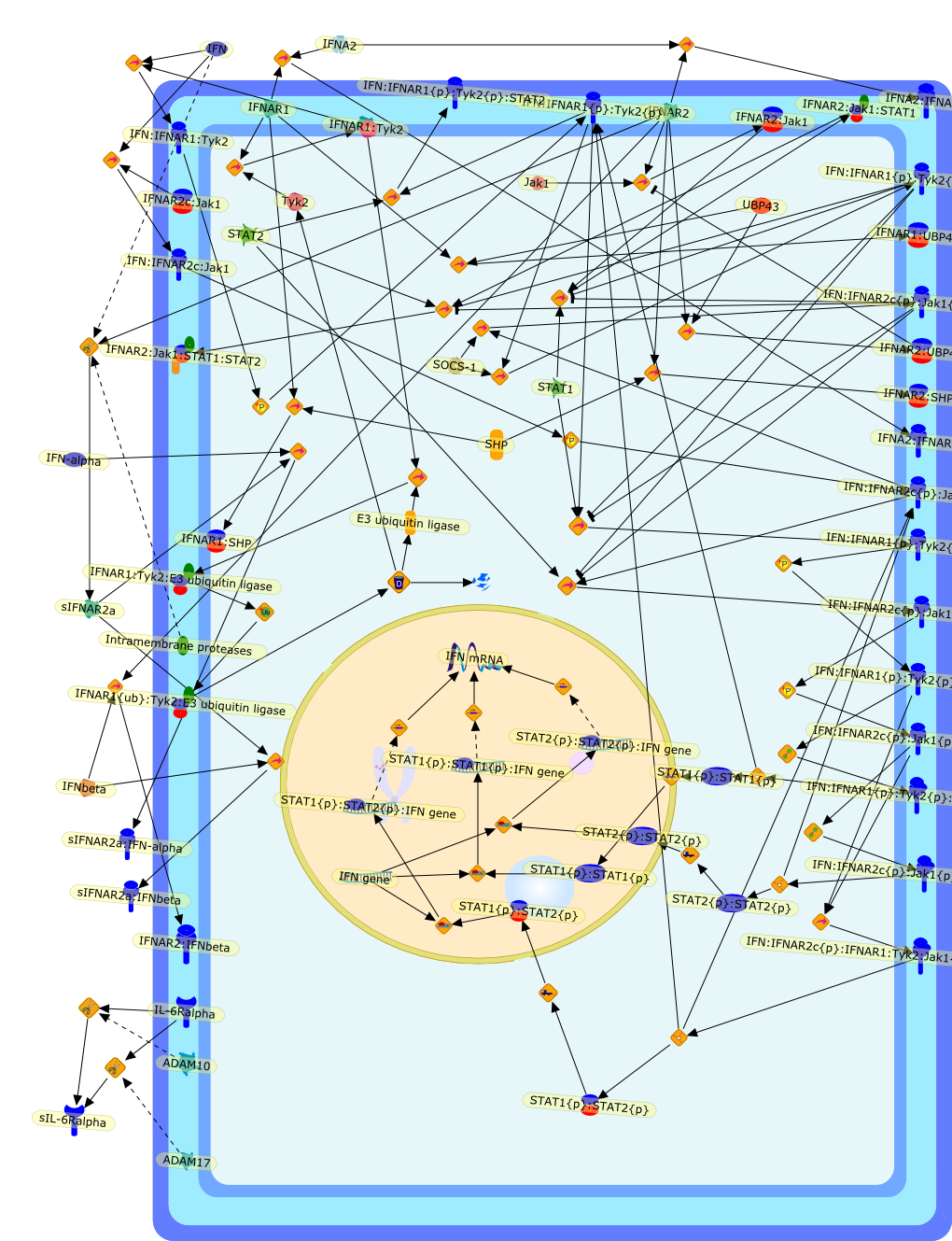{kind=link}
{kind=link}
ADAM10,ADAM17,E3 ubiquitin ligase,IFN,IFN gene,IFN mRNA,IFN-alpha,IFN:IFNAR1:Tyk2,IFN:IFNAR1{p}:Tyk2{p},IFN:IFNAR1{p}:Tyk2{p}:SOCS-1,IFN:IFNAR1{p}:Tyk2{p}:STAT1,IFN:IFNAR1{p}:Tyk2{p}:STAT1{p},IFN:IFNAR1{p}:Tyk2{p}:STAT1{p}(2),IFN:IFNAR1{p}:Tyk2{p}:STAT2,IFN:IFNAR2c:Jak1,IFN:IFNAR2c{p}:IFNAR1:Tyk2:Jak1{p}:STAT2{p}:STAT1{p},IFN:IFNAR2c{p}:Jak1{p},IFN:IFNAR2c{p}:Jak1{p}:SOCS-1,IFN:IFNAR2c{p}:Jak1{p}:STAT2,IFN:IFNAR2c{p}:Jak1{p}:STAT2{p},IFN:IFNAR2c{p}:Jak1{p}:STAT2{p}(2),IFNA2,IFNA2:IFNAR1,IFNA2:IFNAR2,IFNAR1,IFNAR1:SHP,IFNAR1:Tyk2,IFNAR1:Tyk2:E3 ubiquitin ligase,IFNAR1:UBP43,IFNAR1{ub}:Tyk2:E3 ubiquitin ligase,IFNAR2,IFNAR2:IFNbeta,IFNAR2:Jak1,IFNAR2:Jak1:STAT1,IFNAR2:Jak1:STAT1:STAT2,IFNAR2:SHP,IFNAR2:UBP43,IFNAR2c:Jak1,IFNbeta,IL-6Ralpha,Intramembrane proteases,Jak1,SHP,SOCS-1,STAT1,STAT1{p}:STAT1{p},STAT1{p}:STAT1{p}:IFN gene,STAT1{p}:STAT2{p},STAT1{p}:STAT2{p}:IFN gene,STAT2,STAT2{p}:STAT2{p},STAT2{p}:STAT2{p}:IFN gene,Tyk2,UBP43,degradants,sIFNAR2a,sIFNAR2a:IFN-alpha,sIFNAR2a:IFNbeta,sIL-6Ralpha
Transcription factors of the interferon regulatory factor (IRF) family commandsthe entire type I interferon (IFN) system from induction of IFNs to diverse IFNresponses, thereby providing a principal basis for host resistance againstpathogens. However, the family has various additional roles. Regulating thedevelopment of the immune system, IRFs shape the establishment and execution ofinnate and adaptive immunity. IRFs also regulate growth and differentiation ofmany cell types, thus playing a role in leukemia and other cancers. In addition,evidence indicates that IRFs confer antiviral mechanisms not directly ascribedto the IFN system. This review deals with the diverse roles of IRFs in hostdefense and discusses the molecular mechanisms by which they regulate targetgene transcription.
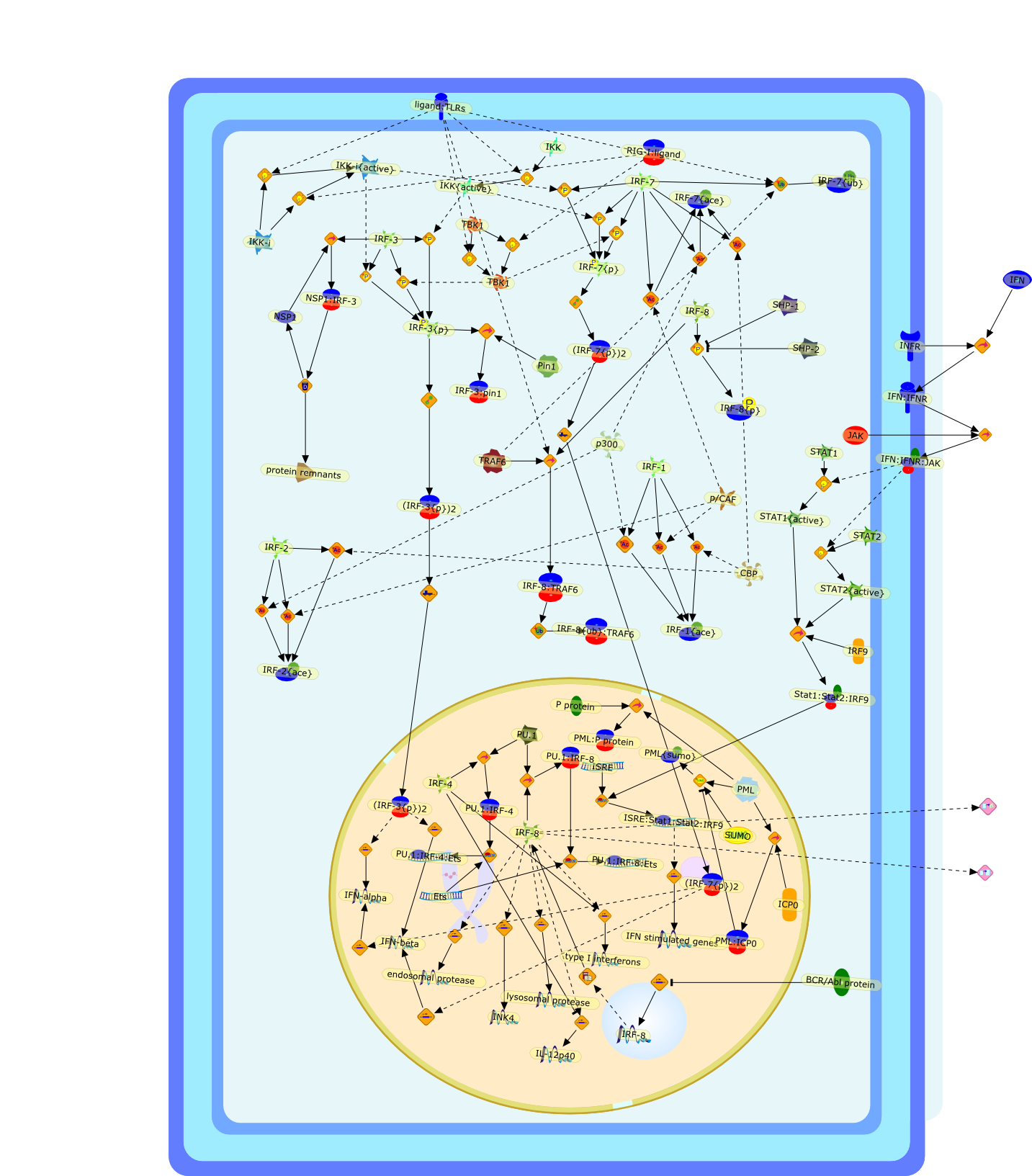{kind=link}
{kind=link}
(IRF-3{p})2,(IRF-7{p})2,BCR/Abl protein,CBP,Ets,ICP0,IFN,IFN stimulated genes,IFN-alpha,IFN-beta,IFN:IFNR,IFN:IFNR:JAK,IKK,IKK-i,IKK-i{active},IKK{active},IL-12p40,INFR,INK4,IRF-1,IRF-1{ace},IRF-2,IRF-2{ace},IRF-3,IRF-3:pin1,IRF-3{p},IRF-4,IRF-7,IRF-7{ace},IRF-7{p},IRF-7{ub},IRF-8,IRF-8:TRAF6,IRF-8{p},IRF-8{ub}:TRAF6,IRF9,ISRE,ISRE:Stat1:Stat2:IRF9,JAK,NSP1,NSP1:IRF-3,P protein,PML,PML:ICP0,PML:P protein,PML{sumo},PU.1,PU.1:IRF-4,PU.1:IRF-4:Ets,PU.1:IRF-8,PU.1:IRF-8:Ets,Pin1,RIG-I:ligand,SHP-1,SHP-2,STAT1,STAT1{active},STAT2,STAT2{active},SUMO,Stat1:Stat2:IRF9,TBK1,TRAF6,endosomal protease,ligand:TLRs,lysosomal protease,p/CAF,p300,protein remnants,type I interferons
Nitric oxide (NO) is synthesized from arginine and O2 by NO synthase (NOS).Citrulline formed as a by-product of the NOS reaction can be recycled toarginine by argininosuccinate synthetase (AS) and argininosuccinate lyase (AL).We found that AS and sometimes AL are coinduced with inducible NOS (iNOS) invarious cells. In these cells, NO was synthesized from citrulline (via arginine)as well as from arginine, indicating operation of the citrulline-NO cycle. Onthe other hand, we found that arginase isoforms (types I and II) are coinducedwith iNOS by LPS in rodent tissues and cultured macrophages. Km values forarginine of arginase I and II (approximately 10 mmol/L) are much higher thanthat of iNOS (approximately 5 micromol/L), whereas Vmax of arginase I and IIwere 10(3)-10(4) times higher than that of iNOS in activated macrophages. Thus,Vmax/Km values of arginases were close to that of iNOS, and these enzymes wereexpected to compete for arginine in the cells. In fact, NO production by iNOS inactivated macrophages was decreased by coinduction of arginase I or arginase II.Low concentrations of NO protect cells from apoptosis, whereas excessive NOcauses apoptosis. We found that NO causes endoplasmic reticulum (ER) stress,induces a transcription factor, CAAT/enhancer binding protein (C/EBP) homologousprotein (CHOP), and leads to apoptosis. These results suggest that the argininemetabolic enzymes and the ER stress-CHOP pathway can be good targets to regulateNO production and NO-mediated apoptosis in diseases associated withoverproduction or impaired production of NO.
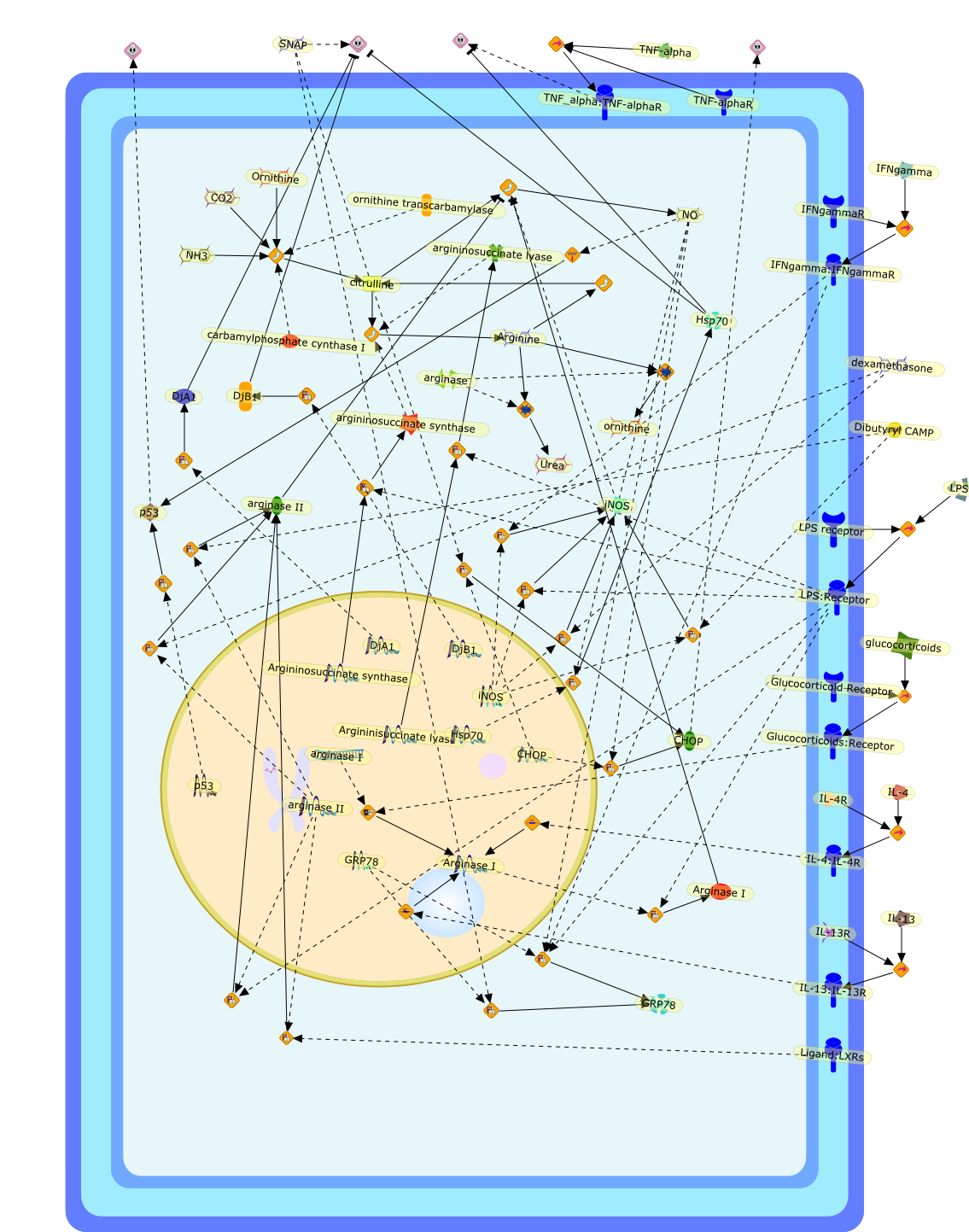{kind=link}
{kind=link}
Arginase I,Arginine,Argininisuccinate lyase,Argininosuccinate synthase,CHOP,CO2,Dibutyryl CAMP,DjA1,DjB1,GRP78,Glucocorticoid Receptor,Glucocorticoids:Receptor,Hsp70,IFNgamma,IFNgamma:IFNgammaR,IFNgammaR,IL-13,IL-13:IL-13R,IL-13R,IL-4,IL-4:IL-4R,IL-4R,LPS,LPS receptor,LPS:Receptor,Ligand:LXRs,NH3,NO,Ornithine,SNAP,TNF-alpha,TNF-alphaR,TNF_alpha:TNF-alphaR,Urea,arginase,arginase I,arginase II,argininosuccinate lyase,argininosuccinate synthase,carbamylphosphate cynthase I,citrulline,dexamethasone,glucocorticoids,iNOS,ornithine,ornithine transcarbamylase,p53
Pathogens are sensed by pattern recognition receptors (PRRs), which are germline-encoded receptors, including transmembrane Toll-like receptors (TLRs) andcytosolic nucleotide oligomerisation domain (NOD) proteins, containingleucine-rich repeats (NLRs). Activation of PRRs by specific pathogen-associatedmolecular patterns (PAMPs) results in genomic responses in host cells involvingactivation transcription factors and the induction of genes. There are now atleast 10 TLRs in humans and 13 in mice, and 2 NLRs (NOD1 and NOD2). TLRsignalling is via interactions with adaptor proteins including MyD88 andtoll-receptor associated activator of interferon (TRIF). NOD signalling is viathe inflammasome and involves activation of Rip-like interactive clarp kinase(RICK). Bacterial lipopolysaccharide (LPS) from Gram-negative bacteria is thebest-studied PAMP and is activated by or 'sensed' by TLR4. Lipoteichoic acid(LTA) from Gram-positive bacteria is sensed by TLR2. TLR4 and TLR2 havedifferent signalling cascades, although activation of either results in symptomsof sepsis and shock. This review describes the rapidly expanding field ofpathogen-sensing receptors and uses LPS and LTA as examples of how thesepathways parallel and diverge from each other. The role of pathogen-sensingpathways in disease is also discussed.
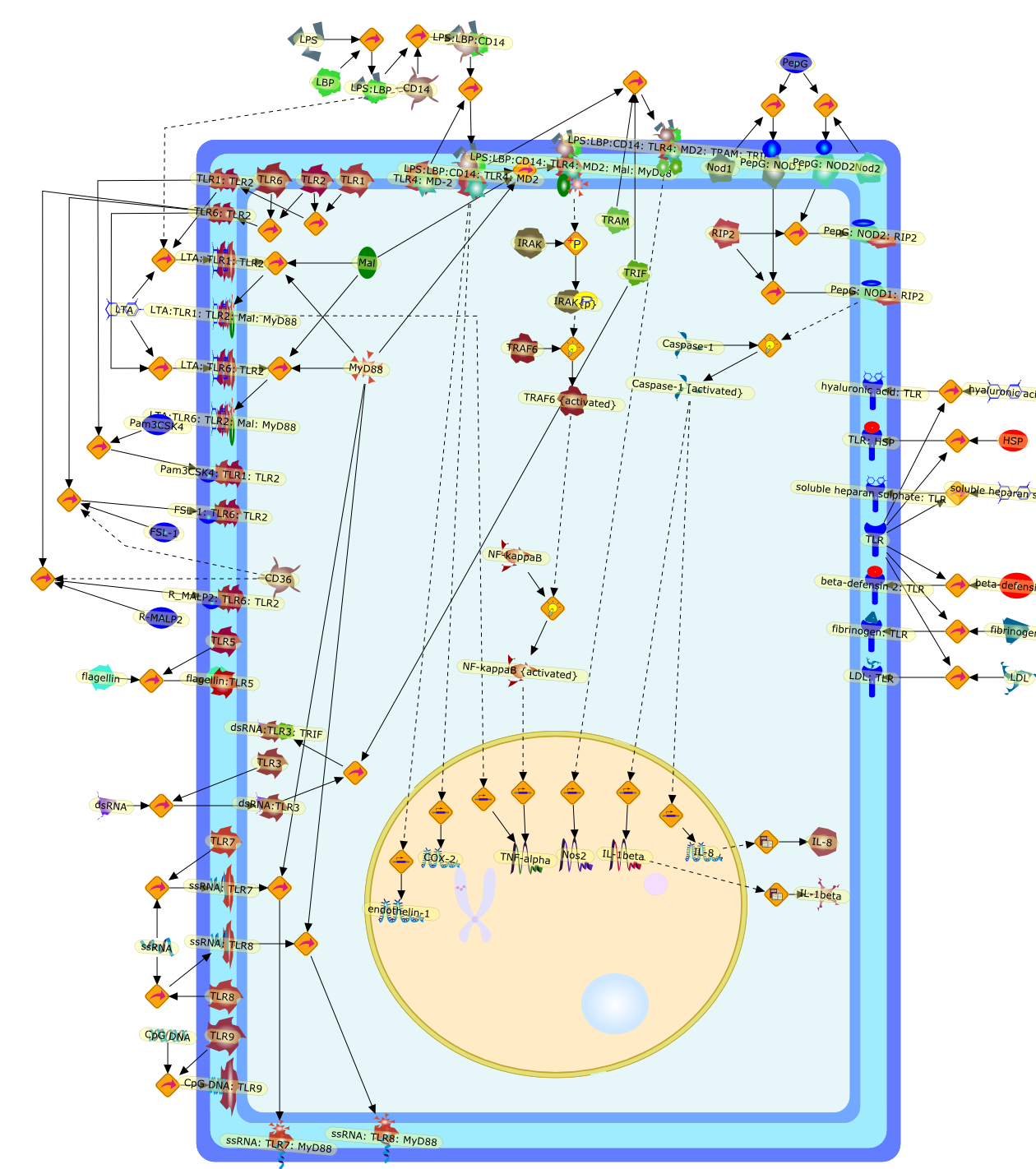{kind=link}
{kind=link}
CD14,CD36,COX-2,Caspase-1,Caspase-1 [activated},CpG DNA,CpG DNA: TLR9,FSL-1,FSL-1: TLR6: TLR2,HSP,IL-1beta,IL-8,IRAK,IRAK{p},LBP,LDL,LDL: TLR,LPS,LPS:LBP,LPS:LBP:CD14,LPS:LBP:CD14: TLR4: MD2,LPS:LBP:CD14: TLR4: MD2: Mal: MyD88,LPS:LBP:CD14: TLR4: MD2: TRAM: TRIF,LTA,LTA: TLR1: TLR2,LTA: TLR6: TLR2,LTA:TLR1: TLR2: Mal: MyD88,LTA:TLR6: TLR2: Mal: MyD88,Mal,MyD88,NF-kappaB,NF-kappaB {activated},Nod1,Nod2,Nos2,Pam3CSK4,Pam3CSK4: TLR1: TLR2,PepG,PepG: NOD1,PepG: NOD1: RIP2,PepG: NOD2,PepG: NOD2: RIP2,R-MALP2,RIP2,R_MALP2: TLR6: TLR2,TLR,TLR1,TLR1: TLR2,TLR2,TLR3,TLR4: MD-2,TLR5,TLR6,TLR6: TLR2,TLR7,TLR8,TLR9,TLR: HSP,TNF-alpha,TRAF6,TRAF6 {activated},TRAM,TRIF,beta-defensin 2,beta-defensin 2: TLR,dsRNA,dsRNA:TLR3,dsRNA:TLR3: TRIF,endothelin-1,fibrinogen,fibrinogen: TLR,flagellin,flagellin:TLR5,hyaluronic acid,hyaluronic acid: TLR,soluble heparan sulphate,soluble heparan sulphate: TLR,ssRNA,ssRNA: TLR7,ssRNA: TLR7: MyD88,ssRNA: TLR8,ssRNA: TLR8: MyD88
Type I interferons (IFN) and dendritic cells (DC) share an overlapping history,with rapidly accumulating evidence for vital roles for both production of type 1IFN by DC and the interaction of this IFN both with DC and components of theinnate and adaptive immune responses. Within the innate immune response, theplasmacytoid DC (pDC) are the "professional" IFN producing cells, expressingspecialized toll-like receptors (TLR7 and -9) and high constitutive expressionof IRF-7 that allow them to respond to viruses with rapid and extremely robustIFN production; following activation and production of IFN, the pDC subsequentlymature into antigen presenting cells that help to shape the adaptive immuneresponse. However, like most cells in the body, the myeloid or conventional DC(mDC or cDC) also produce type I IFNs, albeit typically at a lower level thanthat observed with pDC, and this IFN is also important in innate and adaptiveimmunity induced by these classic antigen presenting cells. These two major DCsubsets and their IFN products interact both with each other as well as with NKcells, monocytes, T helper cells, T cytotoxic cells, T regulatory cells and Bcells to orchestrate the early immune response. This review discusses some ofthe converging history of DC and IFN as well as mechanisms for IFN induction inDC and the effects of this IFN on the developing immune response.
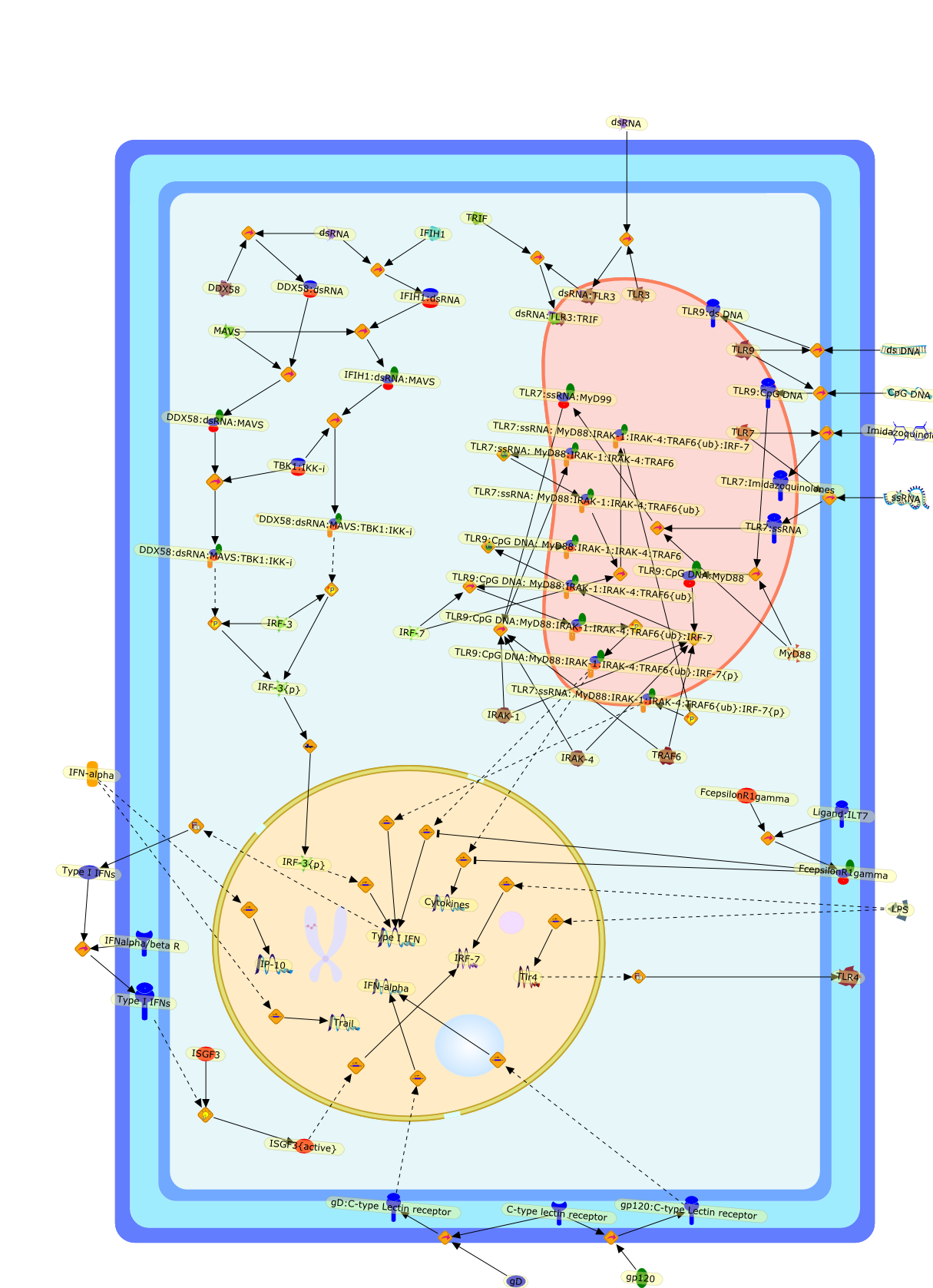{kind=link}
{kind=link}
C-type lectin receptor,CpG DNA,Cytokines,DDX58,DDX58:dsRNA,DDX58:dsRNA:MAVS,DDX58:dsRNA:MAVS:TBK1:IKK-i,FcepsilonR1gamma,IFIH1,IFIH1:dsRNA,IFIH1:dsRNA:MAVS,IFN-alpha,IFNalpha/beta R,IP-10,IRAK-1,IRAK-4,IRF-3,IRF-3{p},IRF-7,ISGF3,ISGF3{active},Imidazoquinolones,LPS,Ligand:ILT7,MAVS,MyD88,TBK1:IKK-i,TLR3,TLR4,TLR7,TLR7:Imidazoquinolones,TLR7:ssRNA,TLR7:ssRNA: MyD88:IRAK-1:IRAK-4:TRAF6,TLR7:ssRNA: MyD88:IRAK-1:IRAK-4:TRAF6{ub},TLR7:ssRNA: MyD88:IRAK-1:IRAK-4:TRAF6{ub}:IRF-7,TLR7:ssRNA: MyD88:IRAK-1:IRAK-4:TRAF6{ub}:IRF-7{p},TLR7:ssRNA:MyD99,TLR9,TLR9:CpG DNA,TLR9:CpG DNA: MyD88:IRAK-1:IRAK-4:TRAF6,TLR9:CpG DNA: MyD88:IRAK-1:IRAK-4:TRAF6{ub},TLR9:CpG DNA:MyD88,TLR9:CpG DNA:MyD88:IRAK-1:IRAK-4:TRAF6{ub}:IRF-7,TLR9:CpG DNA:MyD88:IRAK-1:IRAK-4:TRAF6{ub}:IRF-7{p},TLR9:ds DNA,TRAF6,TRIF,Tlr4,Trail,Type I IFN,Type I IFNs,ds DNA,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,gD,gD:C-type Lectin receptor,gp120,gp120:C-type Lectin receptor,ssRNA
Toll-like receptors (TLRs) are responsible for the recognition of a variety ofmicrobial pathogens and the initial induction of immune and inflammatoryresponses. These responses are normally restricted by the adrenally producedglucocorticoid hormones which provide a feedback mechanism to curb unabatedinflammation. Glucocorticoids act through a ligand-dependent transcriptionfactor-the glucocorticoid receptor (GR), which engages in a complex network ofprotein:protein and protein:DNA interactions ultimately activating or repressingtarget gene transcription. Not surprisingly, multiple mechanisms account for theglucocorticoid interference with TLR signaling including enhanced expression ofthe natural inhibitors of TLR pathways, direct repression of TLR-activatedtranscriptional regulators and cross-utilization of cofactors essential for bothGR and TLR signaling. Here we discuss recent and unexpected examples ofcrosstalk between the two transcriptional networks and the emerging role of GRin the regulation of innate immunity.
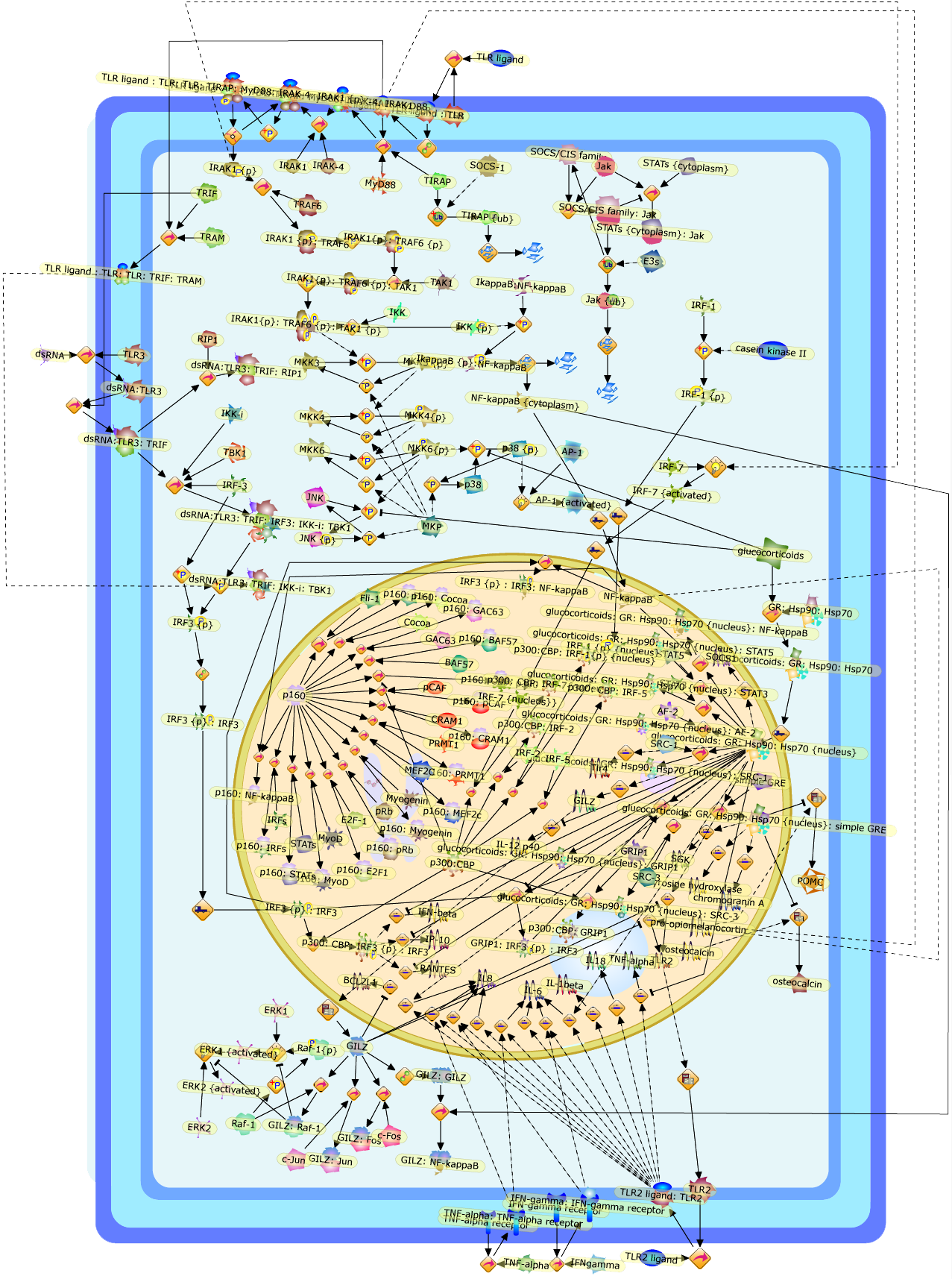{kind=link}
{kind=link}
AF-2,AP-1,AP-1 {activated},BAF57,BCL2L1,CRAM1,Cocoa,E2F-1,E3s,ERK1,ERK1 {activated},ERK2,ERK2 {activated},Fli-1,GAC63,GILZ,GILZ: Fos,GILZ: GILZ,GILZ: Jun,GILZ: NF-kappaB,GILZ: Raf-1,GR: Hsp90: Hsp70,GRIP1,GRIP1: IRF3 {p} : IRF3,IFN-beta,IFN-gamma receptor,IFN-gamma: IFN-gamma receptor,IFNgamma,IKK,IKK {p},IKK-i,IL-12 p40,IL-1beta,IL-6,IL18,IL8,IP-10,IRAK-4,IRAK1,IRAK1 {p},IRAK1 {p}: TRAF6,IRAK1{p}: TRAF6 {p},IRAK1{p}: TRAF6 {p}: TAK1,IRAK1{p}: TRAF6 {p}: TAK1 {p},IRF-1,IRF-1 {p},IRF-1 {p} {nucleus},IRF-2,IRF-3,IRF-5,IRF-7,IRF-7 {activated},IRF-7 {nucleus}},IRF3 {p},IRF3 {p} : IRF3,IRF3 {p} : IRF3: NF-kappaB,IRFs,IkappaB remnants,IkappaB {p}:NF-kappaB,IkappaB:NF-kappaB,JNK,JNK {p},Jak,Jak remnants,Jak {ub},MEF2C,MKK3,MKK3 {p},MKK4,MKK4{p},MKK6,MKK6{p},MKP,MyD88,MyoD,Myogenin,NF-kappaB,NF-kappaB {cytoplasm},POMC,PRMT1,RANTES,RIP1,Raf-1,Raf-1{p},SGK,SOCS-1,SOCS/CIS family,SOCS/CIS family: Jak,SOCS1,SRC-1,SRC-3,STAT3,STAT5,STATs,STATs {cytoplasm},STATs {cytoplasm}: Jak,TAK1,TBK1,TIRAP,TIRAP {ub},TIRAp ramnents,TLR ligand,TLR ligand : TLR,TLR ligand : TLR: TLR,TLR ligand : TLR: TLR: TIRAP: MyD88,TLR ligand : TLR: TLR: TIRAP: MyD88: IRAK-4: IRAK1,TLR ligand : TLR: TLR: TIRAP: MyD88: IRAK-4: IRAK1 {p},TLR ligand : TLR: TLR: TRIF: TRAM,TLR2,TLR2 ligand,TLR2 ligand: TLR2,TLR3,TLRs,TNF-alpha,TNF-alpha receptor,TNF-alpha: TNF-alpha receptor,TRAF6,TRAM,TRIF,Tlr4,c-Fos,c-Jun,casein kinase II,chromogranin A,dsRNA,dsRNA:TLR3,dsRNA:TLR3: TRIF,dsRNA:TLR3: TRIF: IKK-i: TBK1,dsRNA:TLR3: TRIF: IRF3: IKK-i: TBK1,dsRNA:TLR3: TRIF: RIP1,glucocorticoids,glucocorticoids: GR: Hsp90: Hsp70,glucocorticoids: GR: Hsp90: Hsp70 {nucleus},glucocorticoids: GR: Hsp90: Hsp70 {nucleus}: AF-2,glucocorticoids: GR: Hsp90: Hsp70 {nucleus}: GRIP1,glucocorticoids: GR: Hsp90: Hsp70 {nucleus}: NF-kappaB,glucocorticoids: GR: Hsp90: Hsp70 {nucleus}: SRC-1,glucocorticoids: GR: Hsp90: Hsp70 {nucleus}: SRC-3,glucocorticoids: GR: Hsp90: Hsp70 {nucleus}: STAT3,glucocorticoids: GR: Hsp90: Hsp70 {nucleus}: STAT5,glucocorticoids: GR: Hsp90: Hsp70 {nucleus}: simple GRE,osteocalcin,p160,p160: BAF57,p160: CBP: p300,p160: CRAM1,p160: Cocoa,p160: E2F1,p160: Fli-1,p160: GAC63,p160: IRFs,p160: MEF2c,p160: MyoD,p160: Myogenin,p160: NF-kappaB,p160: PRMT1,p160: STATs,p160: pCAF,p160: pRb,p300: CBP: IRF-5,p300: CBP: IRF-7{nucleus},p300: CBP: IRF3 {p} : IRF3,p300:CBP,p300:CBP: GRIP1,p300:CBP: IRF-1{p} {nucleus},p300:CBP: IRF-2,p38,p38 {p},pCAF,pRb,pro-opiomelanocortin,simple GRE,tyrosine hydroxylase
Toll-like receptors (TLRs) are essential mediators of both innate and adaptiveimmunity by recognizing and eliciting responses upon invasion of pathogens. Theresponse of TLRs must be stringently regulated as exaggerated expression ofsignalling components as well as pro-inflammatory cytokines can have devastatingeffects on the host, resulting in chronic inflammatory diseases, autoimmunedisorders and aid in the pathogenesis of TLR-associated human diseases.Therefore, it is essential that negative regulators act at multiple levelswithin TLR signalling cascades, as well as through eliciting negative-feedbackmechanisms in order to synchronize the positive activation and negativeregulation of signal transduction to avert potentially harmful immunologicalconsequences. This review explores the various mechanisms employed by negativeregulators to ensure the appropriate modulation of both immune and inflammatoryresponses.
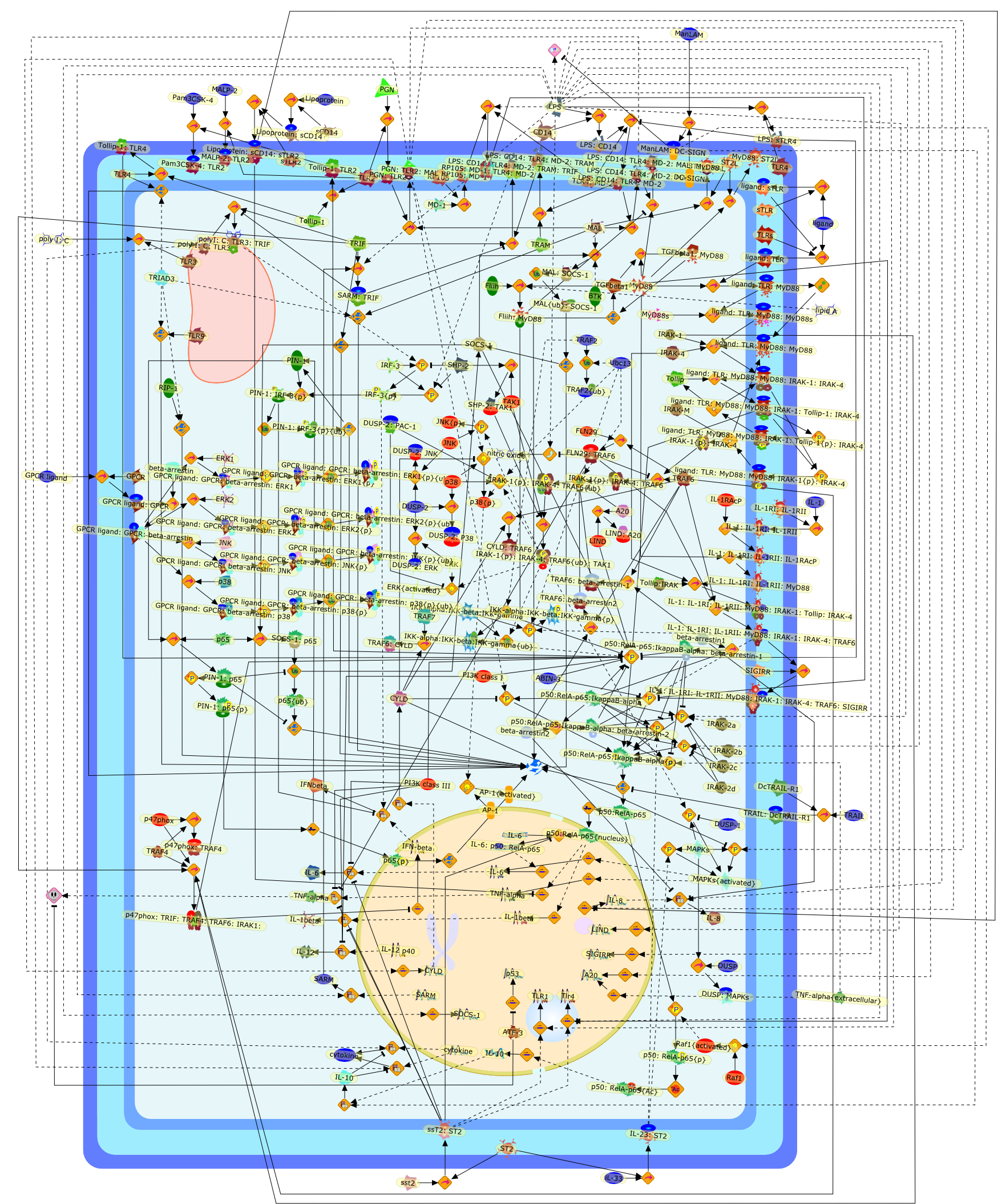{kind=link}
{kind=link}
A20,ABIN-3,AP-1,AP-1{activated},ATF-3,BTK,CD14,CYLD,CYLD: TRAF6,DC-SIGN,DUSP,DUSP-1,DUSP-2,DUSP-2: ERK,DUSP-2: JNK,DUSP-2: P38,DUSP-2: PAC-1,DUSP: MAPKs,DcTRAIL-R1,ERK,ERK1,ERK2,ERK{activated},FLN29,FLN29: TRAF6,Fliih,Fliih: MyD88,GPCR,GPCR ligand,GPCR ligand: GPCR,GPCR ligand: GPCR: beta-arrestin,GPCR ligand: GPCR: beta-arrestin: ERK1,GPCR ligand: GPCR: beta-arrestin: ERK1{p},GPCR ligand: GPCR: beta-arrestin: ERK1{p}{ub},GPCR ligand: GPCR: beta-arrestin: ERK2,GPCR ligand: GPCR: beta-arrestin: ERK2{p},GPCR ligand: GPCR: beta-arrestin: ERK2{p}{ub},GPCR ligand: GPCR: beta-arrestin: JNK,GPCR ligand: GPCR: beta-arrestin: JNK{p},GPCR ligand: GPCR: beta-arrestin: JNK{p}{ub},GPCR ligand: GPCR: beta-arrestin: p38,GPCR ligand: GPCR: beta-arrestin: p38{p},GPCR ligand: GPCR: beta-arrestin: p38{p}{ub},IFN-beta,IFNbeta,IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma{p},IKK-alpha:IKK-beta:IKK-gamma{ub},IL-1,IL-10,IL-12,IL-12 p40,IL-1: IL-1RI: IL-1RII,IL-1: IL-1RI: IL-1RII: IL-1RAcP,IL-1: IL-1RI: IL-1RII: MyD88,IL-1: IL-1RI: IL-1RII: MyD88: IRAK-1: IRAK-4: TRAF6,IL-1: IL-1RI: IL-1RII: MyD88: IRAK-1: IRAK-4: TRAF6: SIGIRR,IL-1: IL-1RI: IL-1RII: MyD88: IRAK-1: Tollip: IRAK-4,IL-1RAcP,IL-1RI: IL-1RII,IL-1beta,IL-23: ST2,IL-6,IL-6: p50: RelA-p65,IL-8,IRAK-1,IRAK-1{p}: IRAK-4,IRAK-1{p}: IRAK-4: TRAF6,IRAK-1{p}: IRAK-4: TRAF6{ub},IRAK-1{p}: IRAK-4: TRAF6{ub}: TAK1,IRAK-2a,IRAK-2b,IRAK-2c,IRAK-2d,IRAK-4,IRAK-M,IRF-3,IRF-3{p},JNK,JNK{p},LIND,LIND: A20,LPS,LPS: CD14,LPS: CD14: TLR4: MD-2,LPS: CD14: TLR4: MD-2: MAL,LPS: CD14: TLR4: MD-2: MAL: MyD88,LPS: CD14: TLR4: MD-2: TRAM,LPS: CD14: TLR4: MD-2: TRAM: TRIF,LPS: sTLR4,Lipoprotein,Lipoprotein: sCD14,Lipoprotein: sCD14: sTLR2,MAL,MAL: SOCS-1,MALP-2,MALP-2: TLR2,MAL{ub}: SOCS-1,MAPKs,MAPKs{activated},MD-1,ManLAM,ManLAM: DC-SIGN,MyD88,MyD88: ST2L,MyD88s,PGN,PGN: TLR2,PGN: TLR2: MAL,PI3K class I,PI3K class III,PIN-1,PIN-1: IRF-3{p},PIN-1: IRF-3{p}{ub},PIN-1: p65,PIN-1: p65{p},Pam3CSK-4,Pam3CSK-4: TLR2,RIP-1,RP105,RP105: MD-1,RP105: MD-1: TLR4: MD-2,Raf1,Raf1{activated},SARM,SARM: TRIF,SHP-2,SHP-2: TAK1,SIGIRR,SOCS-1,SOCS-1: p65,ST2,ST2L,ST2L: MAL,TAK1,TGFbeta1,TGFbeta1: MyD88,TLR1,TLR2,TLR3,TLR4,TLR4: MD-2,TLR9,TLRs,TNF-alpha,TNF-alpha{extracellular},TRAF2,TRAF2{ub},TRAF4,TRAF6,TRAF6: CYLD,TRAF6: beta-arrestin-1,TRAF6: beta-arrestin2,TRAF7,TRAIL,TRAIL: DcTRAIL-R1,TRAM,TRIAD3,TRIF,Tlr4,Tollip,Tollip-1,Tollip-1: TLR2,Tollip-1: TLR4,Tollip:IRAK,Ubc13,beta-arrestin,beta-arrestin1,beta-arrestin2,cytokine,iL-33,ligand,ligand: TLR,ligand: TLR: MyD88,ligand: TLR: MyD88: MyD88,ligand: TLR: MyD88: MyD88: IRAK-1: IRAK-4,ligand: TLR: MyD88: MyD88: IRAK-1: Tollip-1: IRAK-4,ligand: TLR: MyD88: MyD88: IRAK-1: Tollip-1{p}: IRAK-4,ligand: TLR: MyD88: MyD88: IRAK-1{p}: IRAK-4,ligand: TLR: MyD88: MyD88s,ligand: sTLR,lipid A,nitric oxide,p38,p38{p},p47phox,p47phox: TRAF4,p47phox: TRIF: TRAF4: TRAF6: IRAK1: ,p50: RelA-p65{Ac},p50: RelA-p65{p},p50:RelA-p65,p50:RelA-p65:IkappaB-alpha,p50:RelA-p65:IkappaB-alpha: beta-arrestin-1,p50:RelA-p65:IkappaB-alpha: beta-arrestin-2,p50:RelA-p65:IkappaB-alpha{p},p50:RelA-p65{nucleus},p53,p65,p65{p},p65{ub},poly I: C,poly I: C: TLR3,polyI: C: TLR3: TRIF,protein remnants,sCD14,sTLR,sTLR2,ssT2: ST2,sst2
Members of the Toll-like receptor (TLR) and interleukin-1 receptor (IL-1R)family play important roles in immunity and inflammation. They initiate commonintracellular signalling cascades leading to the activation of nuclear factor-?B(NF-?B) and other transcription factors that stimulate the expression of avariety of genes that shape an appropriate immune response. TLR/IL-1R signallinginvolves multiple proteinprotein interactions, but the mechanisms that regulatethese interactions are still largely unclear. In this context, Pellino proteinshave been suggested to function as evolutionary conserved scaffold proteins inTLR/IL-1R signalling. However, recently Pellino proteins were also proposed tofunction as novel ubiquitin ligases for IL-1R associated kinase 1 (IRAK-1). Herewe review our current knowledge on the expression, biological role and mechanismof action of Pellino proteins in TLR/IL-1R-induced signalling.
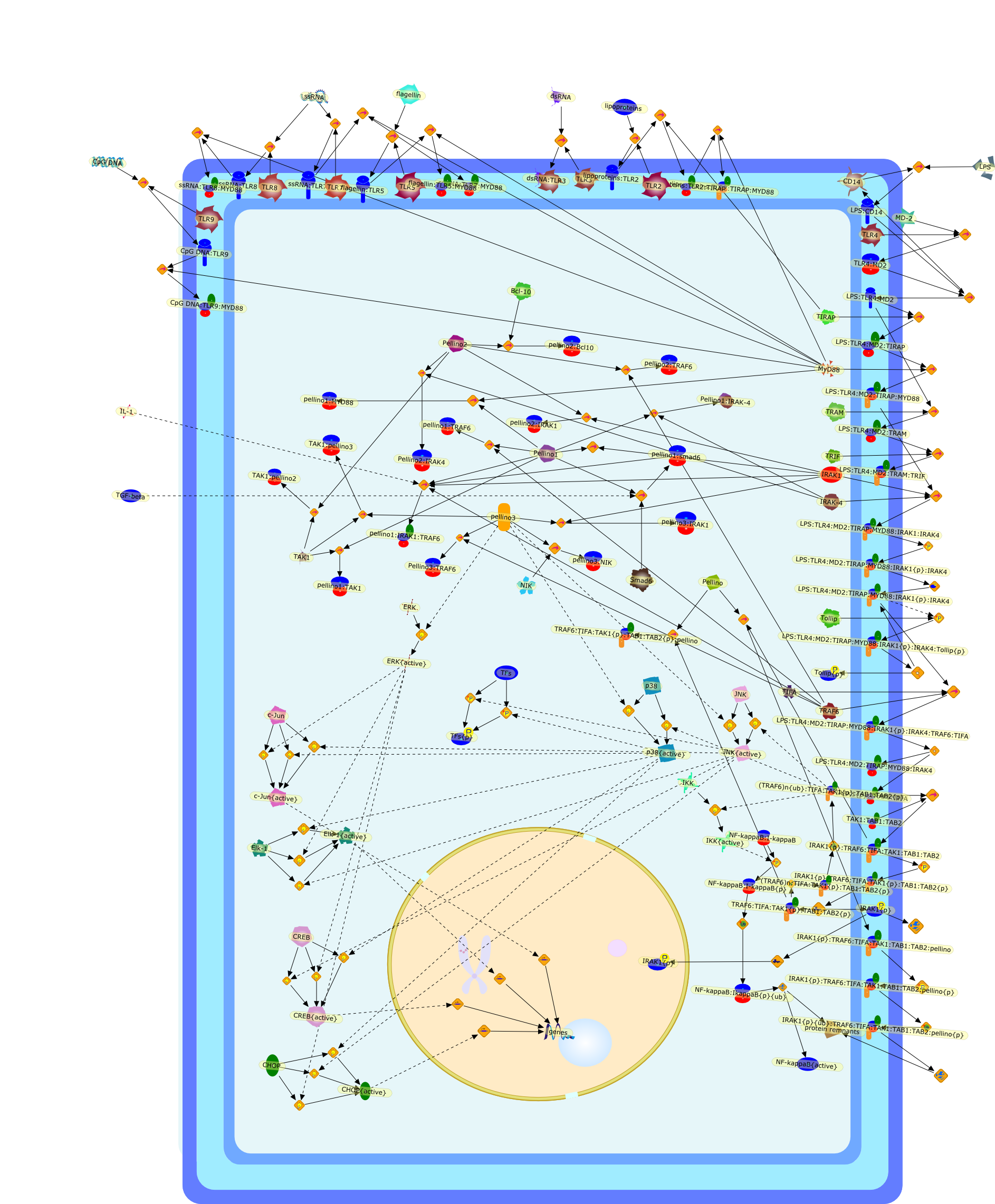{kind=link}
{kind=link}
(TRAF6)n:TIFA:TAK1{p}:TAB1:TAB2{p},(TRAF6)n{ub}:TIFA:TAK1{p}:TAB1:TAB2{p},Bcl-10,CD14,CHOP,CHOP{active},CREB,CREB{active},CpG DNA,CpG DNA:TLR9,CpG DNA:TLR9:MYD88,ERK,ERK{active},Elk-1,Elk-1{active},IKK,IKK{active},IL-1,IRAK-4,IRAK1,IRAK1{p},IRAK1{p}:TRAF6:TIFA,IRAK1{p}:TRAF6:TIFA:TAK1:TAB1:TAB2,IRAK1{p}:TRAF6:TIFA:TAK1:TAB1:TAB2:pellino,IRAK1{p}:TRAF6:TIFA:TAK1:TAB1:TAB2:pellino{p},IRAK1{p}:TRAF6:TIFA:TAK1{p}:TAB1:TAB2{p},IRAK1{p}{ub}:TRAF6:TIFA:TAK1:TAB1:TAB2:pellino{p},JNK,JNK{active},LPS,LPS:CD14,LPS:TLR4:MD2,LPS:TLR4:MD2:TIRAP,LPS:TLR4:MD2:TIRAP:MYD88,LPS:TLR4:MD2:TIRAP:MYD88:IRAK1:IRAK4,LPS:TLR4:MD2:TIRAP:MYD88:IRAK1{p}:IRAK4,LPS:TLR4:MD2:TIRAP:MYD88:IRAK1{p}:IRAK4:TRAF6:TIFA,LPS:TLR4:MD2:TIRAP:MYD88:IRAK1{p}:IRAK4:Tollip{p},LPS:TLR4:MD2:TIRAP:MYD88:IRAK4,LPS:TLR4:MD2:TRAM,LPS:TLR4:MD2:TRAM:TRIF,MD-2,MyD88,NF-kappaB:I-kappaB,NF-kappaB:I-kappaB{p},NF-kappaB:IkappaB{p}{ub},NF-kappaB{active},NIK,Pellino,Pellino1,Pellino1:IRAK-4,Pellino2,Pellino2:IRAK4,Pellino3:TRAF6,Smad6,TAK1,TAK1:TAB1:TAB2,TAK1:pellino2,TAK1:pellino3,TFs,TFs{p},TGF-beta,TIFA,TIRAP,TLR2,TLR3,TLR4,TLR4:MD2,TLR5,TLR7,TLR8,TLR9,TRAF6,TRAF6:TIFA:TAK1{p}:TAB1:TAB2{p},TRAF6:TIFA:TAK1{p}:TAB1:TAB2{p}:pellino,TRAM,TRIF,Tollip,Tollip{p},c-Jun,c-Jun{active},dsRNA,dsRNA:TLR3,flagellin,flagellin:TLR5,flagellin:TLR5:MYD88,genes,lipoproteins,lipoproteins:TLR2,lipoproteins:TLR2:TIRAP,lipoproteins:TLR2:TIRAP:MYD88,p38,p38{active},pellino1:IRAK1:TRAF6,pellino1:MYD88,pellino1:TAK1,pellino1:TRAF6,pellino1:smad6,pellino2:Bcl10,pellino2:IRAK1,pellino2:TRAF6,pellino3,pellino3:IRAK1,pellino3:NIK,protein remnants,ssDNA:TLR7:MYD88,ssRNA,ssRNA:TLR7,ssRNA:TLR8,ssRNA:TLR8:MYD88
The antiviral innate immune response follows the detection of viral componentsby host pattern recognition receptors (PRRs). Two families of PRRs have emergedas key sensors of viral infection: Toll-like receptors (TLRs) and retinoic acidinducible gene-I like RNA helicases (RLHs). TLRs patrol the extracellular andendosomal compartments; signalling results in a type-1 interferon responseand/or the production of pro-inflammatory cytokines. In contrast, RLHs surveythe cytoplasm for the presence of viral double-stranded RNA. In the face of suchhost defence, viruses have developed strategies to evade TLR/RLH signalling.Such host-virus interactions provide the opportunity for manipulation of PRRsignalling as a novel therapeutic approach.
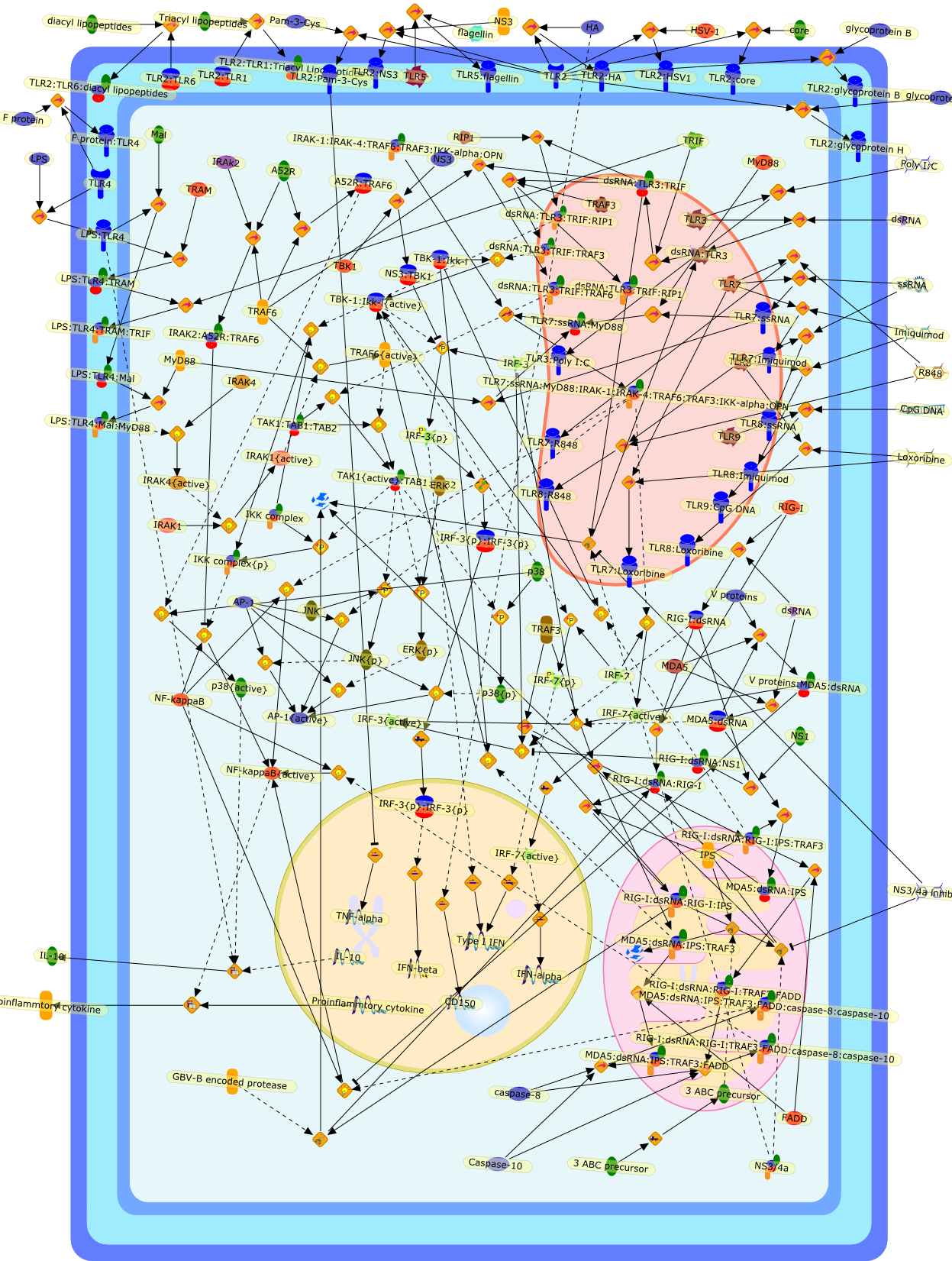{kind=link}
{kind=link}
3 ABC precursor,A52R,A52R:TRAF6,AP-1,AP-1{active},CD150,Caspase-10,CpG DNA,ERK,ERK{p},F protein,F protein:TLR4,FADD,GBV-B encoded protease,HA,HSV-1,IFN-alpha,IFN-beta,IKK complex,IKK complex{p},IL-10,IPS,IRAK-1:IRAK-4:TRAF6:TRAF3:IKK-alpha:OPN,IRAK1,IRAK1{active},IRAK2:A52R:TRAF6,IRAK4,IRAK4{active},IRAk2,IRF-3,IRF-3{active},IRF-3{p},IRF-3{p}:IRF-3{p},IRF-7,IRF-7{active},IRF-7{p},Imiquimod,JNK,JNK{p},LPS,LPS:TLR4,LPS:TLR4:Mal,LPS:TLR4:Mal:MyD88,LPS:TLR4:TRAM,LPS:TLR4:TRAM:TRIF,Loxoribine,MDA5,MDA5:dsRNA,MDA5:dsRNA:IPS,MDA5:dsRNA:IPS:TRAF3,MDA5:dsRNA:IPS:TRAF3:FADD,MDA5:dsRNA:IPS:TRAF3:FADD:caspase-8:caspase-10,Mal,MyD88,NF-kappaB,NF-kappaB{active},NS1,NS3,NS3/4a,NS3/4a inhibitor,NS3:TBK1,Pam-3-Cys,Poly I:C,Proinflammtory cytokine,R848,RIG-I,RIG-I:dsRNA,RIG-I:dsRNA:NS1,RIG-I:dsRNA:RIG-I,RIG-I:dsRNA:RIG-I:IPS,RIG-I:dsRNA:RIG-I:IPS:TRAF3,RIG-I:dsRNA:RIG-I:TRAF3:FADD,RIG-I:dsRNA:RIG-I:TRAF3:FADD:caspase-8:caspase-10,RIP1,TAK1:TAB1:TAB2,TAK1{active}:TAB1:TAB2,TBK-1:Ikk-i,TBK-1:Ikk-i{active},TBK1,TLR2,TLR2:HA,TLR2:HSV1,TLR2:NS3,TLR2:Pam-3-Cys,TLR2:TLR1,TLR2:TLR1:Triacyl Lipopeptides,TLR2:TLR6,TLR2:TLR6:diacyl lipopeptides,TLR2:core,TLR2:glycoprotein B,TLR2:glycoprotein H,TLR3,TLR3:Poly I:C,TLR4,TLR5,TLR5:flagellin,TLR7,TLR7:Imiquimod,TLR7:Loxoribine,TLR7:R848,TLR7:ssRNA,TLR7:ssRNA:MyD88,TLR7:ssRNA:MyD88:IRAK-1:IRAK-4:TRAF6:TRAF3:IKK-alpha:OPN,TLR8,TLR8:Imiquimod,TLR8:Loxoribine,TLR8:R848,TLR8:ssRNA,TLR9,TLR9:CpG DNA,TNF-alpha,TRAF3,TRAF6,TRAF6{active},TRAM,TRIF,Triacyl lipopeptides,Type I IFN ,V proteins,V proteins:MDA5:dsRNA,caspase-8,core,diacyl lipopeptides,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:RIP1,dsRNA:TLR3:TRIF:TRAF3,dsRNA:TLR3:TRIF:TRAF6,flagellin,glycoprotein B,glycoprotein H,p38,p38{active},p38{p},protein remnants,ssRNA
The Toll/IL-1 receptor (TIR) domain plays a central role in Toll-like receptor(TLR) signalling. All TLRs contain a cytoplasmic TIR domain, which, uponactivation, acts as a scaffold to recruit adaptor proteins. The adaptor proteinsMyD88, Mal, TRIF, TRAM and SARM are also characterized by the presence of a TIRdomain. MyD88, Mal, TRIF and TRAM associate with the TLRs via homophilic TIRdomain interactions whereas SARM utilizes its TIR domain to negatively regulateTRIF. It is well established that the differential recruitment of adaptors toTLRs provides a significant amount of specificity to the TLR-signallingpathways. Despite this, the TIR-TIR interface has not been well defined.However, structural studies have indicated the importance of TIR domain surfacesin mediating specific TIR-TIR interactions. Furthermore, recent findingsregarding the regulation of adaptors provide further insight into the crucialrole of the TIR domain in TLR signalling.
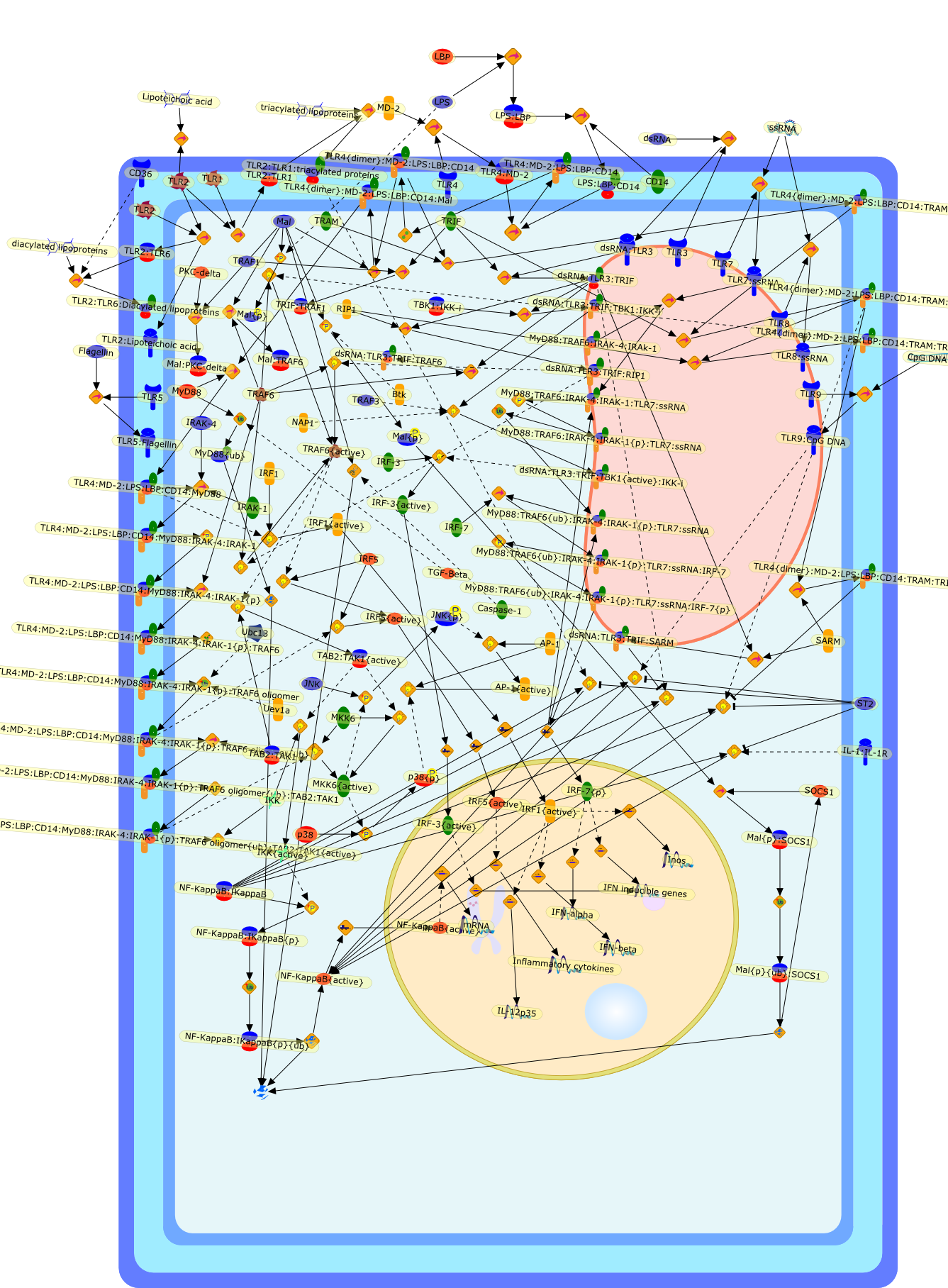{kind=link}
{kind=link}
AP-1,AP-1{active},Btk,CD14,CD36,Caspase-1,CpG DNA,Flagellin,IFN inducible genes,IFN-alpha,IFN-beta,IKK,IKK{active},IL-12p35,IL-1:IL-1R,IRAK-1,IRAK-4,IRF-3,IRF-3{active},IRF-7,IRF-7{p},IRF1,IRF1{active},IRF5,IRF5{active},Inflammatory cytokines,Inos,JNK,JNK{p},LBP,LPS,LPS:LBP,LPS:LBP:CD14,Lipoteichoic acid,MD-2,MKK6,MKK6{active},Mal,Mal:PKC-delta,Mal:TRAF6,Mal{p},Mal{p}:SOCS1,Mal{p}{ub}:SOCS1,MyD88,MyD88:TRAF6:IRAK-4:IRAK-1:TLR7:ssRNA,MyD88:TRAF6:IRAK-4:IRAK-1{p}:TLR7:ssRNA,MyD88:TRAF6:IRAK-4:IRAk-1,MyD88:TRAF6{ub}:IRAK-4:IRAK-1{p}:TLR7:ssRNA,MyD88:TRAF6{ub}:IRAK-4:IRAK-1{p}:TLR7:ssRNA:IRF-7,MyD88:TRAF6{ub}:IRAK-4:IRAK-1{p}:TLR7:ssRNA:IRF-7{p},MyD88{ub},NAP1,NF-KappaB:IKappaB,NF-KappaB:IKappaB{p},NF-KappaB:IKappaB{p}{ub},NF-KappaB{active},PKC-delta,Proteasome degradants,RIP1,SARM,SOCS1,ST2,TAB2:TAK1,TAB2:TAK1{active},TBK1:IKK-i,TGF-Beta,TLR1,TLR2,TLR2:Lipoteichoic acid,TLR2:TLR1,TLR2:TLR1:triacylated proteins,TLR2:TLR6,TLR2:TLR6:Diacylated lipoproteins,TLR3,TLR4,TLR4:MD-2,TLR4:MD-2:LPS:LBP:CD14,TLR4:MD-2:LPS:LBP:CD14:MyD88,TLR4:MD-2:LPS:LBP:CD14:MyD88:IRAK-4:IRAK-1,TLR4:MD-2:LPS:LBP:CD14:MyD88:IRAK-4:IRAK-1{p},TLR4:MD-2:LPS:LBP:CD14:MyD88:IRAK-4:IRAK-1{p}:TRAF6,TLR4:MD-2:LPS:LBP:CD14:MyD88:IRAK-4:IRAK-1{p}:TRAF6 oligomer,TLR4:MD-2:LPS:LBP:CD14:MyD88:IRAK-4:IRAK-1{p}:TRAF6 oligomer{ub},TLR4:MD-2:LPS:LBP:CD14:MyD88:IRAK-4:IRAK-1{p}:TRAF6 oligomer{ub}:TAB2:TAK1,TLR4:MD-2:LPS:LBP:CD14:MyD88:IRAK-4:IRAK-1{p}:TRAF6 oligomer{ub}:TAB2:TAK1{active},TLR4{dimer}:MD-2:LPS:LBP:CD14,TLR4{dimer}:MD-2:LPS:LBP:CD14:Mal,TLR4{dimer}:MD-2:LPS:LBP:CD14:TRAM,TLR4{dimer}:MD-2:LPS:LBP:CD14:TRAM:TRIF,TLR4{dimer}:MD-2:LPS:LBP:CD14:TRAM:TRIF:RIP1,TLR4{dimer}:MD-2:LPS:LBP:CD14:TRAM:TRIF:SARM,TLR5,TLR5:Flagellin,TLR7,TLR7:ssRNA,TLR8,TLR8:ssRNA,TLR9,TLR9:CpG DNA,TRAF1,TRAF3,TRAF6,TRAF6{active},TRAM,TRIF,TRIF:TRAF1,Ubc13,Uev1a,diacylated lipoproteins,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:RIP1,dsRNA:TLR3:TRIF:SARM,dsRNA:TLR3:TRIF:TBK1:IKK-i,dsRNA:TLR3:TRIF:TBK1{active}:IKK-i,dsRNA:TLR3:TRIF:TRAF6,mRNA,p38,p38{p},ssRNA,triacylated lipoproteins
Kinins have been reported to be produced and act at the site of injury andinflammation. Despite many reports that they are likely to initiate a particularcascade of inflammatory events, bradykinin (BK) has anti-inflammatory effects inthe brain mediated by glial cells. In the present review, we have attempted todescribe the complex responses and immediate reaction of glial cells to BK.Glial cells express BK receptors and induce Ca(2+)-dependent signal cascades.Among them, production of prostaglandin E(2) (PGE(2)), via B(1) receptors inprimary cultured microglia, has a negative feedback effect on lipopolysaccharide(LPS)-induced release of tumor necrosis factor-alpha (TNF-alpha) via increasingintracellular cyclic adenosine monophosphate (cAMP). In addition, BKup-regulates the production of neurotrophic factors such as nerve growth factor(NGF) via B(2) receptors in astrocytes. These results suggest that BK may haveanti-inflammatory and neuroprotective effects in the brain through multiplefunctions on glial cells. These observations may help to understand the paradoxon the role of kinins in the central nervous system and may be useful fortherapeutic strategy.
{kind=link}
{kind=link}
ATP,ATP:P2X,ATP:P2Y,BAPTA-AM,BDKRB1,BDNF,Bradykinin,Bradykinin receptor,Bradykinin:Bradykinin receptor,COX2,ET:ETR,ETR,IL-1beta,IL-6,LPS,LPS:TLR4,MMP-9,NGF,NS398,P2X,P2Y,PGE2,PLA2,RpcAMP,TLR4,TNF-alpha,calcium,endotheline-1,iNOS
Adipose tissue has long been regarded as a mostly resting tissue that isdedicated solely to energy storage and release. However, in recent years, thisview has changed dramatically following new insights into the metabolic andimmunological functions of preadipocytes and adipocytes. There are several linesof evidence for the involvement of adipose tissue in innate and acquired immuneresponses. First, adipocytes are potent producers of proinflammatory cytokines,such as interleukin-6 and tumor necrosis factor (TNF), and chemokines.Furthermore, adipocytes secrete high amounts of adipokines, such as leptin,adiponectin and resistin, that regulate monocyte/macrophage function, and alsosecrete molecules associated with the innate immune system, such as theC1qTNF-related protein superfamily. Finally, preadipocytes and adipocytesexpress a broad spectrum of functional Toll-like receptors and the former canconvert into macrophage-like cells. Collectively, these data clearly establishthe role of adipose tissue as a new member of the immune system.
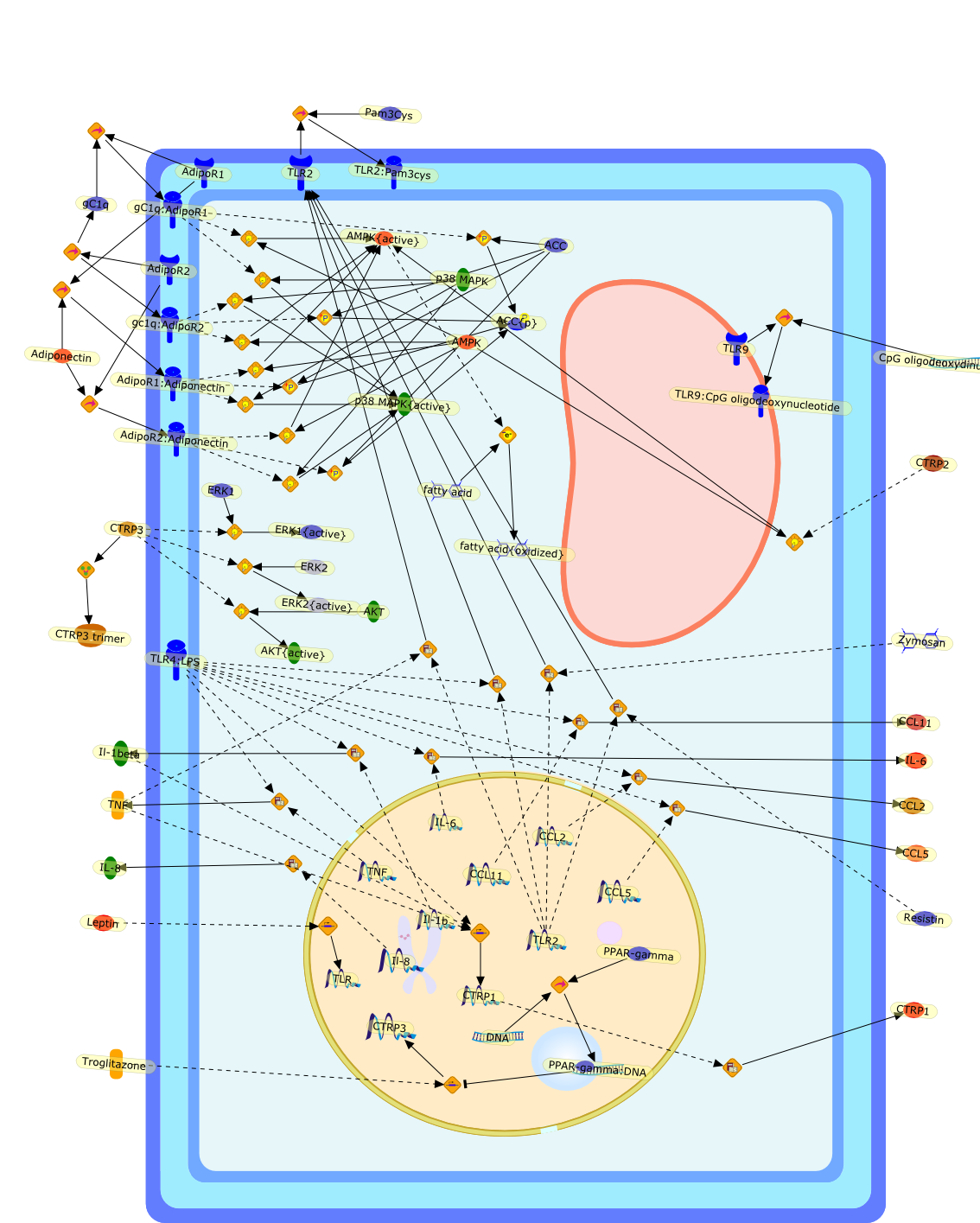{kind=link}
{kind=link}
ACC,ACC{p},AKT,AKT{active},AMPK,AMPK{active},AdipoR1,AdipoR1:Adiponectin,AdipoR2,AdipoR2:Adiponectin,Adiponectin,CCL11,CCL2,CCL5,CTRP1,CTRP2,CTRP3,CTRP3 trimer,CpG oligodeoxydinucleotide,DNA,ERK1,ERK1{active},ERK2,ERK2{active},IL-6,IL-8,Il-1b,Il-1beta,Il-8,Leptin,PPAR-gamma,PPAR-gamma:DNA,Pam3Cys,Resistin,TLR,TLR2,TLR2:Pam3cys,TLR4:LPS,TLR9,TLR9:CpG oligodeoxynucleotide,TNF,Troglitazone,Zymosan,fatty acid,fatty acid{oxidized},gC1q,gC1q:AdipoR1,gc1q:AdipoR2,p38 MAPK,p38 MAPK{active}
Dendritic cells (DCs) are powerful sensors of foreign pathogens as well ascancer cells and provide the first line of defence against infection. They alsoserve as a major link between innate and adaptive immunity. Immature DCs respondto incoming danger signals and undergo maturation to produce high levels ofproinflammatory cytokines including type I interferons (IFNs) to establishinnate immunity. They then present antigens to T lymphocytes to stimulatelasting specific immune responses. Recent studies point to the importance of DCsin the induction of peripheral tolerance. Transcription factors of the IRFfamily have emerged as crucial controllers of many aspects of DC activity,playing an essential role in the establishment of early innate immunity.Furthermore, eight of the nine members of the IRF family have been shown tocontrol either the differentiation and/or the functional activities of DCs. Inthis review, we focus on three aspects of DC properties that are under thecontrol of IRFs: (1) the development and differentiation, (2) maturation inresponse to toll-like receptor (TLR) signalling and the production ofanti-microbial cytokines, and (3) activation and expansion of lymphocytes togenerate protective or tolerogenic immune responses.
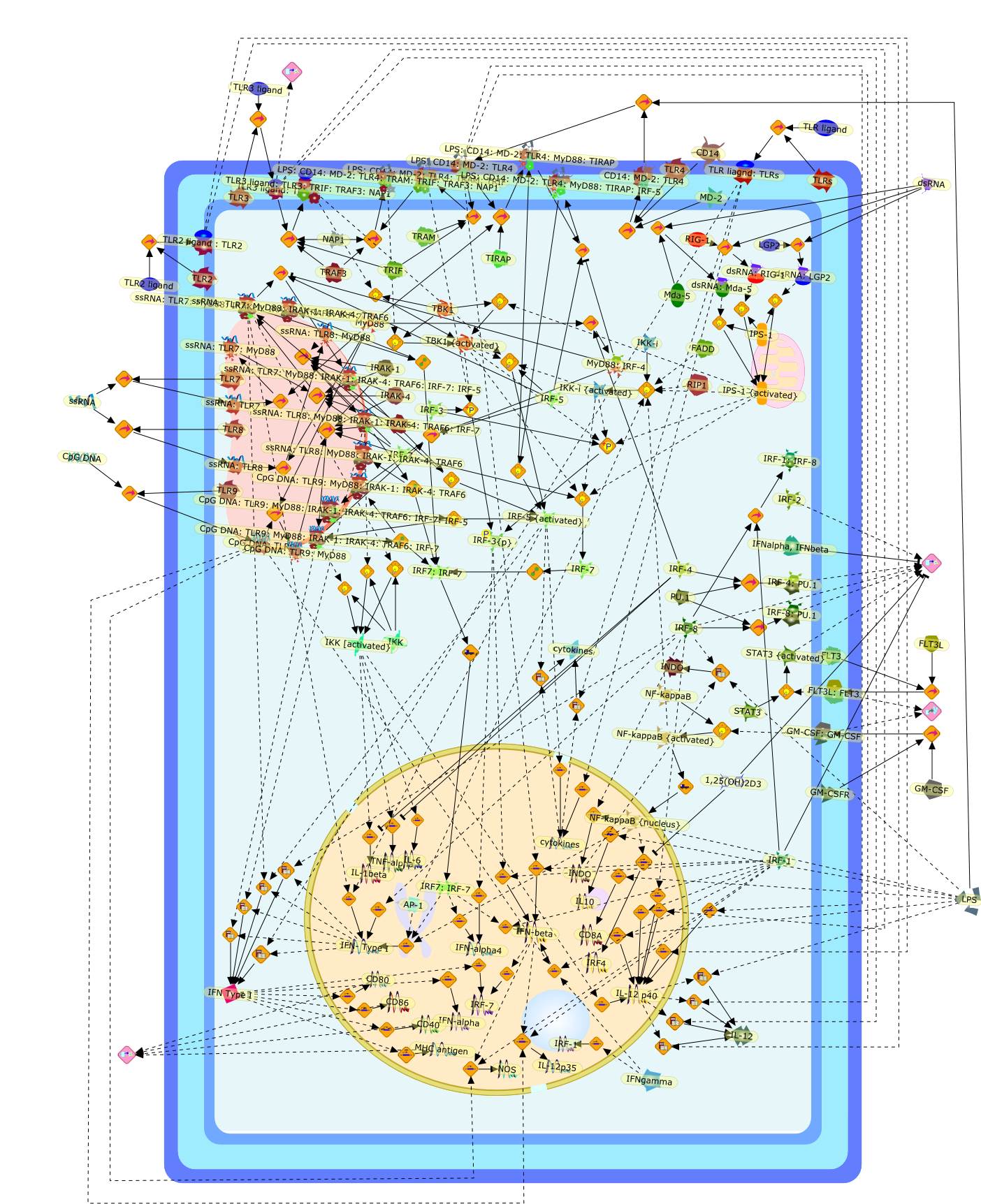{kind=link}
{kind=link}
1,25(OH)2D3,AP-1,CD14,CD14: MD-2: TLR4,CD40,CD80,CD86,CD8A,CpG DNA,CpG DNA: TLR9,CpG DNA: TLR9: MyD88,CpG DNA: TLR9: MyD88: IRAK-1: IRAK-4: TRAF6,CpG DNA: TLR9: MyD88: IRAK-1: IRAK-4: TRAF6: IRF-7,CpG DNA: TLR9: MyD88: IRAK-1: IRAK-4: TRAF6: IRF-7: IRF-5,FADD,FLT3,FLT3L,FLT3L: FLT3,GM-CSF,GM-CSF: GM-CSF,GM-CSFR,IFN Type I,IFN- Type I,IFN-alpha,IFN-alpha4,IFN-beta,IFNalpha, IFNbeta,IFNgamma,IKK,IKK [activated},IKK-i,IKK-i {activated},IL-12,IL-12 p40,IL-12p35,IL-1beta,IL-6,IL10,INDO,IPS-1,IPS-1 {activated},IRAK-1,IRAK-4,IRF-1,IRF-1: IRF-8,IRF-2,IRF-3,IRF-3{p},IRF-4,IRF-4: PU.1,IRF-5,IRF-5 {activated},IRF-7,IRF-8,IRF-8: PU.1,IRF4,IRF7: IRF-7,LGP2,LPS,LPS: CD14: MD-2: TLR4,LPS: CD14: MD-2: TLR4: MyD88: TIRAP,LPS: CD14: MD-2: TLR4: MyD88: TIRAP: IRF-5,LPS: CD14: MD-2: TLR4: TRAM: TRIF,LPS: CD14: MD-2: TLR4: TRAM: TRIF: TRAF3: NAP1,MD-2,MHC antigen,Mda-5,MyD88,MyD88: IRF-4,NAP1,NF-kappaB,NF-kappaB {activated},NF-kappaB {nucleus},NOS,PU.1,RIG-1,RIP1,STAT3,STAT3 {activated},TBK1,TBK1 {activated},TIRAP,TLR liagnd: TLRs,TLR ligand,TLR2,TLR2 ligand,TLR2 ligand : TLR2,TLR3,TLR3 ligand,TLR3 ligand: TLR3,TLR3 ligand: TLR3: TRIF: TRAF3: NAP1,TLR4,TLR7,TLR8,TLR9,TLRs,TNF-alpha,TRAF3,TRAF6,TRAM,TRIF,cytokines,dsRNA,dsRNA: LGP2,dsRNA: Mda-5,dsRNA: RIG-1,ssRNA,ssRNA: TLR7,ssRNA: TLR7: MyD88,ssRNA: TLR7: MyD88: IRAK-1: IRAK-4: TRAF6,ssRNA: TLR7: MyD88: IRAK-1: IRAK-4: TRAF6: IRF-7,ssRNA: TLR7: MyD88: IRAK-1: IRAK-4: TRAF6: IRF-7: IRF-5,ssRNA: TLR8,ssRNA: TLR8: MyD88,ssRNA: TLR8: MyD88: IRAK-1: IRAK-4: TRAF6,ssRNA: TLR8: MyD88: IRAK-1: IRAK-4: TRAF6: IRF-7
The type I interferon (IFN) alpha and beta promoters have been a leadingparadigm of virus-activated transcriptional regulation for more than twodecades, and have contributed substantially to our understanding ofvirus-inducible gene regulation, the coordinated activities of NF-kappaB and IRFtranscription factors, the temporal and spatial recruitment of co-activators tothe enhanceosome, and signaling pathways that trigger the innate antiviralresponse. In 2003, the ISICR Milstein Award was presented to John Hiscott ofMcGill University and Tom Maniatis of Harvard University for their ongoingresearch describing the mechanisms of regulation of type 1 interferon genes andspecifically for the identification of key signaling kinases involved inphosphorylation of the transcription factors IRF-3 and IRF-7. The specific rolesplayed by IRFs and the IKK-related kinases TBK1 and IKKvarepsilon are nowrecognized within the broader framework of TLR and RIG-I signaling pathways.This review summarizes the unique features of the IKK-related kinases and offersa summary of recent advances in the regulation of the early host response tovirus infection.
{kind=link}
{kind=link}
CBP,DDX58,DDX58 {ub}:dsRNA:Efp,DDX58:dsRNA,DDX58:dsRNA:Efp,DDX58{ub}:dsRNA:Efp:MAVS,DDX58{ub}:dsRNA:Efp:MAVS:TRAF3,DDX58{ub}:dsRNA:Efp:MAVS:TRAF3:IKK:TANK{p}:TBK1:IKK-i,DDX58{ub}:dsRNA:Efp:MAVS:TRAF3:IKK:TANK{p}:TBK1:IKK-i{active},DDX58{ub}:dsRNA:Efp:MAVS:TRAF6,DDX58{ub}:dsRNA:Efp:MAVS:TRAF6:IKK:TANK{p}:TBK1:IKK-i,DDX58{ub}:dsRNA:Efp:MAVS:TRAF6:IKK{active}:TANK{p}:TBK1:IKK-i,Efp,IFIH1,IFN-beta,IKK,IKK-alpha,IKK-beta,IKK-gamma,IKK-i,IKK:TANK{p}:TBK1:Ikk-i,IKappaB-alpha{p},IRF-3,IRF-3{p},IRF-3{p} dimer,IRF-3{p}dimer:CBP,IRF-7,IRF-7{active},MAVS,NAP1,NAP1:IKK-i:TBK1,NF-KappaB:IkappaB-alpha,NF-kappaB,PolyI:C,PolyI:C:IFIH1,STAT1,STAT1{p},TANK:TRAF2,TANK:TRAF2:TBK1:IKK-i,TANK{p}:TBK1:IKK-i,TANK{p}:TRAF2:TBK1:IKK-i,TBK1,TRAF2,TRAF3,TRAF6,dsRNA,proinflammatory genes
Recent advances in our understanding of the molecular basis of mammalian hostimmune responses to microbial invasion suggest that the first line of defenseagainst microbes is the recognition of pathogen-associated molecular patterns byToll-like receptors (TLRs). Phosphoinositide 3-kinase (PI3K) is thought toparticipate in the TLR signaling pathway. The activation of PI3K is commonlyobserved after stimulation with various TLR ligands. The resultant activation ofa serine-threonine protein kinase Akt leads to the phosphorylation of glycogensynthase kinase (GSK)-3beta, which is active in resting cells but is inactivatedby phosphorylation. GSK-3beta has been linked to the regulation of a multitudeof transcription factors, including NF-kappaB, AP-1, NF-AT, and CREB eithernegatively or positively. Thus, the altered activity of GSK-3beta causes diverseeffects on cytokine expression. Generally, activation of PI3K results in theinhibition of proinflammatory events such as expression of IL-12 and TNF-alpha.Thus, PI3K is a negative regulator of TLR signaling. Among the members of theClass I PI3K family, p85/p110beta appears to be the subtype activated on TLRligation, but the molecular basis for this specificity has yet to be elucidated.
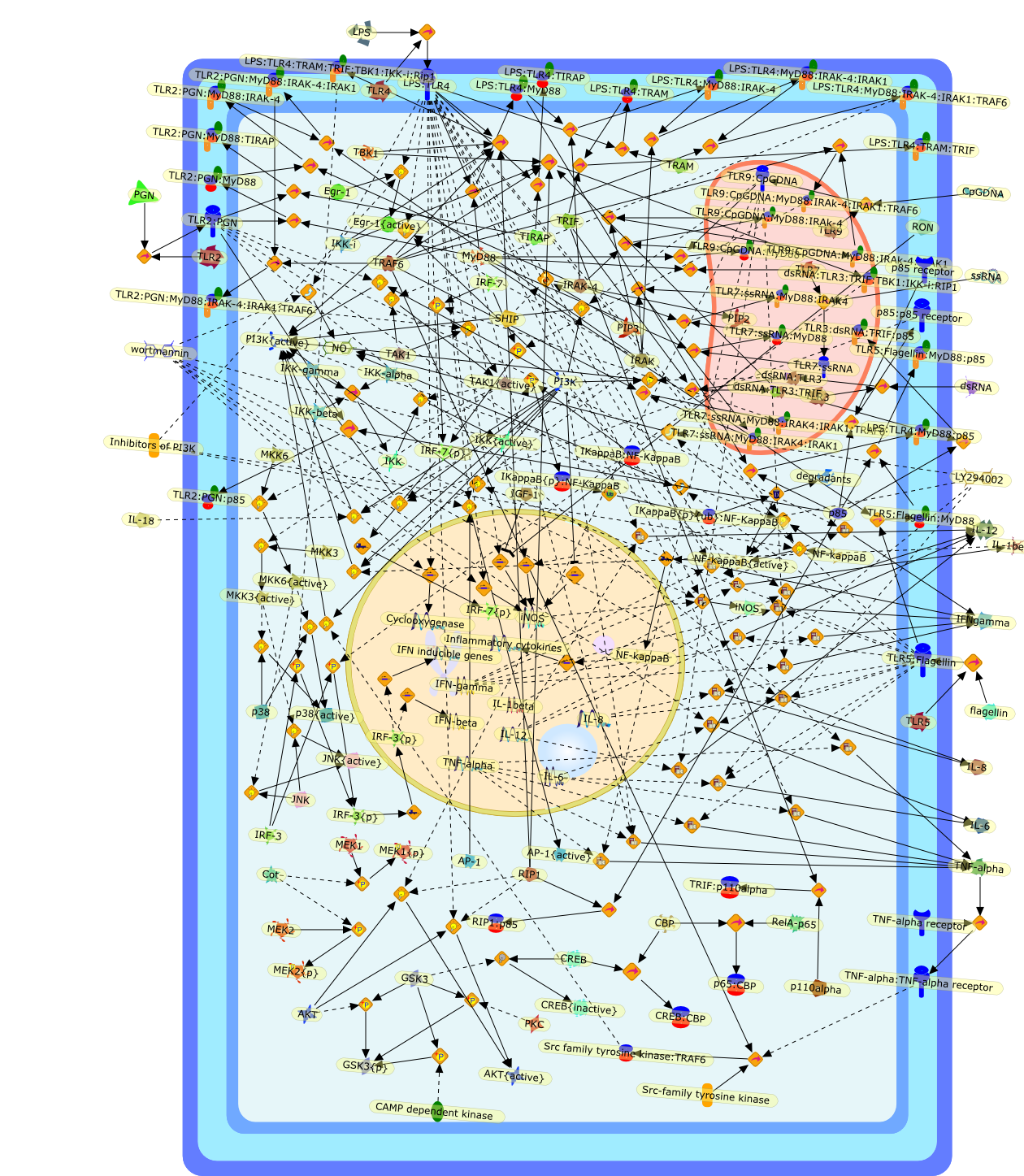{kind=link}
{kind=link}
AKT,AKT{active},AP-1,AP-1{active},CAMP dependent kinase,CBP,CREB,CREB:CBP,CREB{inactive},Cot,CpGDNA,Cyclooxygenase,Egr-1,Egr-1{active},GSK3,GSK3{p},IFN inducible genes,IFN-beta,IFN-gamma,IFNgamma,IGF-1,IKK,IKK-alpha,IKK-beta,IKK-gamma,IKK-i,IKK{active},IKappaB:NF-KappaB,IKappaB{p}:NF-KappaB,IKappaB{p}{ub}:NF-KappaB,IL-12,IL-18,IL-1beta,IL-6,IL-8,IRAK,IRAK-4,IRF-3,IRF-3{p},IRF-7,IRF-7{p},Inflammatory cytokines,Inhibitors of PI3K,JNK,JNK{active},LPS,LPS:TLR4,LPS:TLR4:MyD88,LPS:TLR4:MyD88:IRAK-4,LPS:TLR4:MyD88:IRAK-4:IRAK1,LPS:TLR4:MyD88:IRAK-4:IRAK1:TRAF6,LPS:TLR4:MyD88:p85,LPS:TLR4:TIRAP,LPS:TLR4:TRAM,LPS:TLR4:TRAM:TRIF,LPS:TLR4:TRAM:TRIF:TBK1:IKK-i:Rip1,LY294002,MEK1,MEK1{p},MEK2,MEK2{p},MKK3,MKK3{active},MKK6,MKK6{active},MyD88,NF-kappaB,NF-kappaB{active},NO,PGN,PI3K,PI3K{active},PIP2,PIP3,PKC,RIP1,RIP1:p85,RON,RelA-p65,SHIP,Src family tyrosine kinase:TRAF6,Src-family tyrosine kinase,TAK1,TAK1{active},TBK1,TIRAP,TLR2,TLR2:PGN,TLR2:PGN:MyD88,TLR2:PGN:MyD88:IRAK-4,TLR2:PGN:MyD88:IRAK-4:IRAK1,TLR2:PGN:MyD88:IRAK-4:IRAK1:TRAF6,TLR2:PGN:MyD88:TIRAP,TLR2:PGN:p85,TLR3,TLR3:dsRNA:TRIF:p85,TLR4,TLR5,TLR5:Flagellin,TLR5:Flagellin:MyD88,TLR5:Flagellin:MyD88:p85,TLR7,TLR7:ssRNA,TLR7:ssRNA:MyD88,TLR7:ssRNA:MyD88:IRAK4,TLR7:ssRNA:MyD88:IRAK4:IRAK1,TLR7:ssRNA:MyD88:IRAK4:IRAK1:TRAF6,TLR9,TLR9:CpGDNA,TLR9:CpGDNA:MyD88,TLR9:CpGDNA:MyD88:IRAk-4,TLR9:CpGDNA:MyD88:IRAk-4:IRAK1,TLR9:CpGDNA:MyD88:IRAk-4:IRAK1:TRAF6,TNF-alpha,TNF-alpha receptor,TNF-alpha:TNF-alpha receptor,TRAF6,TRAM,TRIF,TRIF:p110alpha,degradants,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:TBK1:IKK-i:RIP1,flagellin,iNOS,p110alpha,p38,p38{active},p65:CBP,p85,p85 receptor,p85:p85 receptor,ssRNA,wortmannin
The development of autoimmunity is often associated with the presence ofpathogen-associated molecular patterns (PAMPs) and signaling through toll-likereceptors (TLRs). Largely, the importance of PAMP-TLR ligation has beenattributed to inducing the maturation of antigen-presenting cells and productionof proinflammatory cytokines and chemokines. Recent evidence now shows thatPAMPs can activate effector and regulatory T cells revealing a further level ofcomplexity in the development of autoimmunity. TLR signaling on T cells acts asa form of costimulation, lowering the 'strength of signal' required forproliferation and survival. This apparent mechanism of immune homeostasis maybreak tolerance or anergy upon pathogen infection and promote the development ofimmune responses against self-antigens.
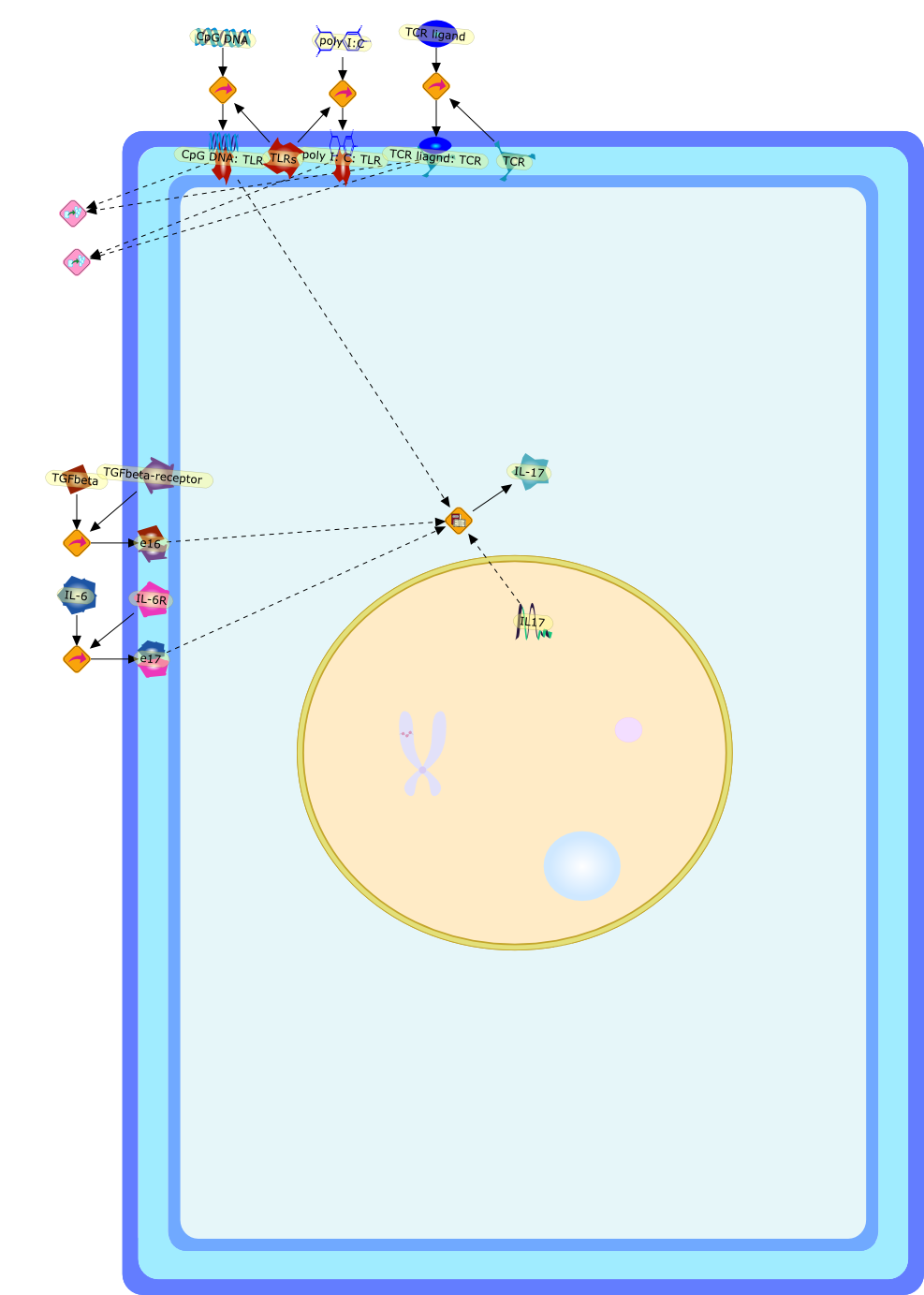{kind=link}
{kind=link}
CpG DNA,CpG DNA: TLR,IL-17,IL-6,IL-6R,IL17,TCR,TCR liagnd: TCR,TCR ligand,TGFbeta,TGFbeta-receptor,TLRs,e16,e17,poly I: C: TLR,poly I:C
The innate immune system is equipped with sensitive and efficient machineries toprovide an immediate, first line defense against infections. Toll-like receptors(TLRs) detect pathogens and the IL-1 receptor (IL-1R) family enables cells toquickly respond to inflammatory cytokines by mounting an efficient protectiveresponse. Interleukin-1 receptor activated kinases (IRAKs) are key mediators inthe signaling pathways of TLRs/IL-1Rs. By means of their kinase and adaptorfunctions, IRAKs initiate a cascade of signaling events eventually leading toinduction of inflammatory target gene expression. Due to this pivotal role, IRAKfunction is also highly regulated via multiple mechanisms. In this review, wefocus on IRAK1, the earliest known and yet the most interesting member of thisfamily. An overview on its structure, function and biology is given, withemphasis on the different novel mechanisms that regulate IRAK1 function. We alsohighlight several unresolved questions in this field and evaluate the potentialof IRAK1 as a target for therapeutic intervention.
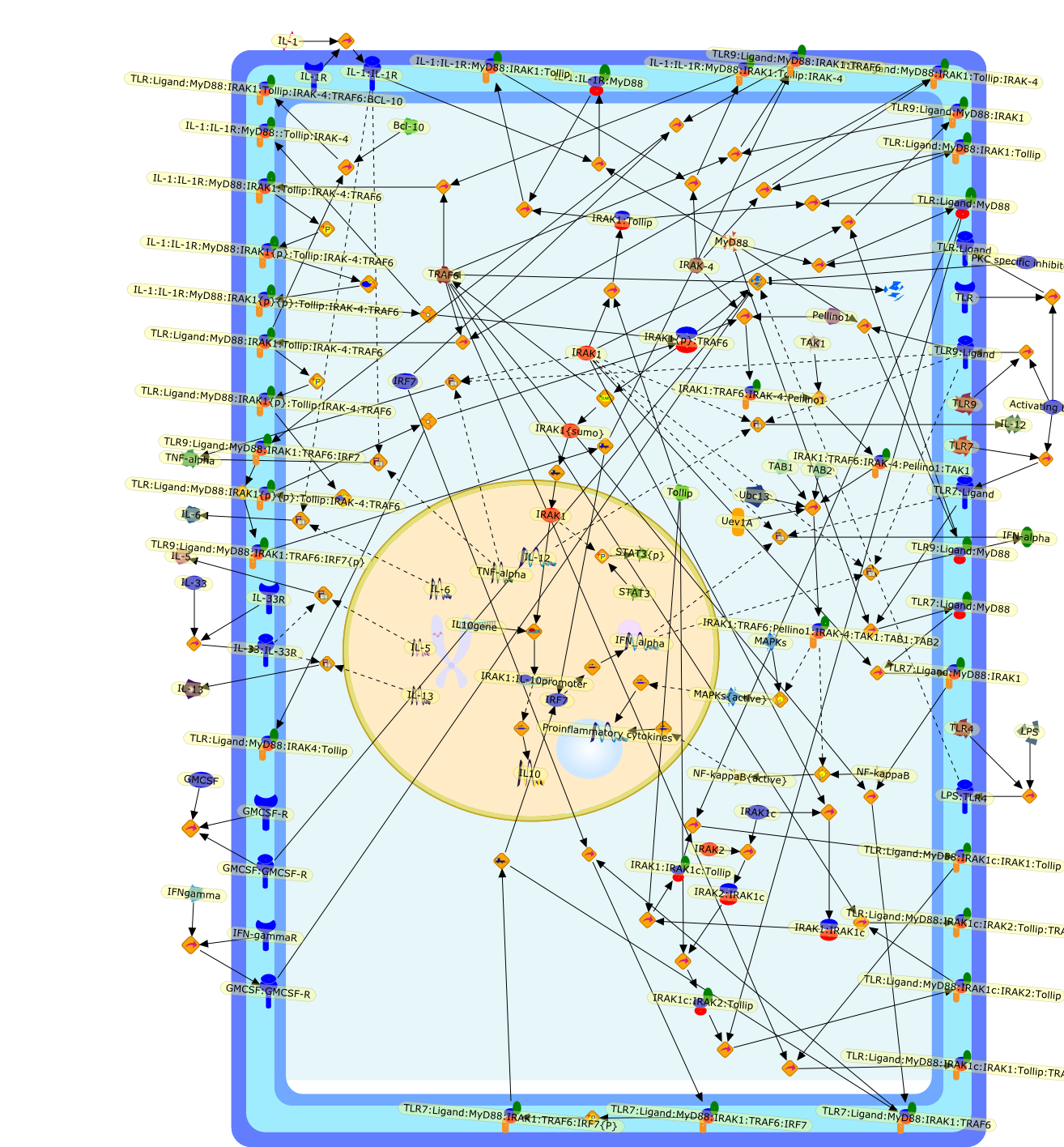{kind=link}
{kind=link}
Activating Ligand,Bcl-10,GMCSF,GMCSF-R,GMCSF:GMCSF-R,IFN-alpha,IFN-gammaR,IFN_alpha,IFNgamma,IL-1,IL-12,IL-13,IL-1:IL-1R,IL-1:IL-1R:MyD88,IL-1:IL-1R:MyD88::Tollip:IRAK-4,IL-1:IL-1R:MyD88:IRAK1:Tollip,IL-1:IL-1R:MyD88:IRAK1:Tollip:IRAK-4,IL-1:IL-1R:MyD88:IRAK1:Tollip:IRAK-4:TRAF6,IL-1:IL-1R:MyD88:IRAK1{p}:Tollip:IRAK-4:TRAF6,IL-1:IL-1R:MyD88:IRAK1{p}{p}:Tollip:IRAK-4:TRAF6,IL-1R,IL-33,IL-33:IL-33R,IL-33R,IL-5,IL-6,IL10,IL10gene,IRAK-4,IRAK1,IRAK1:IL-10promoter,IRAK1:IRAK1c,IRAK1:IRAK1c:Tollip,IRAK1:TRAF6:IRAK-4:Pellino1,IRAK1:TRAF6:IRAK-4:Pellino1:TAK1,IRAK1:TRAF6:Pellino1:IRAK-4:TAK1:TAB1:TAB2,IRAK1:Tollip,IRAK1c,IRAK1c:IRAK2:Tollip,IRAK1{p}:TRAF6,IRAK1{sumo},IRAK2,IRAK2:IRAK1c,IRF7,LPS,LPS:TLR4,MAPKs,MAPKs{active},MyD88,NF-kappaB,NF-kappaB{active},PKC specific inhibitors,Pellino1,Proinflammatory cytokines,STAT3,STAT3{p},TAB1,TAB2,TAK1,TLR,TLR4,TLR7,TLR7:Ligand,TLR7:Ligand:MyD88,TLR7:Ligand:MyD88:IRAK1,TLR7:Ligand:MyD88:IRAK1:TRAF6,TLR7:Ligand:MyD88:IRAK1:TRAF6:IRF7,TLR7:Ligand:MyD88:IRAK1:TRAF6:IRF7{P},TLR9,TLR9:Ligand,TLR9:Ligand:MyD88,TLR9:Ligand:MyD88:IRAK1,TLR9:Ligand:MyD88:IRAK1:TRAF6,TLR9:Ligand:MyD88:IRAK1:TRAF6:IRF7,TLR9:Ligand:MyD88:IRAK1:TRAF6:IRF7{p},TLR:Ligand,TLR:Ligand:MyD88,TLR:Ligand:MyD88:IRAK1:Tollip,TLR:Ligand:MyD88:IRAK1:Tollip:IRAK-4,TLR:Ligand:MyD88:IRAK1:Tollip:IRAK-4:TRAF6,TLR:Ligand:MyD88:IRAK1:Tollip:IRAK-4:TRAF6:BCL-10,TLR:Ligand:MyD88:IRAK1c:IRAK1:Tollip,TLR:Ligand:MyD88:IRAK1c:IRAK1:Tollip:TRAF6,TLR:Ligand:MyD88:IRAK1c:IRAK2:Tollip,TLR:Ligand:MyD88:IRAK1c:IRAK2:Tollip:TRAF6,TLR:Ligand:MyD88:IRAK1{p}:Tollip:IRAK-4:TRAF6,TLR:Ligand:MyD88:IRAK1{p}{p}:Tollip:IRAK-4:TRAF6,TLR:Ligand:MyD88:IRAK4:Tollip,TNF-alpha,TRAF6,Tollip,Ubc13,Uev1A,degradants
Type I interferon-alpha/beta play an essential role in immunity to viruses.While interferon-beta has been used as a model of a complex promoter, many ofthe signalling pathways leading to interferon-beta gene expression remaincontroversial. Recent milestones include the discovery of Toll-like receptorsand RNA helicases that signal via a novel kinase complex composed of I kappa Bkinase-iota/epsilon or TANK binding kinase-1. This review provides a timelysummary of this rapidly expanding field, focusing specifically on the variousviral RNA binding molecules and their associated signalling pathways.
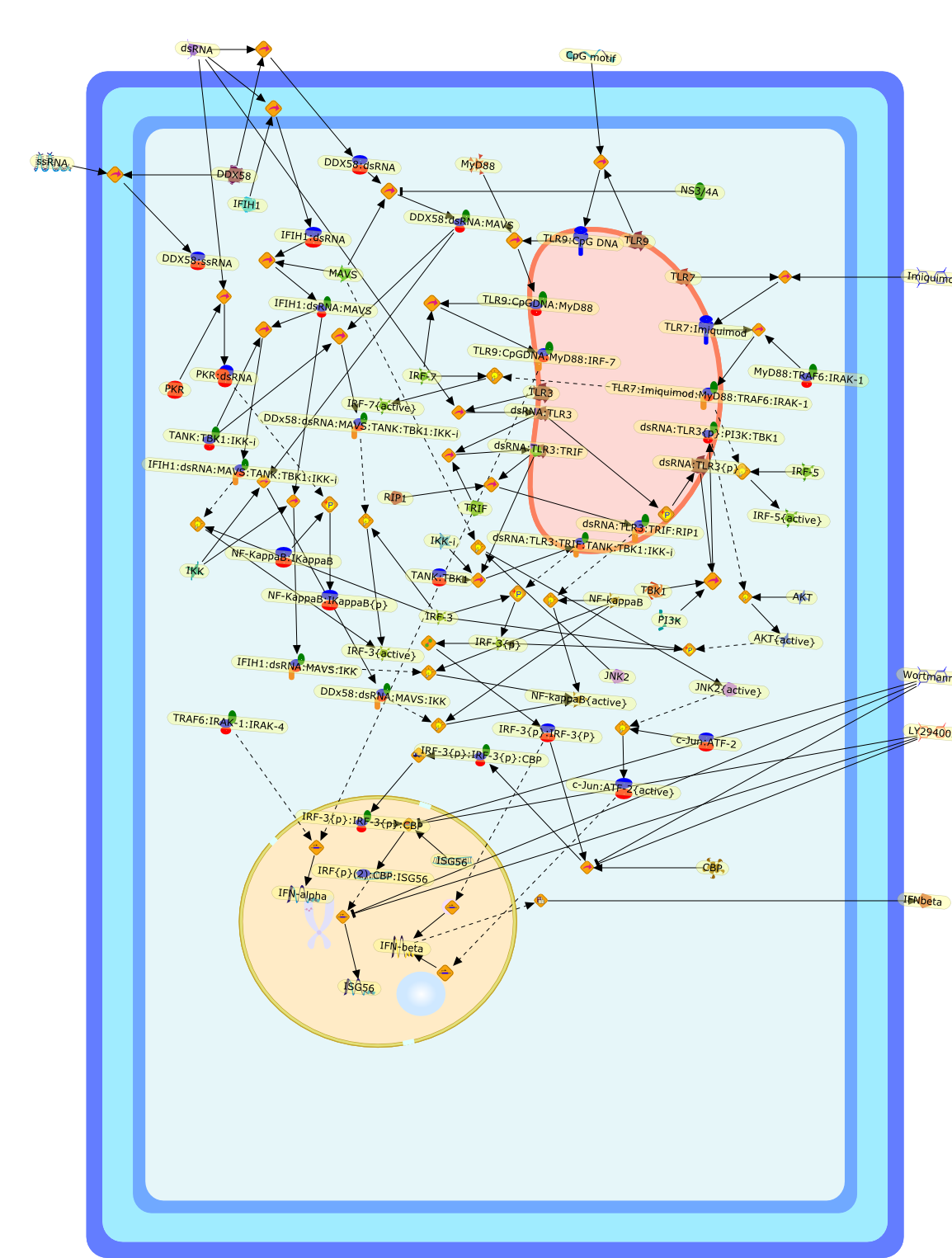{kind=link}
{kind=link}
AKT,AKT{active},CBP,CpG motif,DDX58,DDX58:dsRNA,DDX58:dsRNA:MAVS,DDX58:ssRNA,DDx58:dsRNA:MAVS:IKK,DDx58:dsRNA:MAVS:TANK:TBK1:IKK-i,IFIH1,IFIH1:dsRNA,IFIH1:dsRNA:MAVS,IFIH1:dsRNA:MAVS:IKK,IFIH1:dsRNA:MAVS:TANK:TBK1:IKK-i,IFN-alpha,IFN-beta,IFNbeta,IKK,IKK-i,IRF-3,IRF-3{active},IRF-3{p},IRF-3{p}:IRF-3{P},IRF-3{p}:IRF-3{p}:CBP,IRF-5,IRF-5{active},IRF-7,IRF-7{active},IRF{p}(2):CBP:ISG56,ISG56,Imiquimod,JNK2,JNK2{active},LY294002,MAVS,MyD88,MyD88:TRAF6:IRAK-1,NF-KappaB:IKappaB,NF-KappaB:IKappaB{p},NF-kappaB,NF-kappaB{active},NS3/4A,PI3K,PKR,PKR:dsRNA,RIP1,TANK:TBK1,TANK:TBK1:IKK-i,TBK1,TLR3,TLR7,TLR7:Imiquimod,TLR7:Imiquimod:MyD88:TRAF6:IRAK-1,TLR9,TLR9:CpG DNA,TLR9:CpGDNA:MyD88,TLR9:CpGDNA:MyD88:IRF-7,TRAF6:IRAK-1:IRAK-4,TRIF,Wortmannin,c-Jun:ATF-2,c-Jun:ATF-2{active},dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:RIP1,dsRNA:TLR3:TRIF:TANK:TBK1:IKK-i,dsRNA:TLR3{p},dsRNA:TLR3{p}:PI3K:TBK1,ssRNA
Several cysteine proteases of the caspase family play a central role in manyforms of cell death by apoptosis. Other enzymes of the family are involved incytokine maturation along inflammatory response. In recent years, severalcaspases involved in cell death were shown to play a role in other cellularprocesses such as proliferation and differentiation. In the present review, wesummarize the current knowledge of the role of caspases in the differentiationof erythroid cells and macrophages. Based on these two examples, we show thatthe nature of involved enzymes, the pathways leading to their activation inresponse to specific growth factors, and the specificity of the target proteinsthat are cleaved by the activated enzymes strongly differ from one cell type toanother. Deregulation of these pathways is thought to play a role in thepathophysiology of low-grade myelodysplastic syndromes, characterized byexcessive activation of caspases and erythroid precursor apoptosis, and that ofchronic myelomonocytic leukemia, characterized by a defective activation ofcaspases in monocytes exposed to M-CSF, which blocks their differentiation.
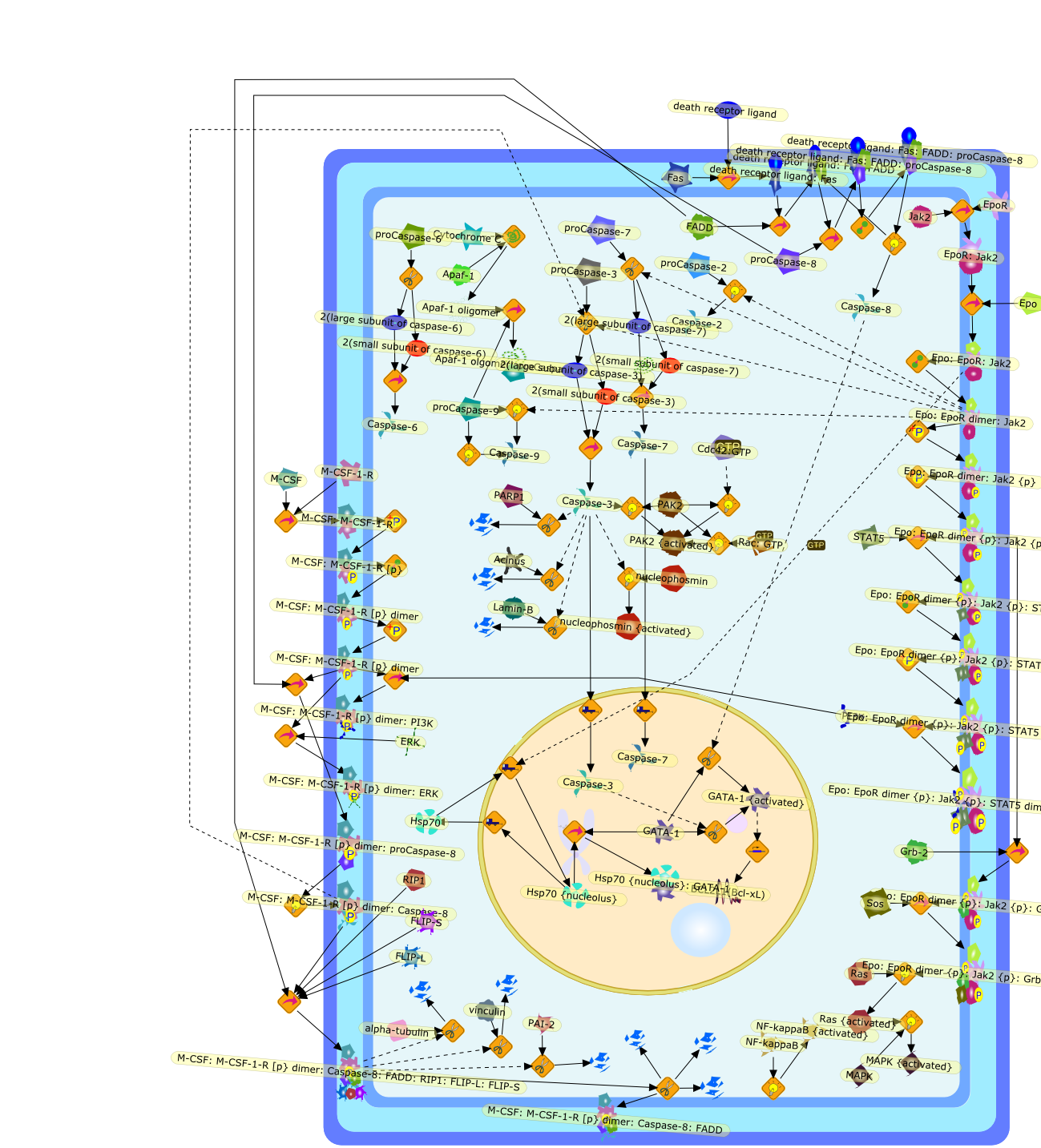{kind=link}
{kind=link}
2(large subunit of caspase-3),2(large subunit of caspase-6),2(large subunit of caspase-7),2(small subunit of caspase-3),2(small subunit of caspase-6),2(small subunit of caspase-7),Acinus,Acinus remnants,Apaf-1,Apaf-1 olgomer: proCaspase-9,Apaf-1 oligomer,BCL2L1(Bcl-xL),Caspase-2,Caspase-3,Caspase-6,Caspase-7,Caspase-8,Caspase-9,Cdc42:GTP,Cytochrome C,ERK,Epo,Epo: EpoR dimer {p}: Jak2 {p},Epo: EpoR dimer {p}: Jak2 {p}: Grb-2,Epo: EpoR dimer {p}: Jak2 {p}: Grb-2: Sos,Epo: EpoR dimer {p}: Jak2 {p}: STAT5,Epo: EpoR dimer {p}: Jak2 {p}: STAT5 dimer,Epo: EpoR dimer {p}: Jak2 {p}: STAT5 dimer {p},Epo: EpoR dimer {p}: Jak2 {p}: STAT5 dimer {p}: PI3K,Epo: EpoR dimer: Jak2,Epo: EpoR dimer: Jak2 {p},Epo: EpoR: Jak2,EpoR,EpoR: Jak2,FADD,FLIP-L,FLIP-L remnant,FLIP-S,FLIP-S remnant,Fas,GATA-1,GATA-1 {activated},Grb-2,Hsp70,Hsp70 {nucleolus},Hsp70 {nucleolus}: GATA-1,Jak2,Lamin-B,Lamin-B remnants,M-CSF,M-CSF-1-R,M-CSF: M-CSF-1-R,M-CSF: M-CSF-1-R [p},M-CSF: M-CSF-1-R [p} dimer,M-CSF: M-CSF-1-R [p} dimer: Caspase-8,M-CSF: M-CSF-1-R [p} dimer: Caspase-8: FADD,M-CSF: M-CSF-1-R [p} dimer: Caspase-8: FADD: RIP1: FLIP-L: FLIP-S,M-CSF: M-CSF-1-R [p} dimer: ERK,M-CSF: M-CSF-1-R [p} dimer: PI3K,M-CSF: M-CSF-1-R [p} dimer: proCaspase-8,MAPK,MAPK {activated},NF-kappaB,NF-kappaB {activated},PAI-2,PAI-2 remnants,PAK2,PAK2 {activated},PARP1,PARP1 remnants,PI3K,RIP1,RIP1 fragment,Rac: GTP,Ras,Ras {activated},STAT5,Sos,alpha-tubulin,alpha-tubulin remnants,death receptor ligand,death receptor ligand: Fas,death receptor ligand: Fas: FADD,death receptor ligand: Fas: FADD: proCaspase-8,nucleophosmin,nucleophosmin {activated},proCaspase-2,proCaspase-3,proCaspase-6,proCaspase-7,proCaspase-8,proCaspase-9,vinculin,vinculin remnants
Although adhesion to extracellular structures is one of the most fundamentalcell biological processes, the intracellular signals triggered by integrins, themost important receptors involved, are incompletely understood. Several recentreports indicate that signaling by beta(2) and beta(3) integrins in various celltypes (neutrophils, macrophages, osteoclasts and platelets) use components ofthe signal transduction machinery of lymphocyte antigen receptors. Central tothis immunoreceptor-like signaling is the phosphorylation of immunoreceptortyrosine-based activation motif (ITAM)-containing adapters (such as DAP12 andthe Fc receptor gamma-chain) by Src-family kinases and the concomitantrecruitment of the Syk tyrosine kinase through its dual SH2 domains. These andother reports reveal an unexpected similarity between the signal-transductionmechanisms used by integrins and immune recognition receptors.
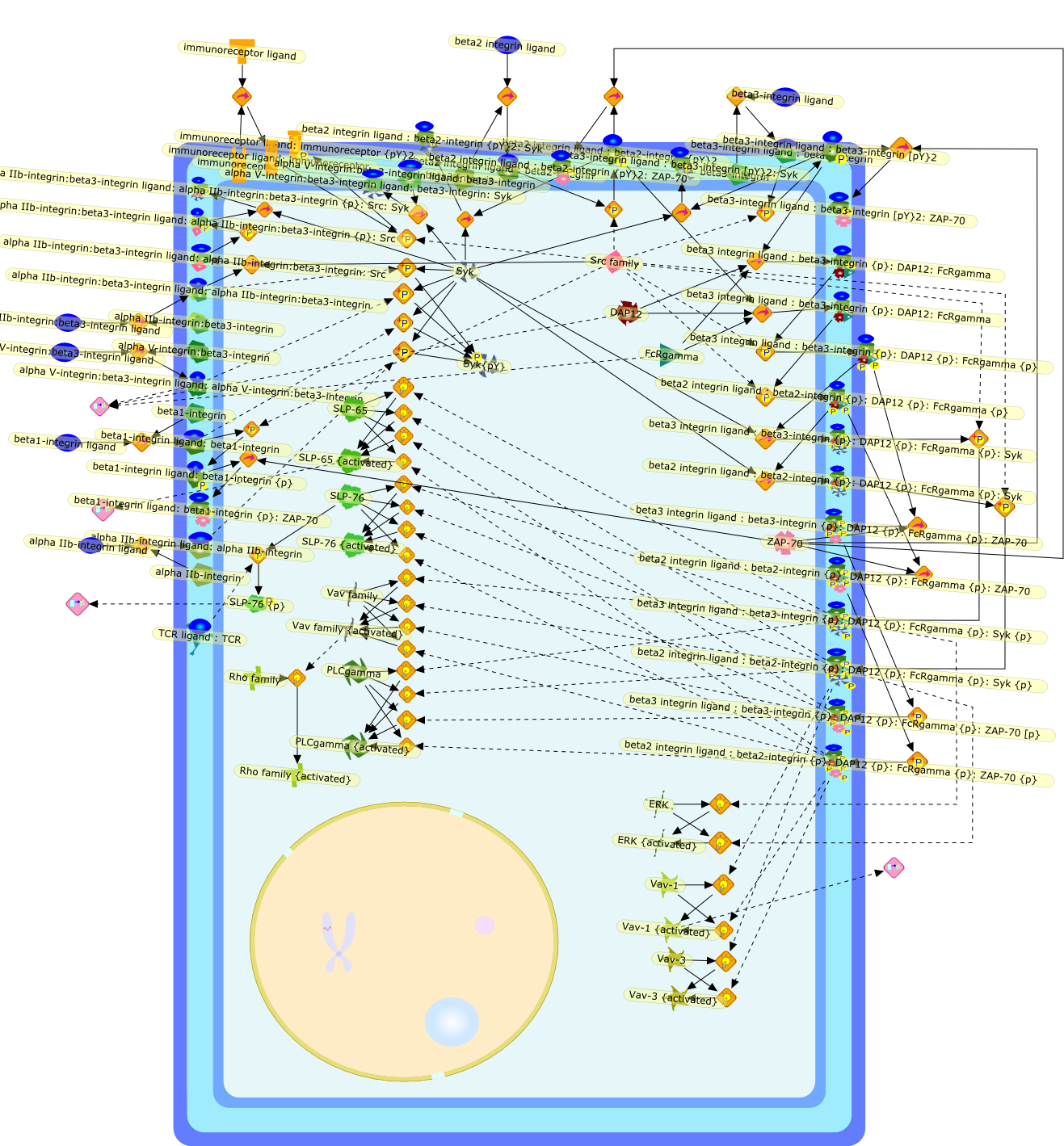{kind=link}
{kind=link}
DAP12,ERK,ERK {activated},FcRgamma,PLCgamma,PLCgamma {activated},Rho family,Rho family {activated},SLP-65,SLP-65 {activated},SLP-76,SLP-76 {activated},SLP-76 {p},Src family,Syk,Syk{pY},TCR ligand : TCR,Vav family,Vav family {activated},Vav-1,Vav-1 {activated},Vav-3,Vav-3 {activated},ZAP-70,alpha IIb-integrin,alpha IIb-integrin ligand,alpha IIb-integrin ligand: alpha IIb-integrin,alpha IIb-integrin:beta3-integrin,alpha IIb-integrin:beta3-integrin ligand,alpha IIb-integrin:beta3-integrin ligand: alpha IIb-integrin:beta3-integrin,alpha IIb-integrin:beta3-integrin ligand: alpha IIb-integrin:beta3-integrin {p}: Src,alpha IIb-integrin:beta3-integrin ligand: alpha IIb-integrin:beta3-integrin {p}: Src: Syk,alpha IIb-integrin:beta3-integrin ligand: alpha IIb-integrin:beta3-integrin: Src,alpha V-integrin:beta3-integrin,alpha V-integrin:beta3-integrin ligand,alpha V-integrin:beta3-integrin ligand: alpha V-integrin:beta3-integrin,alpha V-integrin:beta3-integrin ligand: beta3-integrin,alpha V-integrin:beta3-integrin ligand: beta3-integrin: Syk,beta1-integrin,beta1-integrin ligand,beta1-integrin ligand: beta1-integrin,beta1-integrin ligand: beta1-integrin {p},beta1-integrin ligand: beta1-integrin {p}: ZAP-70,beta2 integrin ligand,beta2 integrin ligand : beta2-integrin,beta2 integrin ligand : beta2-integrin {pY}2,beta2 integrin ligand : beta2-integrin {pY}2: Syk,beta2 integrin ligand : beta2-integrin {pY}2: ZAP-70,beta2 integrin ligand : beta2-integrin {p}: DAP12 {p}: FcRgamma {p},beta2 integrin ligand : beta2-integrin {p}: DAP12 {p}: FcRgamma {p}: Syk,beta2 integrin ligand : beta2-integrin {p}: DAP12 {p}: FcRgamma {p}: Syk {p},beta2 integrin ligand : beta2-integrin {p}: DAP12 {p}: FcRgamma {p}: ZAP-70,beta2 integrin ligand : beta2-integrin {p}: DAP12 {p}: FcRgamma {p}: ZAP-70 {p},beta2-integrin,beta3 integrin ligand : beta3-integrin {p}: DAP12 {p}: FcRgamma {p},beta3 integrin ligand : beta3-integrin {p}: DAP12 {p}: FcRgamma {p}: Syk,beta3 integrin ligand : beta3-integrin {p}: DAP12 {p}: FcRgamma {p}: Syk {p},beta3 integrin ligand : beta3-integrin {p}: DAP12 {p}: FcRgamma {p}: ZAP-70,beta3 integrin ligand : beta3-integrin {p}: DAP12 {p}: FcRgamma {p}: ZAP-70 [p},beta3 integrin ligand : beta3-integrin {p}: DAP12: FcRgamma,beta3-integrin,beta3-integrin ligand,beta3-integrin ligand : beta3-integrin,beta3-integrin ligand : beta3-integrin [pY}2,beta3-integrin ligand : beta3-integrin [pY}2: Syk,beta3-integrin ligand : beta3-integrin [pY}2: ZAP-70,immunoreceptor,immunoreceptor ligand,immunoreceptor ligand: immunoreceptor,immunoreceptor ligand: immunoreceptor {pY}2
Toll-like receptors (TLRs) are the archetypal pattern recognition receptors insensing exogenous pathogens. Activation of TLRs is a first line of defense ofthe immune system, leading to the activation and recruitment of neutrophils andmacrophages to sites of infection and enhances antimicrobial activity. The TLRsignaling through different intracellular molecules, such as MAP kinases andIkappaB kinases which are conserved signaling elements for many receptors, leadsto a distinct set of proinflammatory gene expressions. However, how thesepathways differentially and precisely control the transcription of identicalgenes remains largely unknown. Our review focuses on the details of up-to-datesignaling molecules including negative regulators and their role in controllinginnate immune response. We also stress the importance of developing systemicapproaches for the global understanding of TLR signaling so that appropriatedrug therapeutic targets can be identified for regulating inflammatory diseases.
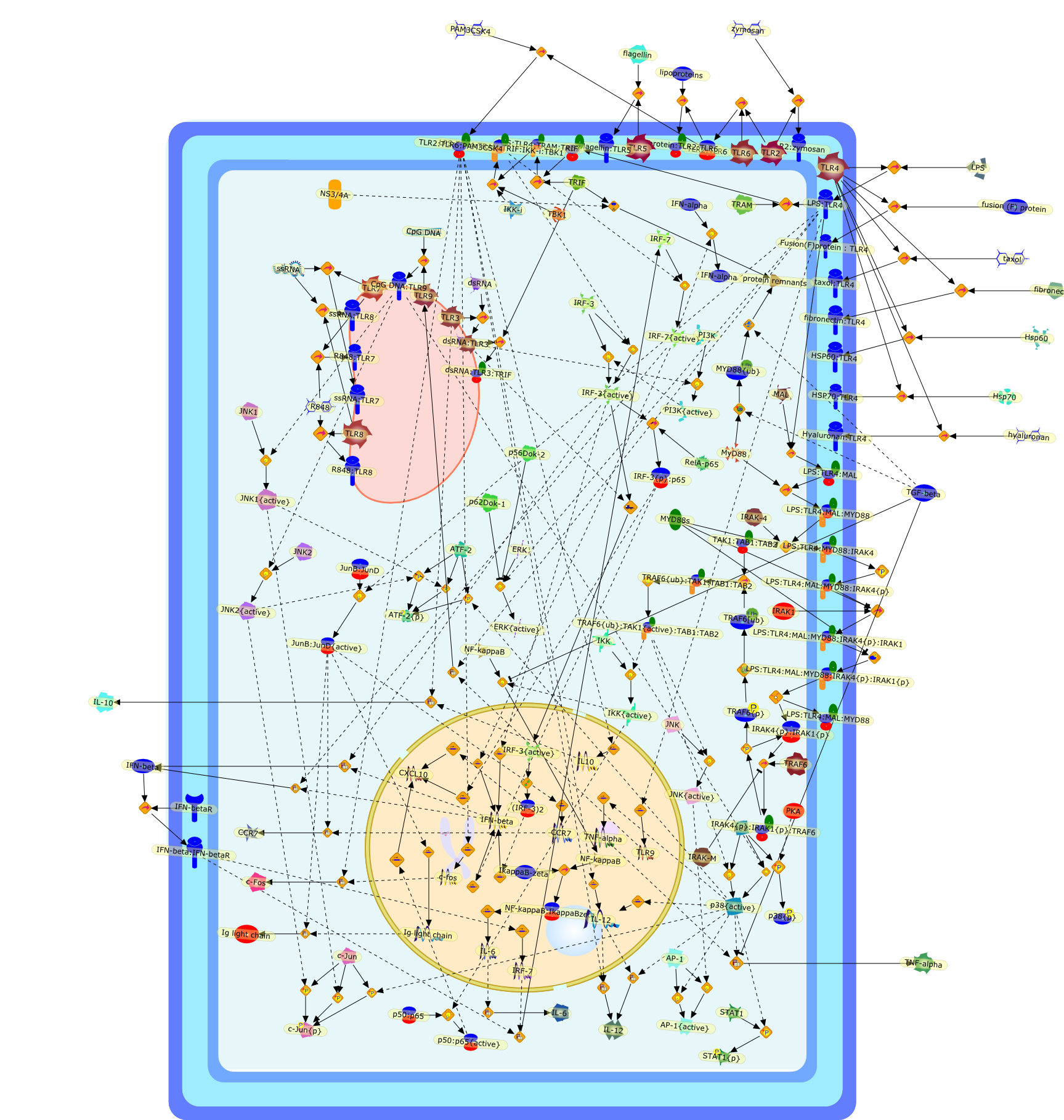{kind=link}
{kind=link}
(IRF-3)2,AP-1,AP-1{active},ATF-2,ATF-2{p},CCR7,CXCL10,CpG DNA,CpG DNA:TLR9,ERK,ERK{active},Fusion(F)protein : TLR4,HSP60:TLR4,HSP70:TLR4,Hsp60,Hsp70,Hyaluronan:TLR4,IFN-alpha,IFN-beta,IFN-beta:IFN-betaR,IFN-betaR,IKK,IKK-i,IKK{active},IL-10,IL-12,IL-6,IL10,IRAK-4,IRAK-M,IRAK1,IRAK4{p}:IRAK1{p},IRAK4{p}:IRAK1{p}:TRAF6,IRF-3,IRF-3{active},IRF-3{p}:p65,IRF-7,IRF-7{active},Ig light chain,IkappaB-zeta,JNK,JNK1,JNK1{active},JNK2,JNK2{active},JNK{active},JunB:JunD,JunB:JunD{active},LPS,LPS:TLR4,LPS:TLR4:MAL,LPS:TLR4:MAL:MYD88,LPS:TLR4:MAL:MYD88:IRAK4{p},LPS:TLR4:MAL:MYD88:IRAK4{p}:IRAK1,LPS:TLR4:MAL:MYD88:IRAK4{p}:IRAK1{p},LPS:TLR4:MYD88:IRAK4,LPS:TLR4:TRAM:TRIF,LPS:TLR4:TRAM:TRIF:IKK-i:TBK1,MAL,MYD88s,MYD88{ub},MyD88,NF-kappaB,NF-kappaB:IkappaBzeta,NS3/4A,PAM3CSK4,PI3K,PI3K{active},PKA,R848,R848:TLR7,R848:TLR8,RelA-p65,STAT1,STAT1{p},TAK1:TAB1:TAB2,TBK1,TGF-beta,TLR2,TLR2:TLR6,TLR2:TLR6:PAM3CSK4,TLR2:zymosan,TLR3,TLR4,TLR5,TLR6,TLR7,TLR8,TLR9,TNF-alpha,TRAF6,TRAF6[ub},TRAF6{p},TRAF6{ub}:TAK1:TAB1:TAB2,TRAF6{ub}:TAK1{active}:TAB1:TAB2,TRAM,TRIF,c-Fos,c-Jun,c-Jun{p},c-fos,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,e67,fibronectin,fibronectin:TLR4,flagellin,flagellin:TLR5,fusion (F) protein,hyaluronan,lipoprotein:TLR2:TLR6,lipoproteins,p38,p38{active},p38{p},p50:p65,p50:p65{active},p56Dok-2,p62Dok-1,protein remnants,ssRNA,ssRNA:TLR7,ssRNA:TLR8,taxol,taxol:TLR4,zymosan
There is a constant interplay between the innate and adaptive immune systems,which leads to a protective immune response against pathogens and contributeseffectively to self-non-self discrimination. Toll-like receptors (TLRs) are keycomponents of the innate immune system, which activate multiple inflammatorypathways and coordinate systemic defense against pathogens. In addition torecognizing unique molecular patterns associated with different classes ofpathogens, TLRs may also recognize a number of self proteins and endogenousnucleic acids. Data originating predominantly from animal models of autoimmunedisease and circumstantial data from human patients suggest that inappropriateactivation of TLR pathways by endogenous or exogenous ligands may lead to theinitiation and/or perpetuation of autoimmune responses and tissue injury.
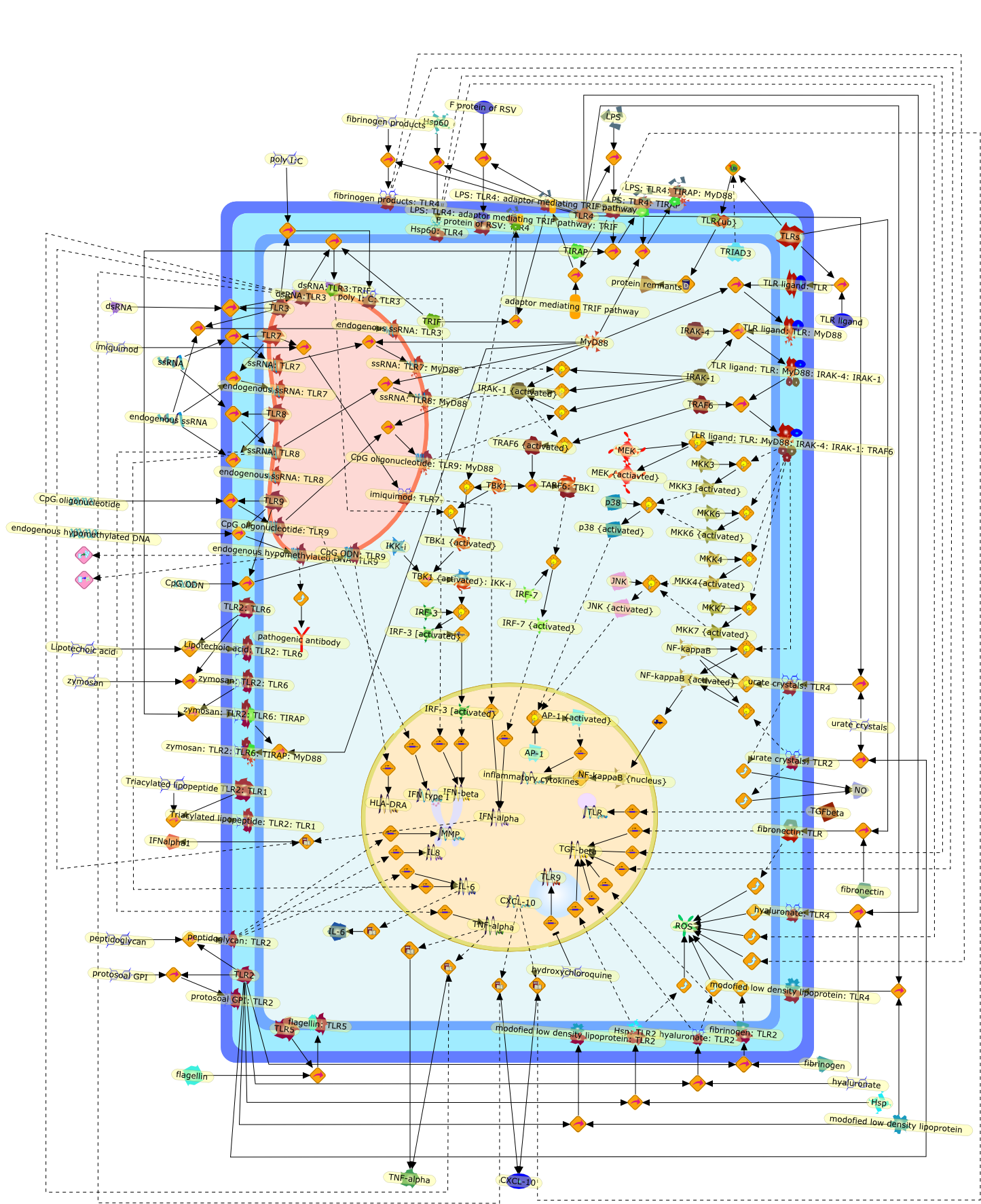{kind=link}
{kind=link}
AP-1,AP-1 {activated},CXCL-10,CpG ODN,CpG ODN: TLR9,CpG oligonucleotide,CpG oligonucleotide: TLR9,CpG oligonucleotide: TLR9: MyD88,F protein of RSV,F protein of RSV: TLR4,HLA-DRA,Hsp,Hsp60,Hsp60: TLR4,Hsp: TLR2,IFN type I,IFN-alpha,IFN-beta,IFNalpha1,IKK-i,IL-6,IL8,IRAK-1,IRAK-1 {activated},IRAK-4,IRF-3,IRF-3 [activated},IRF-7,IRF-7 {activated},JNK,JNK {activated},LPS,LPS: TLR4,LPS: TLR4: TIRAP,LPS: TLR4: TIRAP: MyD88,LPS: TLR4: adaptor mediating TRIF pathway,LPS: TLR4: adaptor mediating TRIF pathway: TRIF,Lipotechoic acid,Lipotechoic acid: TLR2: TLR6,MEK,MEK {actiavted},MKK3,MKK3 [activated},MKK4,MKK4{activated},MKK6,MKK6 {activated},MKK7,MKK7 {activated},MMP,MyD88,NF-kappaB,NF-kappaB {activated},NF-kappaB {nucleus},NO,ROS,TARF6: TBK1,TBK1,TBK1 {activated},TBK1 {activated}: IKK-i,TGF-beta,TGFbeta,TIRAP,TLR,TLR ligand,TLR ligand: TLR,TLR ligand: TLR: MyD88,TLR ligand: TLR: MyD88: IRAK-4: IRAK-1,TLR ligand: TLR: MyD88: IRAK-4: IRAK-1: TRAF6,TLR2,TLR2: TLR1,TLR2: TLR6,TLR3,TLR4,TLR5,TLR7,TLR8,TLR9,TLRs,TLR{ub},TNF-alpha,TRAF6,TRAF6 {activated},TRIAD3,TRIF,Triacylated lipopeptide,Triacylated lipopeptide: TLR2: TLR1,adaptor mediating TRIF pathway,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,endogenous hypomethylated DNA,endogenous hypomethylated DNA: TLR9,endogenous ssRNA,endogenous ssRNA: TLR3,endogenous ssRNA: TLR7,endogenous ssRNA: TLR8,fibrinogen,fibrinogen products,fibrinogen products: TLR4,fibrinogen: TLR2,fibronectin,fibronectin: TLR,flagellin,flagellin: TLR5,hyaluronate,hyaluronate: TLR2,hyaluronate: TLR4,hydroxychloroquine,imiquimod,imiquimod: TLR7,inflammatory cytokines,modofied low density lipoprotein,modofied low density lipoprotein: TLR2,modofied low density lipoprotein: TLR4,p38,p38 {activated},pathogenic antibody,peptidoglycan,peptidoglycan: TLR2,poly I: C: TLR3,poly I:C,protein remnants,protosoal GPI,protosoal GPI: TLR2,ssRNA,ssRNA: TLR7,ssRNA: TLR7: MyD88,ssRNA: TLR8,ssRNA: TLR8: MyD88,urate crystals,urate crystals: TLR2,urate crystals: TLR4,zymosan,zymosan: TLR2: TLR6,zymosan: TLR2: TLR6: TIRAP,zymosan: TLR2: TLR6: TIRAP: MyD88
The innate immune system comprises several classes of pattern recognitionreceptors, including Toll-like receptors (TLRs), NOD-like receptors (NLRs), andRIG-1-like receptors (RLRs). TLRs recognize microbes on the cell surface and inendosomes, whereas NLRs and RLRs detect microbial components in the cytosol.Here we discuss the recent understanding in NLRs. Two NLRs, NOD1 and NOD2, sensethe cytosolic presence of the peptidoglycan fragments meso-DAP and muramyldipeptide, respectively, and drive the activation of mitogen-activated proteinkinase (MAPK) and the transcription factor NF-kappaB. A different set of NLRsinduces caspase-1 activation through the assembly of large protein complexesnamed inflammasomes. Genetic variations in several NLR members are associatedwith the development of inflammatory disorders. Further understanding of NLRsshould provide new insights into the mechanisms of host defense and thepathogenesis of inflammatory diseases.
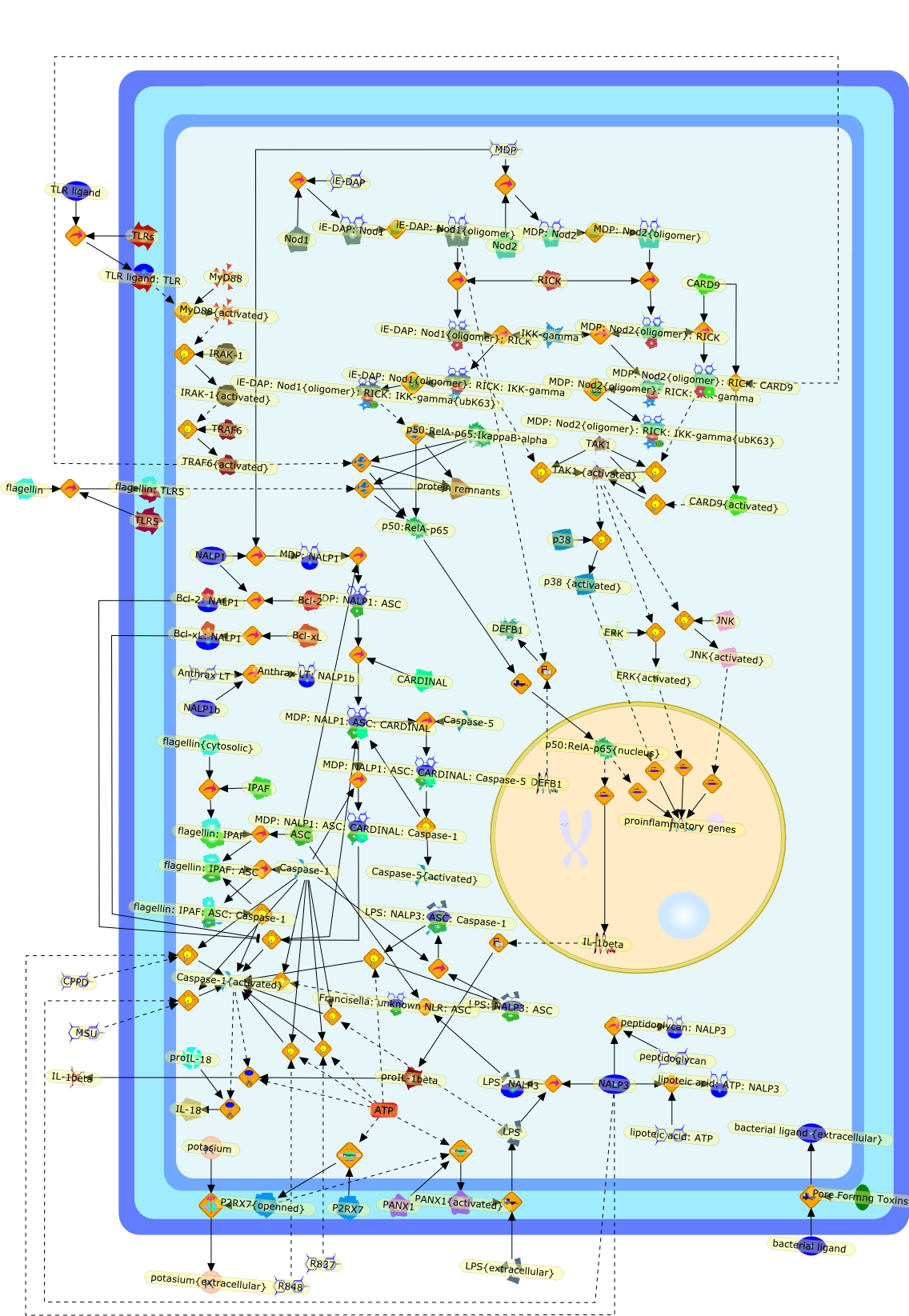{kind=link}
{kind=link}
ASC,ATP,Anthrax LT,Anthrax LT: NALP1b,Bcl-2,Bcl-2: NALP1,Bcl-xL,Bcl-xL: NALP1,CARD9,CARD9{activated},CARDINAL,CPPD,Caspase-1,Caspase-1{activated},Caspase-5,Caspase-5{activated},DEFB1,ERK,ERK{activated},Francisella: unknown NLR: ASC,IKK-gamma,IL-18,IL-1beta,IPAF,IRAK-1,IRAK-1{activated},JNK,JNK{activated},LPS,LPS: NALP3,LPS: NALP3: ASC,LPS: NALP3: ASC: Caspase-1,LPS{extracellular},MDP,MDP: NALP1,MDP: NALP1: ASC,MDP: NALP1: ASC: CARDINAL,MDP: NALP1: ASC: CARDINAL: Caspase-1,MDP: NALP1: ASC: CARDINAL: Caspase-5,MDP: Nod2,MDP: Nod2{oligomer},MDP: Nod2{oligomer}: RICK,MDP: Nod2{oligomer}: RICK: CARD9,MDP: Nod2{oligomer}: RICK: IKK-gamma,MDP: Nod2{oligomer}: RICK: IKK-gamma{ubK63},MSU,MyD88,MyD88{activated},NALP1,NALP1b,NALP3,Nod1,Nod2,P2RX7,P2RX7{openned},PANX1,PANX1{activated},Pore Formng Toxins,R837,R848,RICK,TAK1,TAK1 {activated},TLR ligand,TLR ligand: TLR,TLR5,TLRs,TRAF6,TRAF6{activated},bacterial ligand,bacterial ligand {extracellular},flagellin,flagellin: IPAF,flagellin: IPAF: ASC,flagellin: IPAF: ASC: Caspase-1,flagellin: TLR5,flagellin{cytosolic},iE-DAP,iE-DAP: Nod1,iE-DAP: Nod1{oligomer},iE-DAP: Nod1{oligomer}: RICK,iE-DAP: Nod1{oligomer}: RICK: IKK-gamma,iE-DAP: Nod1{oligomer}: RICK: IKK-gamma{ubK63},lipoteic acid: ATP,lipoteic acid: ATP: NALP3,p38,p38 {activated},p50:RelA-p65,p50:RelA-p65:IkappaB-alpha,p50:RelA-p65{nucleus},peptidoglycan,peptidoglycan: NALP3,potasium,potasium{extracellular},proIL-18,proIL-1beta,proinflammatory genes,protein remnants
Tuning is a key aspect of inflammatory reaction essential in homeostasis andpathology. An emerging mechanism for negative regulation of proinflammatorycytokines is based on non-signaling IL-1/TLR receptors and chemokine receptorscompeting with signaling receptors for ligand binding and sustaining ligandinternalization and degradation. Biological activities of IL-1R/TLR receptorsare under control of membrane-bound binding molecules lacking the signalingdomain, soluble receptor antagonists, and intracellular signaling inhibitors.The chemokine system includes at least three 'silent' receptors with distinctspecificity and tissue distribution. D6 is the best characterized representativemember of this class of negative regulators, binds most inflammatory, but nothomeostatic, CC chemokines and shuttles in a ligand-independent way from theplasma membrane to endocytic compartments where chemokines are targeted todegradation. In vitro and in vivo evidence, including results with gene targetedmice, is consistent with the view that these non-signaling receptors forproinflammatory cytokines possess unique functional and structural featureswhich make them ideally adapted to act as a decoy and scavenger receptors, witha non redundant role in dampening tissue inflammation and tuning draining lymphnodes reactivity.
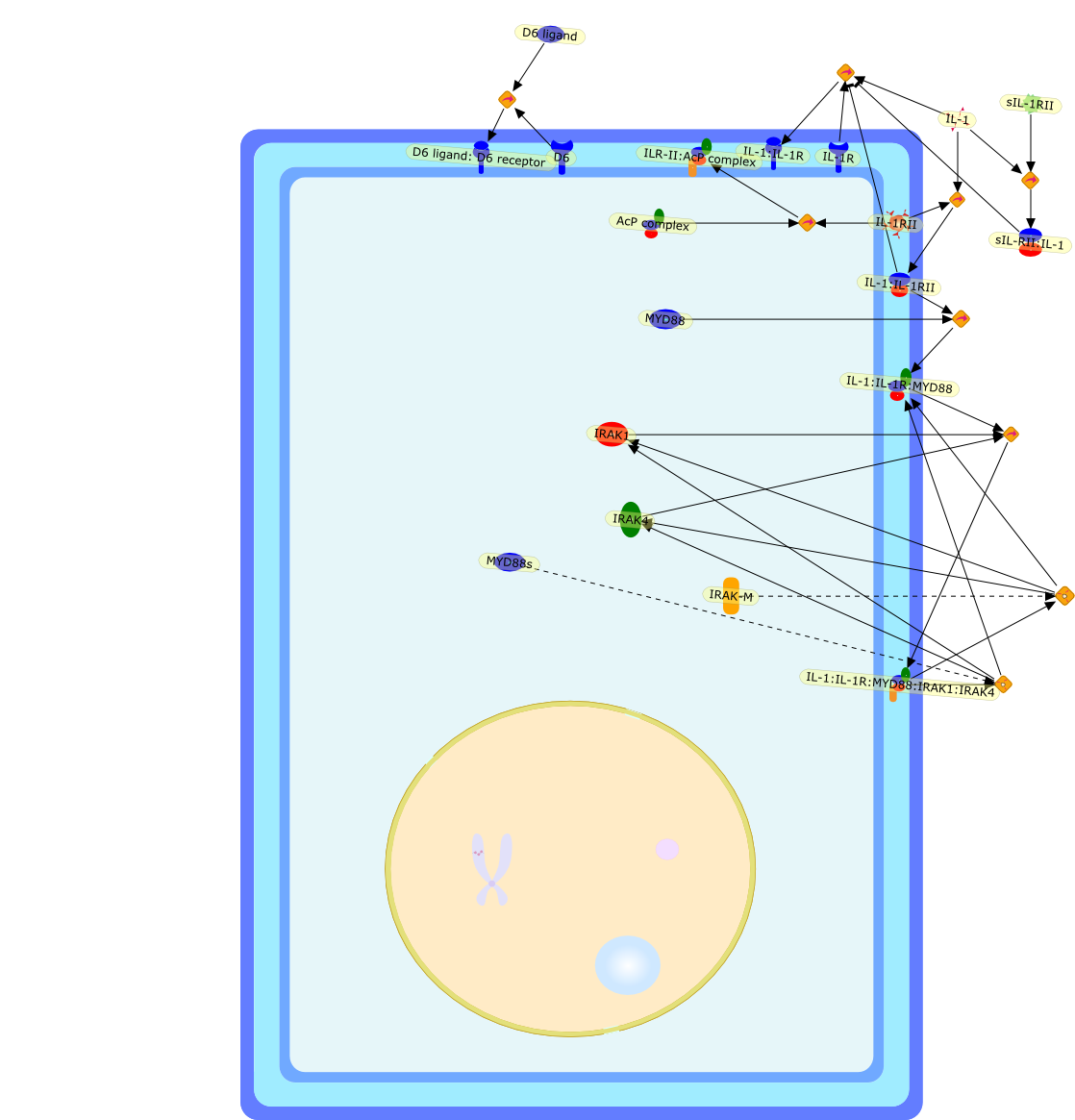{kind=link}
{kind=link}
AcP complex,D6,D6 ligand,D6 ligand: D6 receptor,IL-1,IL-1:IL-1R,IL-1:IL-1R:MYD88,IL-1:IL-1R:MYD88:IRAK1:IRAK4,IL-1:IL-1RII,IL-1R,IL-1RII,ILR-II:AcP complex,IRAK-M,IRAK1,IRAK4,MYD88,MYD88s,sIL-1RII,sIL-RII:IL-1
Toll-like receptors (TLRs) are a family of evolutionarily conserved moleculesthat directly detect pathogen invasion or tissue damage and initiate abiological response. TLRs can signal through two primary intracellular pathwaysand as such can induce either immuno-stimulatory or immuno-modulatory molecules.Both sides of this twin-edged sword are being examined for their therapeuticpotential in combating neurological disease. The immuno-stimulatory propertiesof TLRs are being used to generate tumor-specific immune responses to CNS tumorswhile the immuno-modulatory properties are being used to suppress damaginginflammatory responses to stroke. Recently, a third component of TLR signalinghas begun to emerge--that of direct neuroprotection. Hence, the TLRs offer noveltargets for the treatment of neurological disease.
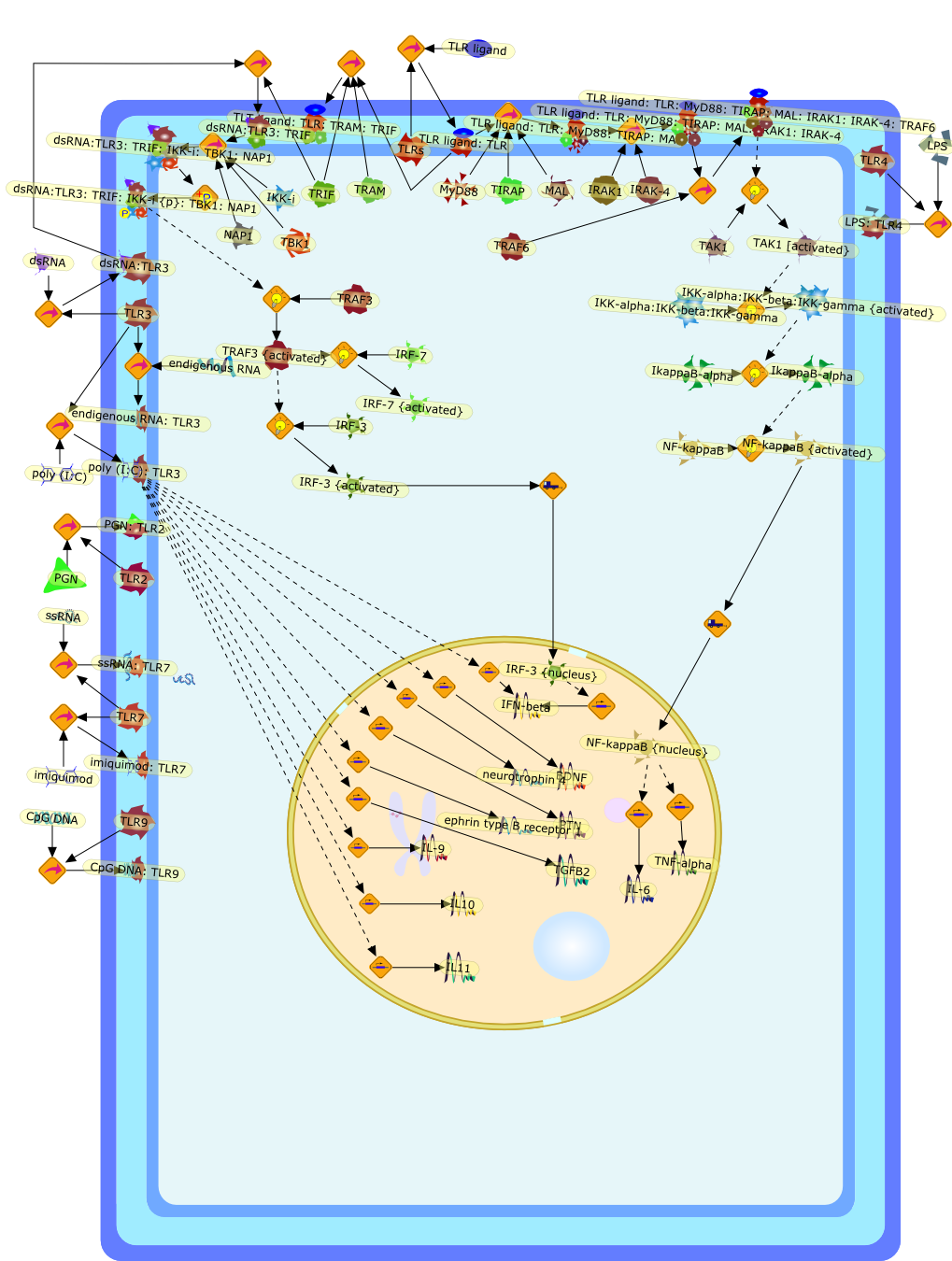{kind=link}
{kind=link}
BDNF,CpG DNA,CpG DNA: TLR9,IFN-beta,IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma {activated},IKK-i,IL-6,IL-9,IL10,IL11,IRAK-4,IRAK1,IRF-3,IRF-3 {activated},IRF-3 {nucleus},IRF-7,IRF-7 {activated},IkappaB-alpha,LPS,LPS: TLR4,MAL,MyD88,NAP1,NF-kappaB,NF-kappaB {activated},NF-kappaB {nucleus},PGN,PGN: TLR2,PTN,TAK1,TAK1 [activated},TBK1,TGFB2,TIRAP,TLR ligand,TLR ligand: TLR,TLR ligand: TLR: MyD88: TIRAP: MAL,TLR ligand: TLR: MyD88: TIRAP: MAL: IRAK1: IRAK-4,TLR ligand: TLR: MyD88: TIRAP: MAL: IRAK1: IRAK-4: TRAF6,TLR2,TLR3,TLR4,TLR7,TLR9,TLRs,TLT ligand: TLR: TRAM: TRIF,TNF-alpha,TRAF3,TRAF3 {activated},TRAF6,TRAM,TRIF,dsRNA,dsRNA:TLR3,dsRNA:TLR3: TRIF,dsRNA:TLR3: TRIF: IKK-i {p}: TBK1: NAP1,dsRNA:TLR3: TRIF: IKK-i: TBK1: NAP1,endigenous RNA,endigenous RNA: TLR3,ephrin type B receptor 1,imiquimod,imiquimod: TLR7,neurotrophin 4,poly (I:C),poly (I:C): TLR3,ssRNA,ssRNA: TLR7
The peroxisome proliferator-activated receptors (PPARalpha, -gamma, and-beta/delta) are nuclear receptors with distinct patterns of expression in manycell types both within and outside the immune system. PPAR ligands haveanti-inflammatory activity in a variety of mouse models for acute and chronicinflammation. In macrophages, PPARgamma ligands repress expression of a subsetof Toll-like receptor (TLR) target genes by a molecular mechanism termedligand-dependent transrepression. In chronic inflammation, ligand-boundPPARalpha represses production of IFNgamma and IL-17 by CD4(+) T cells, andPPARgamma ligands modulate dendritic cell function to elicit the development ofanergic CD4(+) T cells. PPAR ligands also repress expression of cell adhesionmolecules on endothelial cells and the secretion of chemokines by epithelial andother cells, decreasing the recruitment of leukocytes to the site ofinflammation. The anti-inflammatory activity of PPAR ligands in mouse modelssuggests their possible use for treating human inflammatory and autoimmunediseases.
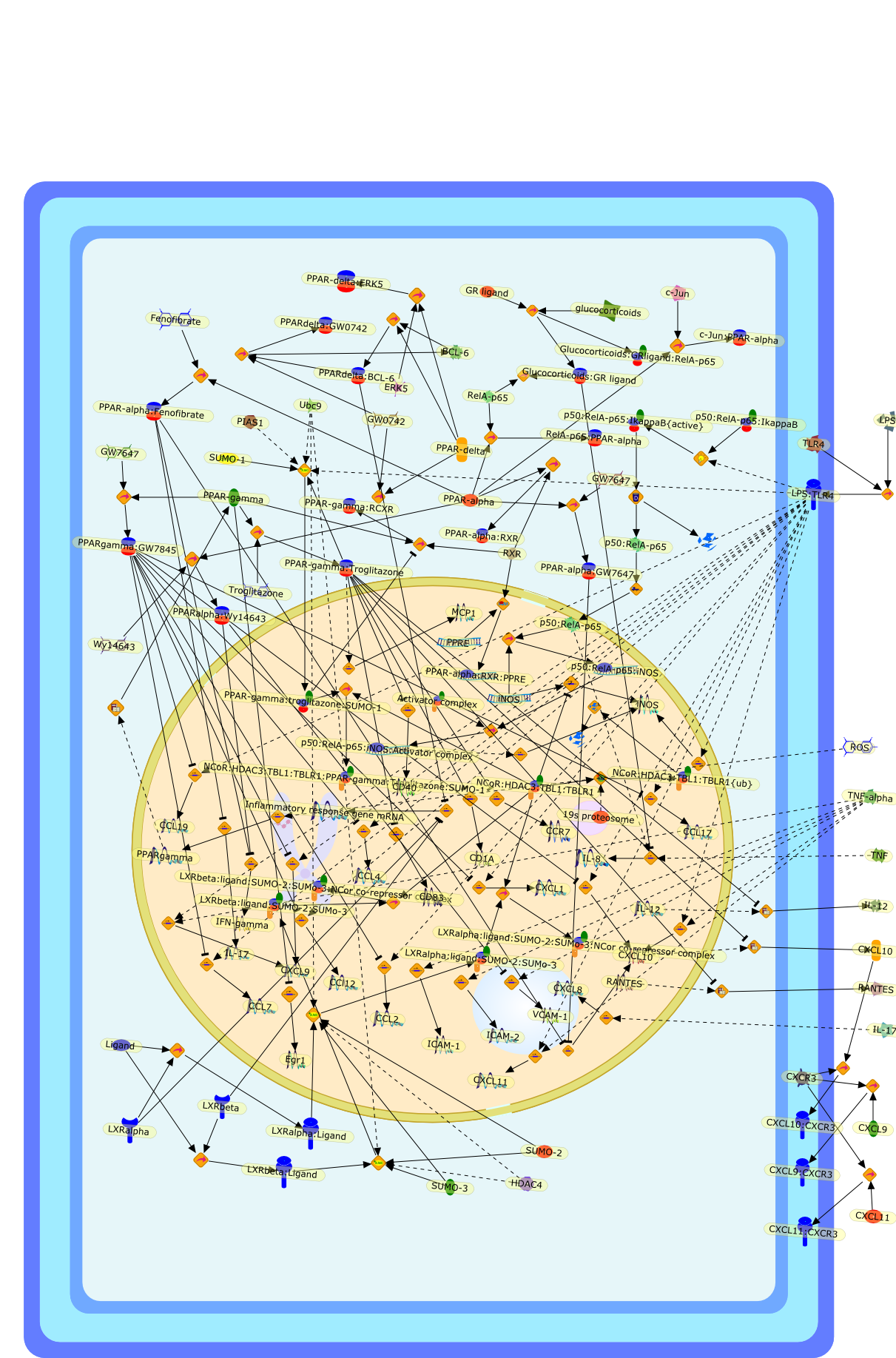{kind=link}
{kind=link}
19s proteosome,Activator complex,BCL-6,CCL17,CCL19,CCL2,CCL4,CCL7,CCR7,CCl12,CD1A,CD40,CD83,CXCL1,CXCL10,CXCL10:CXCR3,CXCL11,CXCL11:CXCR3,CXCL8,CXCL9,CXCL9:CXCR3,CXCR3,Degradants,ERK5,Egr1,Fenofibrate,GR ligand,GW0742,GW7647,Glucocorticoids:GR ligand,Glucocorticoids:GRligand:RelA-p65,HDAC4,ICAM-1,ICAM-2,IFN-gamma,IL-12,IL-17,IL-8,Inflammatory response gene mRNA,LPS,LPS:TLR4,LXRalpha,LXRalpha:Ligand,LXRalpha:ligand:SUMO-2:SUMo-3,LXRalpha:ligand:SUMO-2:SUMo-3:NCor co-repressor complex,LXRbeta,LXRbeta:Ligand,LXRbeta:ligand:SUMO-2:SUMo-3,LXRbeta:ligand:SUMO-2:SUMo-3:NCor co-repressor complex,Ligand,MCP1,NCoR:HDAC3:TBL1:TBLR1,NCoR:HDAC3:TBL1:TBLR1:PPAR-gamma:Troglitazone:SUMO-1,NCoR:HDAC3:TBL1:TBLR1{ub},PIAS1,PPAR-alpha,PPAR-alpha:Fenofibrate,PPAR-alpha:GW7647,PPAR-alpha:RXR,PPAR-alpha:RXR:PPRE,PPAR-delta,PPAR-delta:ERK5,PPAR-gamma,PPAR-gamma:RCXR,PPAR-gamma:Troglitazone,PPAR-gamma:troglitazone:SUMO-1,PPARalpha:Wy14643,PPARdelta:BCL-6,PPARdelta:GW0742,PPARgamma,PPARgamma:GW7845,PPRE,RANTES,ROS,RXR,RelA-p65,RelA-p65:PPAR-alpha,SUMO-1,SUMO-2,SUMO-3,TLR4,TNF,TNF-alpha,Troglitazone,Ubc9,VCAM-1,Wy14643,c-Jun,c-Jun:PPAR-alpha,glucocorticoids,iNOS,p50:RelA-p65,p50:RelA-p65:IkappaB,p50:RelA-p65:IkappaB{active},p50:RelA-p65:iNOS,p50:RelA-p65:iNOS:Activator complex,proteosome degradants
Subclinical inflammation is a candidate etiological factor in the pathogenesisof metabolic syndrome and in the progression of atherosclerosis. A central rolefor activated macrophages has been elucidated recently as important regulatorsof the inflammatory process in atherosclerosis. Macrophage differentiation andfunction can be modulated by a class of transcription factors termed nuclearreceptors. These are activated by intermediary products of basic metabolicprocesses. In this review the contribution of peroxisome proliferator-activatedreceptors and liver X receptors to macrophage functions in inflammation andlipid metabolism will be discussed in light of their roles in macrophages duringatherosclerosis. In the past decade much effort has been made to understand themechanisms how lipids are handled by macrophages and how inflammation couldpromote the atherogenic process. Here, we also provide an overview of these twofields.
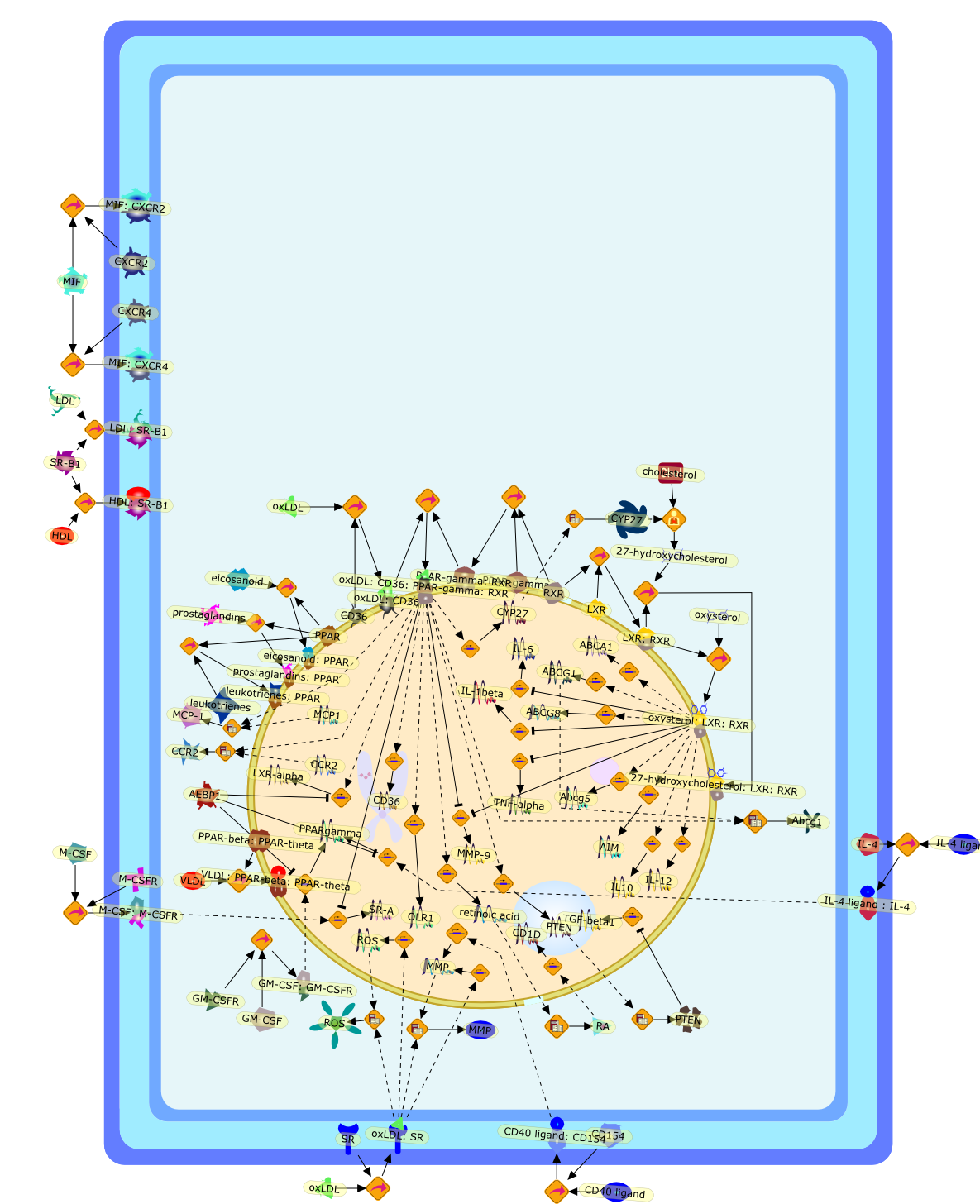{kind=link}
{kind=link}
27-hydroxycholesterol,27-hydroxycholesterol: LXR: RXR,ABCA1,ABCG1,ABCG8,AEBP1,AIM,Abcg1,Abcg5,CCR2,CD154,CD1D,CD36,CD40 ligand,CD40 ligand: CD154,CXCR2,CXCR4,CYP27,GM-CSF,GM-CSF: GM-CSFR,GM-CSFR,HDL,HDL: SR-B1,IL-12,IL-1beta,IL-4,IL-4 ligand,IL-4 ligand : IL-4,IL-6,IL10,LDL,LDL: SR-B1,LXR,LXR-alpha,LXR: RXR,M-CSF,M-CSF: M-CSFR,M-CSFR,MCP-1,MCP1,MIF,MIF: CXCR2,MIF: CXCR4,MMP,MMP-9,OLR1,PPAR,PPAR-beta: PPAR-theta,PPAR-gamma,PPAR-gamma: RXR,PPARgamma,PTEN,RA,ROS,RXR,SR,SR-A,SR-B1,TGF-beta1,TNF-alpha,VLDL,VLDL: PPAR-beta: PPAR-theta,cholesterol,eicosanoid,eicosanoid: PPAR,leukotrienes,leukotrienes: PPAR,oxLDL,oxLDL: CD36,oxLDL: CD36: PPAR-gamma: RXR,oxLDL: SR,oxysterol,oxysterol: LXR: RXR,prostaglandins,prostaglandins: PPAR,retinoic acid
Innate immunity is the first line of defense against invading pathogens. Afamily of Toll-like receptors (TLRs) acts as primary sensors that detect a widevariety of microbial components and elicit innate immune responses. All TLRsignaling pathways culminate in activation of the transcription factor nuclearfactor-kappaB (NF-kappaB), which controls the expression of an array ofinflammatory cytokine genes. NF-kappaB activation requires the phosphorylationand degradation of inhibitory kappaB (IkappaB) proteins, which is triggered bytwo kinases, IkappaB kinase alpha (IKKalpha) and IKKbeta. In addition, severalTLRs activate alternative pathways involving the IKK-related kinases TBK1 [TRAFfamily member-associated NF-kappaB activator (TANK) binding kinase-1] and IKKi,which elicit antiviral innate immune responses. Here, we review recent progressin our understanding of the role of NF-kappaB in TLR signaling pathways anddiscuss potential implications for molecular medicine.
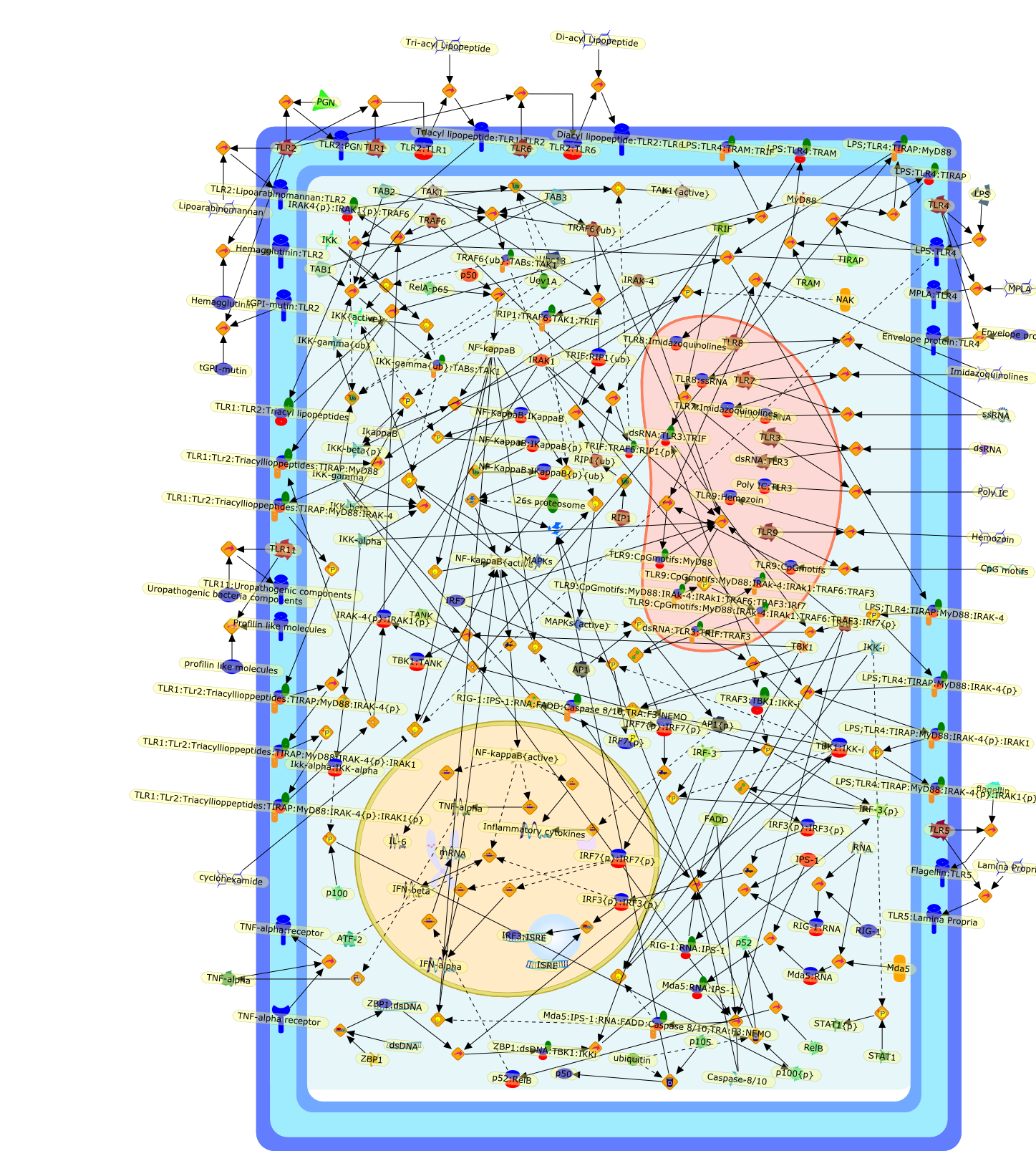{kind=link}
{kind=link}
26s proteosome,AP1,AP1{p},ATF-2,Caspase-8/10,CpG motifs,Di-acyl Lipopeptide,Diacyl lipopeptide:TLR2:TLR6,Envelope protein,Envelope protein:TLR4,FADD,Flagellin:TLR5,Hemagglutinin,Hemagglutinin:TLR2,Hemozoin,IFN-alpha,IFN-beta,IKK,IKK-alpha,IKK-beta,IKK-beta{p},IKK-gamma,IKK-gamma{ub},IKK-gamma{ub}:TABs:TAK1,IKK-i,IKK{active},IL-6,IPS-1,IRAK-4,IRAK-4{p}:IRAK1{p},IRAK1,IRAK4{p}:IRAK1{p}:TRAF6,IRF-3,IRF-3{p},IRF3:ISRE,IRF3{p}:IRF3{p},IRF7,IRF7{p},IRF7{p}:IRF7{p},ISRE,IkappaB,Ikk-alpha:IKK-alpha,Imidazoquinolines,Inflammatory cytokines,LPS,LPS:TLR4,LPS:TLR4:TIRAP,LPS:TLR4:TRAM,LPS:TLR4:TRAM:TRIF,LPS;TLR4:TIRAP:MyD88,LPS;TLR4:TIRAP:MyD88:IRAK-4,LPS;TLR4:TIRAP:MyD88:IRAK-4{p},LPS;TLR4:TIRAP:MyD88:IRAK-4{p}:IRAK1,LPS;TLR4:TIRAP:MyD88:IRAK-4{p}:IRAK1{p},Lamina Propria,Lipoarabinomannan,MAPKs,MAPKs{active},MPLA,MPLA:TLR4,Mda5,Mda5:IPS-1:RNA:FADD:Caspase 8/10,TRA:F3:NEMO,Mda5:RNA,Mda5:RNA:IPS-1,MyD88,NAK,NF-KappaB:IKappaB,NF-KappaB:IKappaB{p},NF-KappaB:IKappaB{p}{ub},NF-kappaB,NF-kappaB{active},PGN,Poly IC,Poly IC:TLR3,Profilin like molecules,RIG-1,RIG-1:IPS-1:RNA:FADD:Caspase 8/10,TRA:F3:NEMO,RIG-1:RNA,RIG-1:RNA:IPS-1,RIP1,RIP1:TRAF6:TAK1:TRIF,RIP1{ub},RNA,RelA-p65,RelB,STAT1,STAT1{p},TAB1,TAB2,TAB3,TAK1,TAK1{active},TANK,TBK1,TBK1:IKK-i,TBK1:TANK,TIRAP,TLR1,TLR11,TLR11:Uropathogenic components,TLR1:TLR2:Triacyl lipopeptides,TLR1:TLr2:Triacyllioppeptides:TIRAP:MyD88,TLR1:TLr2:Triacyllioppeptides:TIRAP:MyD88:IRAK-4,TLR1:TLr2:Triacyllioppeptides:TIRAP:MyD88:IRAK-4{p},TLR1:TLr2:Triacyllioppeptides:TIRAP:MyD88:IRAK-4{p}:IRAK1,TLR1:TLr2:Triacyllioppeptides:TIRAP:MyD88:IRAK-4{p}:IRAK1{p},TLR2,TLR2:Lipoarabinomannan:TLR2,TLR2:PGN,TLR2:TLR1,TLR2:TLR6,TLR3,TLR4,TLR5,TLR5:Lamina Propria,TLR6,TLR7,TLR7:Imidazoquinolines,TLR7:ssRNA,TLR8,TLR8:Imidazoquinolines,TLR8:ssRNA,TLR9,TLR9:CpGmotifs,TLR9:CpGmotifs:MyD88,TLR9:CpGmotifs:MyD88:IRAk-4:IRAk1:TRAF6:TRAF3,TLR9:CpGmotifs:MyD88:IRAk-4:IRAk1:TRAF6:TRAF3:IRf7,TLR9:CpGmotifs:MyD88:IRAk-4:IRAk1:TRAF6:TRAF3:IRf7{p},TLR9:Hemozoin,TNF-alpha,TNF-alpha receptor,TNF-alpha:receptor,TRAF3,TRAF3:TBK1:IKK-i,TRAF6,TRAF6{ub},TRAF6{ub}:TABs:TAK1,TRAM,TRIF,TRIF:RIP1{ub},TRIF:TRAF6:RIP1{p},Tri-acyl Lipopeptide,Triacyl lipopeptide:TLR1:TLR2,Ubc13,Uev1A,Uropathogenic bacteria components,ZBP1,ZBP1:dsDNA,ZBP1:dsDNA:TBK1:IKKi,cyclohexamide,dsDNA,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:TRAF3,flagellin,mRNA,p100,p100{p},p105,p50,p52,p52:RelB,profilin like molecules,proteosome remnant,ssRNA,tGPI-mutin,tGPI-mutin:TLR2,ubiquitin
At one time, atherosclerosis was thought to be a simple lipid storage disease.However, it is now recognized as a chronic and progressive inflammation of thearterial wall. Gene deletion experiments in murine models of atherosclerosisthat reduce the inflammatory process also reduce disease severity. Identifyingthe initiators and mediators of that inflammation can provide promising avenuesfor prevention or therapy. Two prominent risk factors, hyperlipidaemia andinfectious disease, point to innate immune mechanisms as potential contributorsto proatherogenic inflammation. The TLRs (Toll-like receptors), pro-inflammatorysensors of pathogens, are potential links between inflammation, infectiousdisease and atherosclerosis. A mechanism for hyperlipidaemic initiation ofsterile inflammation can be postulated because oxidized lipoproteins or theircomponent oxidized lipids have been identified as TLR ligands. Moreover,infectious agents are correlated with atherosclerosis risk. We have identified arole for TLR2 in atherosclerosis in mice deficient in low-density lipoproteinreceptor. We observed that proatherogenic TLR2 responses to unknown endogenousor unknown endemic exogenous agonists are mediated by non-BMDC(bone-marrow-derived cells), which can include endothelial cells. In contrast,the proatherogenic TLR2 responses to the defined synthetic exogenous agonistPam3 CSK4 are mediated at least in part by BMDC, which can include lymphocytes,monocytes/macrophages and dendritic cells. TLR2-mediated cell activation inresponse to endogenous and exogenous agents is proatherogenic in hyperlipidaemicmice.
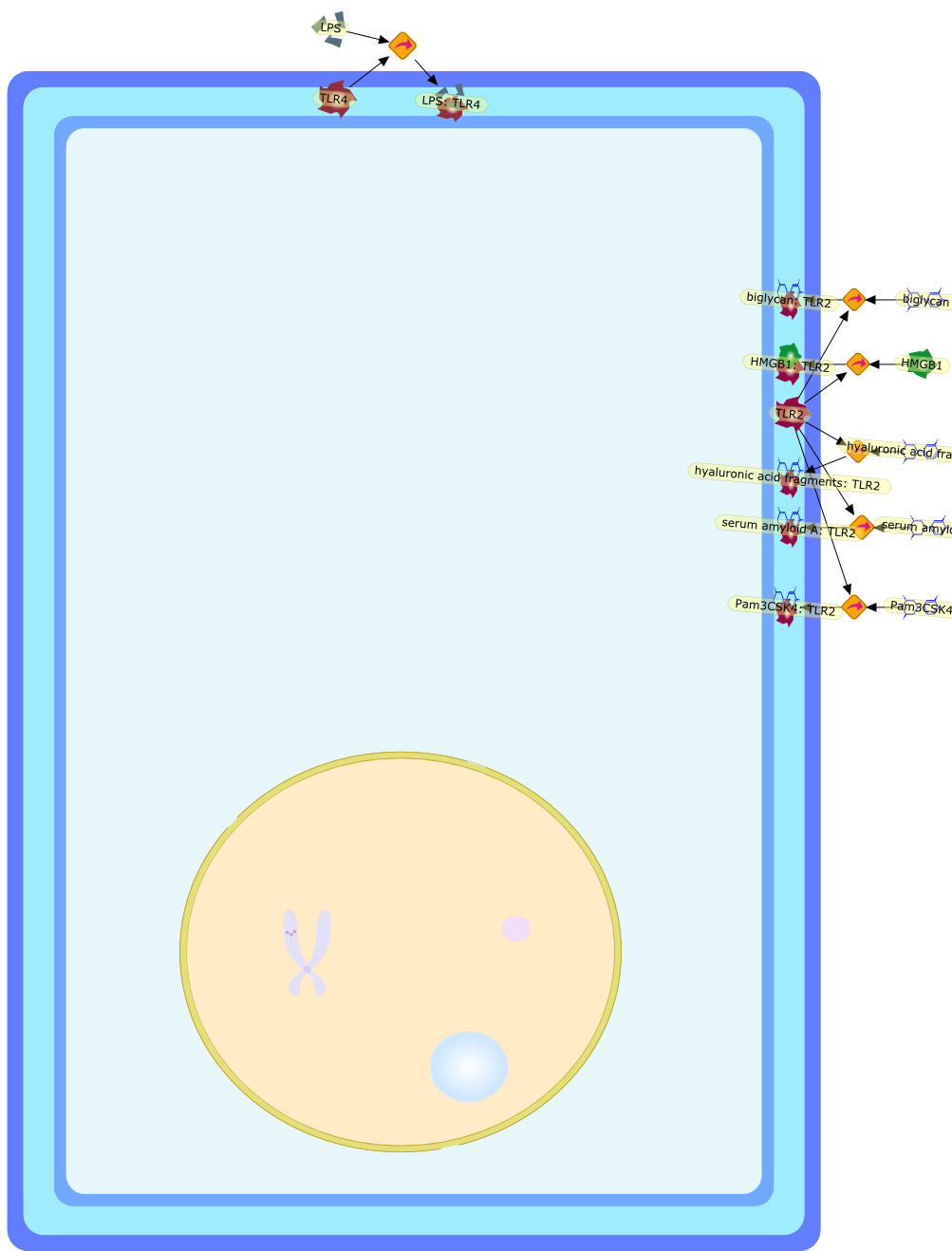{kind=link}
{kind=link}
HMGB1,HMGB1: TLR2,LPS,LPS: TLR4,Pam3CSK4,Pam3CSK4: TLR2,TLR2,TLR4,biglycan,biglycan: TLR2,hyaluronic acid fragments,hyaluronic acid fragments: TLR2,serum amyloid A,serum amyloid A: TLR2
Nod (nucleotide-binding oligomerization domain) 1 and Nod2 are intracellularPRMs (pattern-recognition molecules) of the NLR (Nod-like receptor) family.These proteins are implicated in the detection of bacterial peptidoglycan andregulate pro-inflammatory pathways in response to bacteria by inducingsignalling pathways such as NF-kappaB (nuclear factor kappaB) and MAPKs(mitogen-activated protein kinases). The Nod proteins act independently of theTLR (Toll-like receptor) cascade, but potently synergize with the latter totrigger innate immune responses to microbes. Most importantly, mutations in Nod2have been shown to confer susceptibility to several chronic inflammatorydisorders, including Crohn's disease, Blau syndrome and early-onset sarcoidosis,underscoring the role of Nod2 in inflammatory homoeostasis. This reviewsummarizes the most recent findings in the field of Nod1 and Nod2 research.
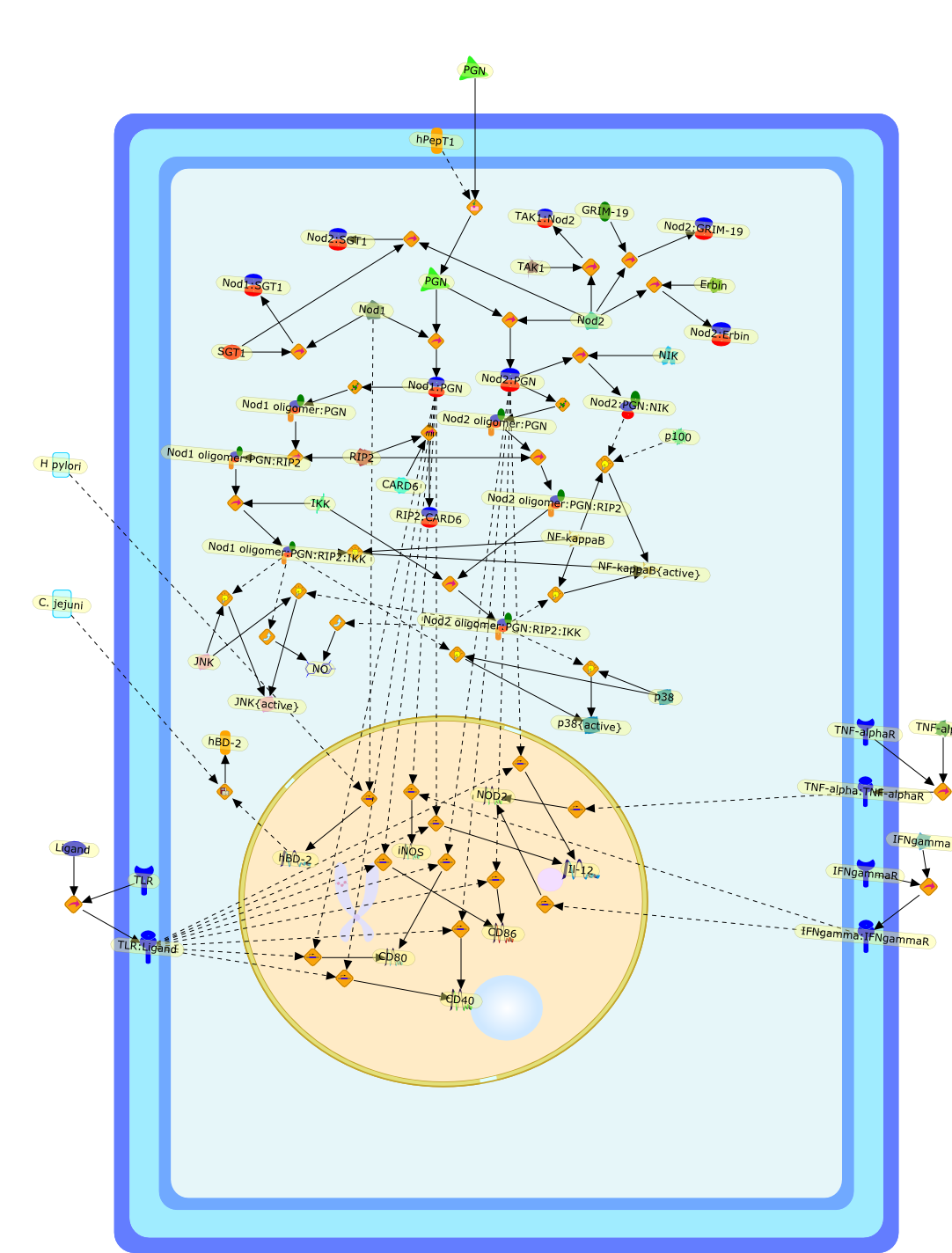{kind=link}
{kind=link}
C. jejuni,CARD6,CD40,CD80,CD86,Erbin,GRIM-19,H pylori,IFNgamma,IFNgamma:IFNgammaR,IFNgammaR,IKK,Il-12,JNK,JNK{active},Ligand,NF-kappaB,NF-kappaB{active},NIK,NO,NOD2,Nod1,Nod1 oligomer:PGN,Nod1 oligomer:PGN:RIP2,Nod1 oligomer:PGN:RIP2:IKK,Nod1:PGN,Nod1:SGT1,Nod2,Nod2 oligomer:PGN,Nod2 oligomer:PGN:RIP2,Nod2 oligomer:PGN:RIP2:IKK,Nod2:Erbin,Nod2:GRIM-19,Nod2:PGN,Nod2:PGN:NIK,Nod2:SGT1,PGN,RIP2,RIP2:CARD6,SGT1,TAK1,TAK1:Nod2,TLR,TLR:Ligand,TNF-alpha,TNF-alpha:TNF-alphaR,TNF-alphaR,hBD-2,hPepT1,iNOS,p100,p38,p38{active}
The neutrophil is a crucial early defence against microbial infection, butneutrophilic inflammation can result in devastating acute and chronicinflammatory diseases. In the lungs, the neutrophil is a principal part of thepathology of the acute respiratory distress syndrome, and its activation mayalso be of substantial importance in chronic obstructive pulmonary disease andsome forms of asthma. Induction of neutrophil recruitment in response tomicrobial attack requires activation of TLR (Toll-like receptor)-basedsignalling pathways and the concerted actions of multiple cell types, includingsentinel cells such as monocytes and macrophages acting together with tissuecell types such as the epithelium or smooth-muscle cell. The present reviewdescribes some of these networks and the resulting potential for their targetingin respiratory disease.
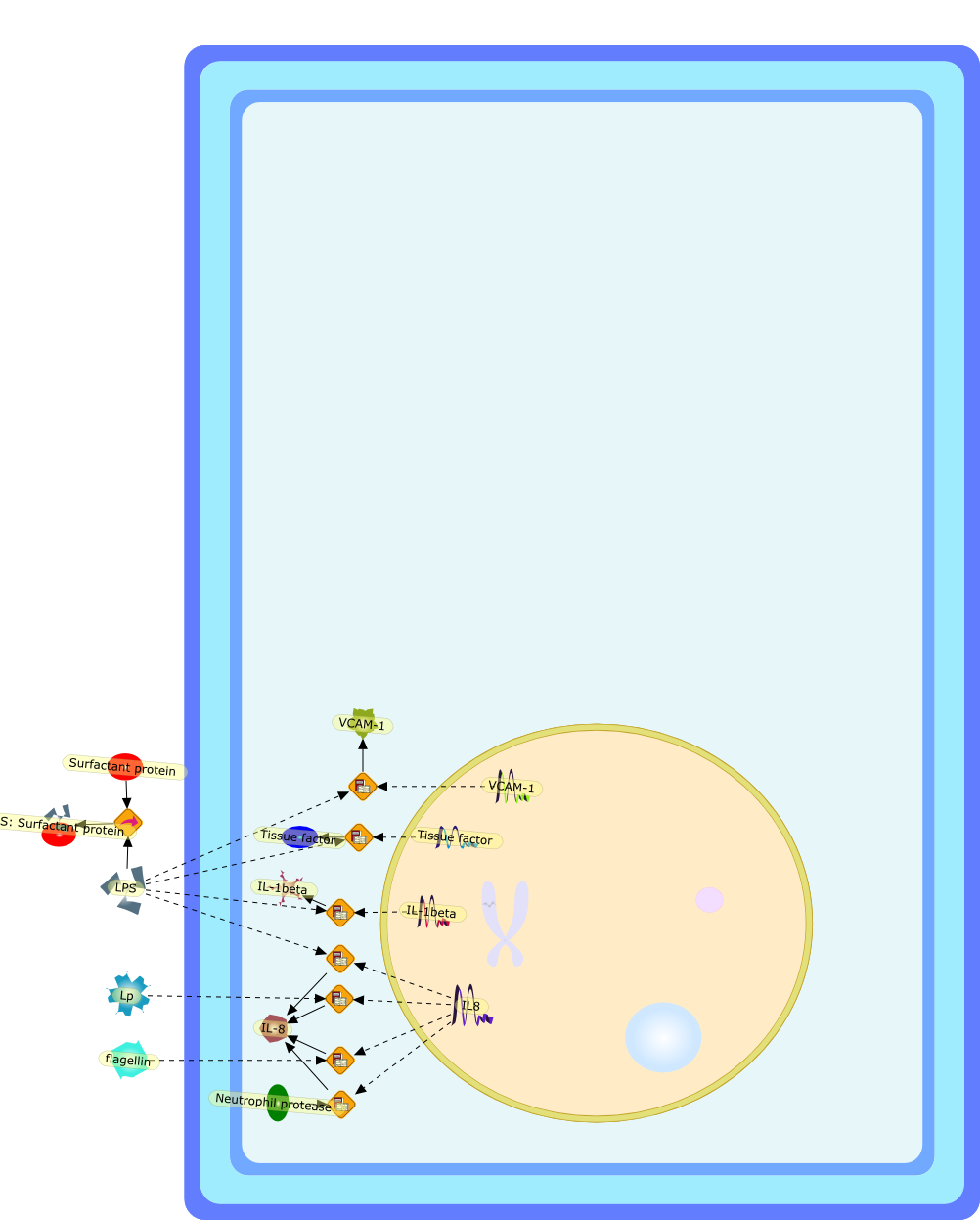{kind=link}
{kind=link}
IL-1beta,IL-8,IL8,LPS,LPS: Surfactant protein,Lp,Neutrophil protease,Surfactant protein,Tissue factor,VCAM-1,flagellin
Viral recognition is mediated by different classes of PRRs (pattern-recognitionreceptors) among which the TLRs (Toll-like receptors) and the RLHs [RIG(retinoic-acid-inducible)-like helicases] play major roles. The detection ofPAMPs (pathogen-associated molecular patterns) by these PRRs leads to theinitiation of signalling pathways that ultimately result in the activation oftranscription factors such as NF-kappaB (nuclear factor kappaB) and IRF-3 [IFN(interferon) regulatory factor-3] and IRF-7 and the induction ofpro-inflammatory cytokines and type I IFNs. Viruses have evolved a fine-tunedmechanism to evade detection by the immune system or to interfere with theresulting signalling pathways. Here, we discuss viral evasion proteins thatspecifically interfere with TLR and/or RLH signalling.
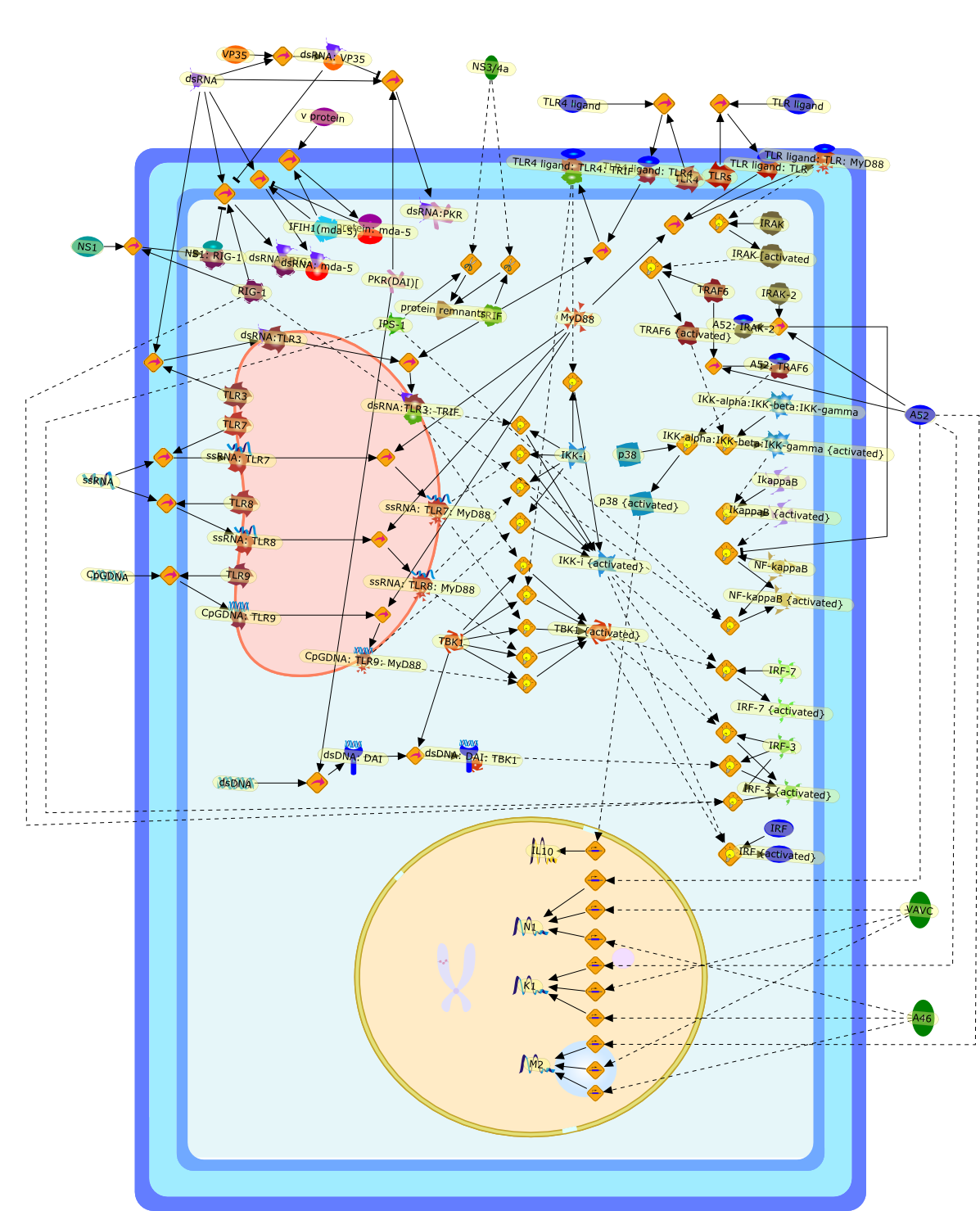{kind=link}
{kind=link}
A46,A52,A52: IRAK-2,A52: TRAF6,CpGDNA,CpGDNA: TLR9,CpGDNA: TLR9: MyD88,IFIH1(mda-5),IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma {activated},IKK-i,IKK-i {activated},IL10,IPS-1,IRAK,IRAK [activated,IRAK-2,IRF,IRF {activated},IRF-3,IRF-3 {activated},IRF-7,IRF-7 {activated},IkappaB,IkappaB {activated},K1,M2,MyD88,N1,NF-kappaB,NF-kappaB {activated},NS1,NS1: RIG-1,NS3/4a,PKR(DAI)[,RIG-1,TBK1,TBK1 {activated},TLR ligand,TLR ligand: TLR,TLR ligand: TLR: MyD88,TLR3,TLR4,TLR4 ligand,TLR4 ligand: TLR4,TLR4 ligand: TLR4: TRIF,TLR7,TLR8,TLR9,TLRs,TRAF6,TRAF6 {activated},TRIF,VAVC,VP35,dsDNA,dsDNA: DAI,dsDNA: DAI: TBK1,dsRNA,dsRNA: RIG-1,dsRNA: VP35,dsRNA: mda-5,dsRNA:PKR,dsRNA:TLR3,dsRNA:TLR3: TRIF,p38,p38 {activated},protein remnants,ssRNA,ssRNA: TLR7,ssRNA: TLR7: MyD88,ssRNA: TLR8,ssRNA: TLR8: MyD88,v protein,v protein: mda-5
The family of interferon (IFN) proteins has now more than reached the potentialenvisioned by early discovering virologists: IFNs are not only antivirals with aspectrum of clinical effectiveness against both RNA and DNA viruses, but arealso the prototypic biological response modifiers for oncology, and showeffectiveness in suppressing manifestations of multiple sclerosis. Studies ofIFNs have resulted in fundamental insights into cellular signalling mechanisms,gene transcription and innate and acquired immunity. Further elucidation of themultitude of IFN-induced genes, as well as drug development strategies targetingIFN production via the activation of the Toll-like receptors (TLRs), will almostcertainly lead to newer and more efficacious therapeutics. Our goal is to offera molecular and clinical perspective that will enable IFNs or their TLR agonistinducers to reach their full clinical potential.
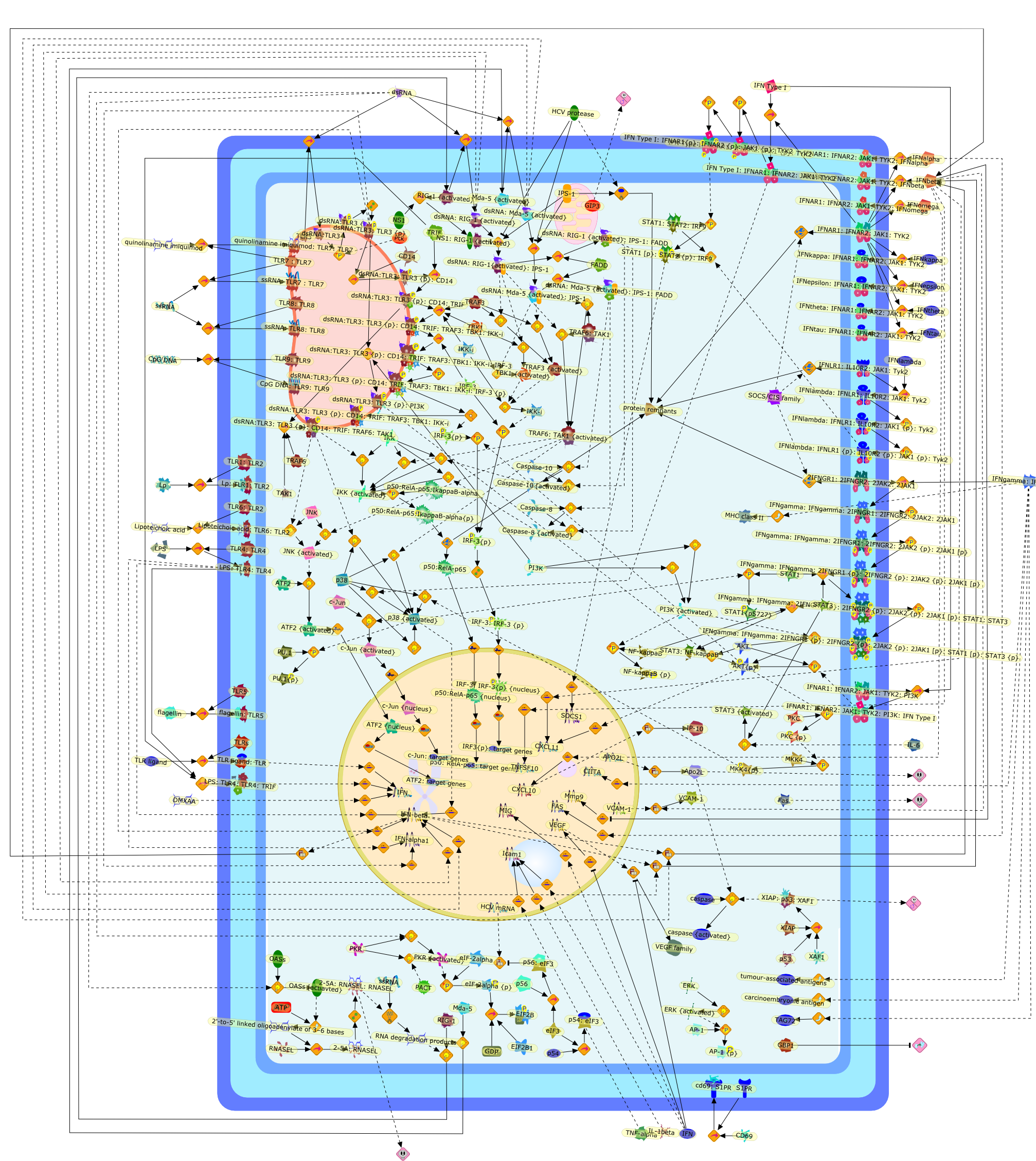{kind=link}
{kind=link}
2'-to-5' linked oligoadenylate of 3–6 bases,2-5A: RNASEL,2-5A: RNASEL: RNASEL,2IFNGR1: 2IFNGR2: 2JAK2: 2JAK1,: EIF2B,AKT,AKT{p},AP-1,AP-1 {p},APO2L,ATF2,ATF2 {activated},ATF2 {nucleus},ATF2: target genes,ATP,Apo2L,CD14,CD69,CIITA,CXCL10,CXCL11,Caspase-10,Caspase-10 [activated},Caspase-8,Caspase-8 {activated},CpG DNA,CpG DNA: TLR9: TLR9,DMXAA,EIF2B1,ERK,ERK {activated},FADD,FAS,Fas,GBP1,GDP,GIP3,HCV mRNA,HCV protease,IFN,IFN Type I,IFN Type I: IFNAR1: IFNAR2: JAK1 {p}: TYK2,IFN Type I: IFNAR1: IFNAR2: JAK1: TYK2,IFN Type I: IFNAR1{p}: IFNAR2 {p}: JAK1 {p}: TYK2,IFN-alpha1,IFN-beta,IFNAR1: IFNAR2: JAK1: TYK2,IFNAR1: IFNAR2: JAK1: TYK2: IFNalpha,IFNAR1: IFNAR2: JAK1: TYK2: IFNbeta,IFNAR1: IFNAR2: JAK1: TYK2: IFNomega,IFNAR1: IFNAR2: JAK1: TYK2: PI3K,IFNAR1: IFNAR2: JAK1: TYK2: PI3K: IFN Type I,IFNLR1: IL10R2: JAK1: Tyk2,IFNalpha,IFNbeta,IFNepsilon,IFNepsilon: IFNAR1: IFNAR2: JAK1: TYK2,IFNgamma: IFNgamma,IFNgamma: IFNgamma: 2IFNGR1 {p}: 2IFNGR2 {p}: 2JAK2 {p}: 2JAK1 [p},IFNgamma: IFNgamma: 2IFNGR1 {p}: 2IFNGR2 {p}: 2JAK2 {p}: 2JAK1 [p}: STAT1 [p}: STAT3 {p},IFNgamma: IFNgamma: 2IFNGR1 {p}: 2IFNGR2 {p}: 2JAK2 {p}: 2JAK1 [p}: STAT1: STAT3,IFNgamma: IFNgamma: 2IFNGR1: 2IFNGR2: 2JAK2 {p}: 2JAK1 [p},IFNgamma: IFNgamma: 2IFNGR1: 2IFNGR2: 2JAK2: 2JAK1,IFNkappa,IFNkappa: IFNAR1: IFNAR2: JAK1: TYK2,IFNlambda,IFNlambda: IFNLR1 {p}: IL10R2 {p}: JAK1 {p}: Tyk2,IFNlambda: IFNLR1: IL10R2: JAK1 {p}: Tyk2,IFNlambda: IFNLR1: IL10R2: JAK1: Tyk2,IFNomega,IFNtau,IFNtau: IFNAR1: IFNAR2: JAK1: TYK2,IFNtheta,IFNtheta: IFNAR1: IFNAR2: JAK1: TYK2,IKK,IKK {activated},IKK-i,IL-1beta,IL-6,IP-10,IPS-1,IRF-3,IRF-3: IRF-3 {p},IRF-3: IRF-3{p} {nucleus},IRF-3{p},IRF3{p}: target genes,Icam1,JNK,JNK {activated},LPS,LPS: TLR4: TLR4,LPS: TLR4: TLR4: TRIF,Lipoteichoic acid,Lipoteichoic acid: TLR6: TLR2,Lp,Lp: TLR1: TLR2,MHC class II,MIG,MKK4,MKK4{p},Mda-5,Mda-5 {activated},Mmp9,NF-kappaB,NF-kappaB {p},NS1,NS1: RIG-1 {activated},OASs,OASs [actiavted},PACT,PI3K,PI3K {activated},PKC,PKC {p},PKR,PKR {activated},PU.1,PU.1{p},Ptk,RIG-1,RIG-1 {activated},RNA degradation products,RNASEL,S1PR,SOCS/CIS family,SOCS1,STAT1,STAT1 [p}: STAT2 {p}: IRF9,STAT1: STAT2: IRF9,STAT1{pS727},STAT3,STAT3 {activated},STAT3: NF-kappaB,TAG72,TAK1,TBK1,TBK1 {activated},TLR ligand,TLR ligand: TLR,TLR1: TLR2,TLR3,TLR4: TLR4,TLR5,TLR6: TLR2,TLR7 : TLR7,TLR8: TLR8,TLR9: TLR9,TLRs,TNF-alpha,TNFSF10,TRAF3,TRAF3 {activated},TRAF6,TRAF6: TAK1,TRAF6: TAK1 {activated},TRIF,VCAM-1,VEGF,VEGF family,XAF1,XIAP,XIAP: p53: XAF1,c-Jun,c-Jun {activated},c-Jun {nucleus},c-Jun: target genes,carcinoembryonic antigen,caspase,caspase {activated},cd69: S1PR,dsRNA,dsRNA: Mda-5 {activated},dsRNA: Mda-5 {activated}: IPS-1,dsRNA: Mda-5 {activated}: IPS-1: FADD,dsRNA: RIG-1 {activated},dsRNA: RIG-1 {activated}: IPS-1: FADD,dsRNA: RIG-1{activated}: IPS-1,dsRNA:TLR3,dsRNA:TLR3 {p},dsRNA:TLR3: TLR3 {p},dsRNA:TLR3: TLR3 {p}: CD14,dsRNA:TLR3: TLR3 {p}: CD14: TRIF,dsRNA:TLR3: TLR3 {p}: CD14: TRIF: TRAF3: TBK1: IKK-i,dsRNA:TLR3: TLR3 {p}: CD14: TRIF: TRAF3: TBK1: IKK-i: IRF-3,dsRNA:TLR3: TLR3 {p}: CD14: TRIF: TRAF3: TBK1: IKK-i: IRF-3 {p},dsRNA:TLR3: TLR3 {p}: CD14: TRIF: TRAF6: TAK1,dsRNA:TLR3: TLR3 {p}: PI3K,eIF-2alpha,eIF-2alpha {p},eIF3,flagellin,flagellin: TLR5,p38,p38 {activated},p50: RelA-p65: target genes,p50:RelA-p65,p50:RelA-p65 {nucleus},p50:RelA-p65:IkappaB-alpha,p50:RelA-p65:IkappaB-alpha{p},p53,p54,p54: eIF3,p56,p56: eIF3,protein remnants,quinolinamine imiquimod,quinolinamine imiquimod: TLR7: TLR7,ssRNA,ssRNA: TLR7 : TLR7,ssRNA: TLR8: TLR8,tumour-associated antigens
A central issue in dendritic cells (DC) biology is to understand how type I IFNsmodulate the immuno-regulatory properties of DC. In this review I will addressthis issue in light of the recent experimental evidence on the expression andfunction of these cytokines in myeloid DC. This knowledge may have importanttherapeutic implications in infectious and neoplastic diseases and open newperspectives in the use of IFNs as vaccine adjuvants and in the development ofDC-based vaccines.
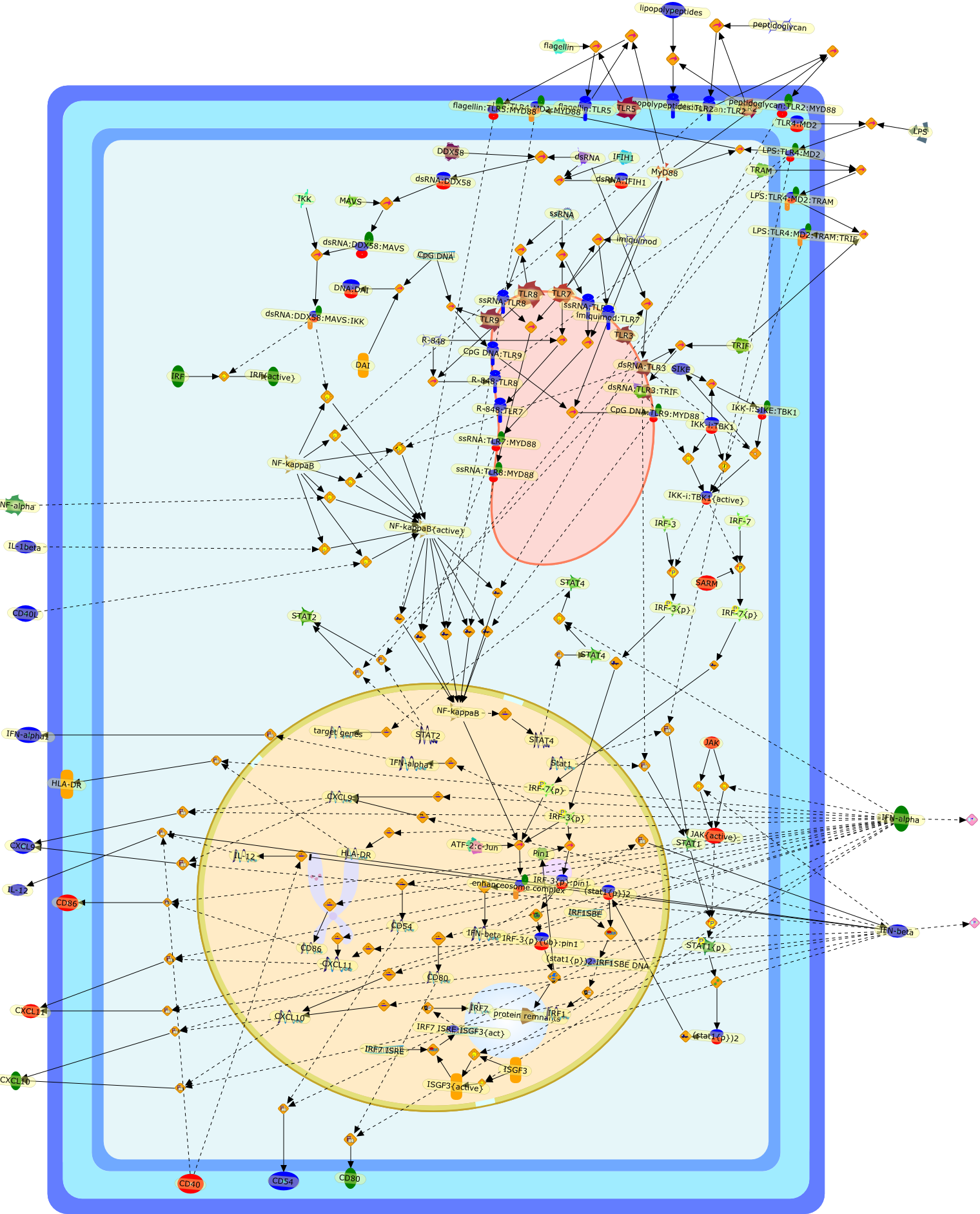{kind=link}
{kind=link}
(stat1{p})2,(stat1{p})2:IRF1SBE DNA,ATF-2:c-Jun,CD40,CD40L,CD54,CD80,CD86,CXCL10,CXCL11,CXCL9,CpG DNA,CpG DNA:TLR9,CpG DNA:TLR9:MYD88,DAI,DDX58,DNA:DAI,HLA-DR,IFIH1,IFN-alpha,IFN-alpha1,IFN-beta,IKK,IKK-i:SIKE:TBK1,IKK-i:TBK1,IKK-i:TBK1{active},IL-12,IL-1beta,IRF,IRF-3,IRF-3{p},IRF-3{p}:pin1,IRF-3{p}{ub}:pin1,IRF-7,IRF-7{p},IRF1,IRF1SBE,IRF7,IRF7 ISRE,IRF7 ISRE:ISGF3{act},IRF{active},ISGF3,ISGF3{active},JAK,JAK{active},LPS,LPS:TLR4:MD2,LPS:TLR4:MD2:MYD88,LPS:TLR4:MD2:TRAM,LPS:TLR4:MD2:TRAM:TRIF,MAVS,MyD88,NF-kappaB,NF-kappaB{active},Pin1,R-848,R-848:TLR7,R-848:TLR8,SARM,SIKE,STAT1,STAT1{p},STAT2,STAT4,Stat1,TLR2,TLR3,TLR4:MD2,TLR5,TLR7,TLR8,TLR9,TNF-alpha,TRAM,TRIF,dsRNA,dsRNA:DDX58,dsRNA:DDX58:MAVS,dsRNA:DDX58:MAVS:IKK,dsRNA:IFIH1,dsRNA:TLR3,dsRNA:TLR3:TRIF,enhanceosome complex,flagellin,flagellin:TLR5,flagellin:TLR5:MYD88,imiquimod,imiquimod:TLR7,lipopolypeptides,lipopolypeptides:TLR2,peptidoglycan,peptidoglycan:TLR2,peptidoglycan:TLR2:MYD88,protein remnants,ssRNA,ssRNA:TLR7,ssRNA:TLR7:MYD88,ssRNA:TLR8,ssRNA:TLR8:MYD88,target genes
During an infection, one of the principal challenges for the host is to detectthe pathogen and activate a rapid defensive response. The Toll-like family ofreceptors (TLRs), among other pattern recognition receptors (PRR), performs thisdetection process in vertebrate and invertebrate organisms. These type Itransmembrane receptors identify microbial conserved structures orpathogen-associated molecular patterns (PAMPs). Recognition of microbialcomponents by TLRs initiates signaling transduction pathways that induce geneexpression. These gene products regulate innate immune responses and furtherdevelop an antigen-specific acquired immunity. TLR signaling pathways areregulated by intracellular adaptor molecules, such as MyD88, TIRAP/Mal, betweenothers that provide specificity of individual TLR- mediated signaling pathways.TLR-mediated activation of innate immunity is involved not only in host defenseagainst pathogens but also in immune disorders. The involvement of TLR-mediatedpathways in auto-immune and inflammatory diseases is described in this reviewarticle.
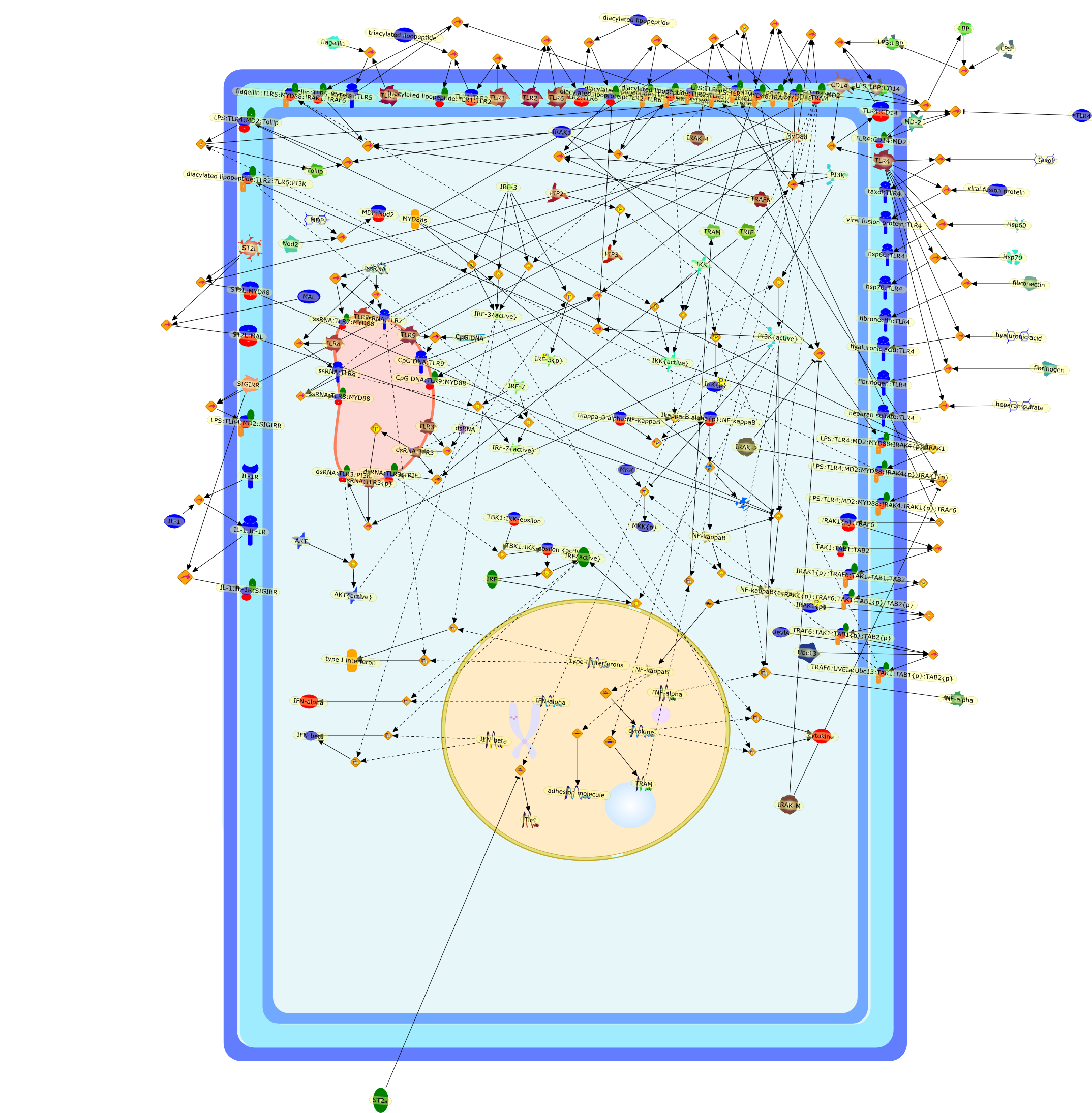{kind=link}
{kind=link}
AKT,AKT{active},CD14,CpG DNA,CpG DNA:TLR9,CpG DNA:TLR9:MYD88,Hsp60,Hsp70,IFN-alpha,IFN-beta,IKK,IKK{active},IKK{p},IL-1,IL-1:IL-1R,IL-1:IL-1R:SIGIRR,IL-1R,IRAK-2,IRAK-4,IRAK-M,IRAK1,IRAK1{p},IRAK1{p}:TRAF6,IRAK1{p}:TRAF6:TAK1:TAB1:TAB2,IRAK1{p}:TRAF6:TAK1:TAB1{p}:TAB2{p},IRF,IRF-3,IRF-3{active},IRF-3{p},IRF-7,IRF-7{active},IRF{active},Ikappa-B alpha:NF-kappaB,Ikappa-B alpha{p}:NF-kappaB,LBP,LPS,LPS:LBP,LPS:LBP:CD14,LPS:TLR4:MD2,LPS:TLR4:MD2:MYD88,LPS:TLR4:MD2:MYD88:IRAK4,LPS:TLR4:MD2:MYD88:IRAK4:IRAK1{p}:TRAF6,LPS:TLR4:MD2:MYD88:IRAK4{p}:IRAK1,LPS:TLR4:MD2:MYD88:IRAK4{p}:IRAK1{p},LPS:TLR4:MD2:SIGIRR,LPS:TLR4:MD2:TRAM,LPS:TLR4:MD2:TRIF:PI3K,LPS:TLR4:MD2:Tollip,LPS:TLR4:MYD88:IRAK4{p},MAL,MD-2,MDP,MDP:Nod2,MKK,MKK{p},MYD88s,MyD88,NF-kappaB,NF-kappaB{active},Nod2,PI3K,PI3K{active},PIP2,PIP3,SIGIRR,ST2L,ST2L:MAL,ST2L:MYD88,ST2s,TAK1:TAB1:TAB2,TBK1:IKK-epsilon,TBK1:IKK-epsilon {active},TLR1,TLR1:TLR2,TLR2,TLR2:TLR6,TLR3,TLR4,TLR4:CD14,TLR4:CD14:MD2,TLR5,TLR6,TLR7,TLR8,TLR9,TNF-alpha,TRAF6,TRAF6:TAK1:TAB1{p}:TAB2{p},TRAF6:UVEIa:Ubc13:TAK1:TAB1{p}:TAB2{p},TRAM,TRIF,Tlr4,Tollip,Ubc13,UevlA,adhesion molecule,cytokine,diacylated lipopeptide,diacylated lipopeptide:TLR2:TLR6:MYD88,diacylated lipopeptide:TLR2:TLR6:MYD88:IRAK1:TRAF6,diacylated lipopeptide:TLR2:TLR6:PI3K,diacylated lipopeptide:TLR2:TLR6:TRIF:PI3K,diacylated lipoprotein:TLR2:TLR6,dsRNA,dsRNA:TLR3,dsRNA:TLR3:PI3K,dsRNA:TLR3:TRIF,dsRNA:TLR3{p},fibrinogen,fibrinogen:TLR4,fibronectin,fibronectin:TLR4,flagellin,flagellin:TLR5,flagellin:TLR5:MYD88,flagellin:TLR5:MYD88:IRAK1:TRAF6,heparan sulfate,heparan sulfate:TLR4,hsp60:TLR4,hsp70:TLR4,hyaluronic acid,hyaluronic acid:TLR4,protein remnants,sTLR4,ssRNA,ssRNA:TLR7,ssRNA:TLR7:MYD88,ssRNA:TLR8,ssRNA:TLR8:MYD88,taxol,taxol:TLR4,triacylated lipopeptide,triacylated lipopeptide:TLR1:TLR2,type I interferon,type I interferons,viral fusion protein,viral fusion protein:TLR4
Current dogma supports the concept that the expression of a disease-inducingsignature cytokine phenotype is important to the maintenance stage of chroniclung disorders. This cytokine phenotype has been characterized as a polarizationtoward type 2 cytokines, which are profibrotic and immunoregulatory. The biologyof this latter activity could mechanistically explain pathogen-inducedexacerbation of chronic lung inflammation, as a skewed cytokine profile in thelung alters dendritic cell function, activates fibroblasts, and facilitates asubsequent "second hit" by an infectious pathogen. In this setting, cytokinebiology is also linked to Toll-like receptors (TLRs) in the maintenance of lungimmunity, as the activity of this receptor-ligand system by both leukocytes andstromal cells is likely an important component of disease chronicity. Theparticipation of dendritic cells via TLRs in chronic lung disease couldfacilitate communication circuits established between antigen-presenting cellsand lymphocytes. Data suggest that TLR activation via myeloid differentiationfactor 88 adaptor protein leads to the induction of a Notch ligand known asDelta-like-4 on dendritic cells that activate the Notch receptor on T cells,promoting a helper T-cell type 1 cytokine response. It is likely that theevolution of host defense signals designed to recognize patterns emitted from ahostile microbial environment may now be superimposed on adaptive immunity andprovide the underpinning to support the maintenance of chronic lung disease.
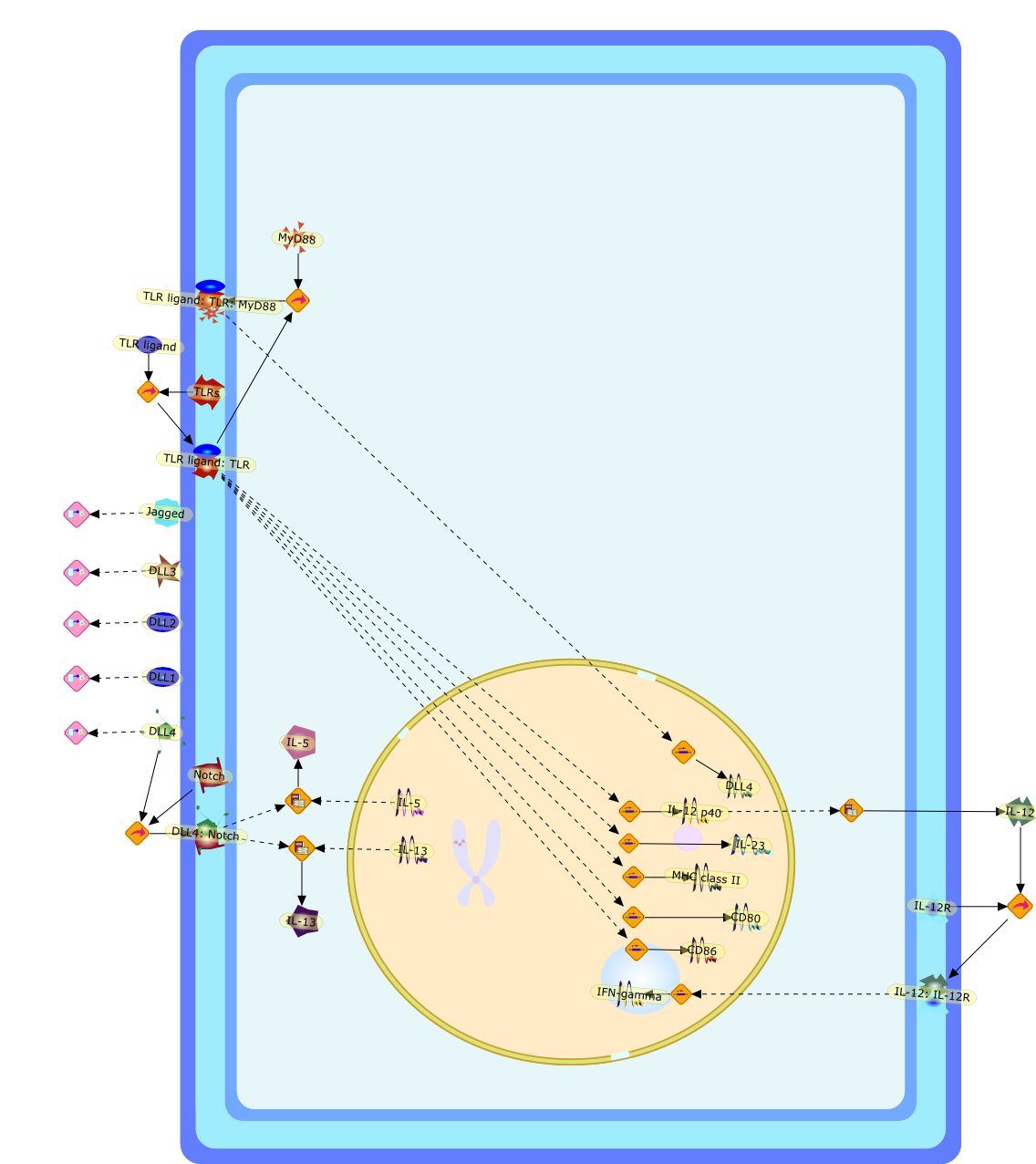{kind=link}
{kind=link}
CD80,CD86,DLL1,DLL2,DLL3,DLL4,DLL4: Notch,IFN-gamma,IL-12,IL-12 p40,IL-12: IL-12R,IL-12R,IL-13,IL-23,IL-5,Jagged,MHC class II,MyD88,Notch,TLR ligand,TLR ligand: TLR,TLR ligand: TLR: MyD88,TLRs
Dendritic cells which are located at the interface of innate and adaptiveimmunity are targets for infection by many different DNA and RNA viruses.Dendritic cell subpopulations express specific nucleic acid recognitionreceptors belonging to the Toll-like receptor family (TLR3, 7, 8, 9) and thecytosolic RNA helicase family (RIG-I, MDA5, LGP2). Activation of dendritic cellsby viral DNA and RNA via these receptors is essential for triggering the innateantiviral immune response and shaping the ensuing adaptive antiviral immunity.This review will summarize our current knowledge of viral nucleic acidrecognition and signaling by Toll-like receptors and RNA helicases focusing onrecent evidence for their specific functions in antiviral defense in vivo.
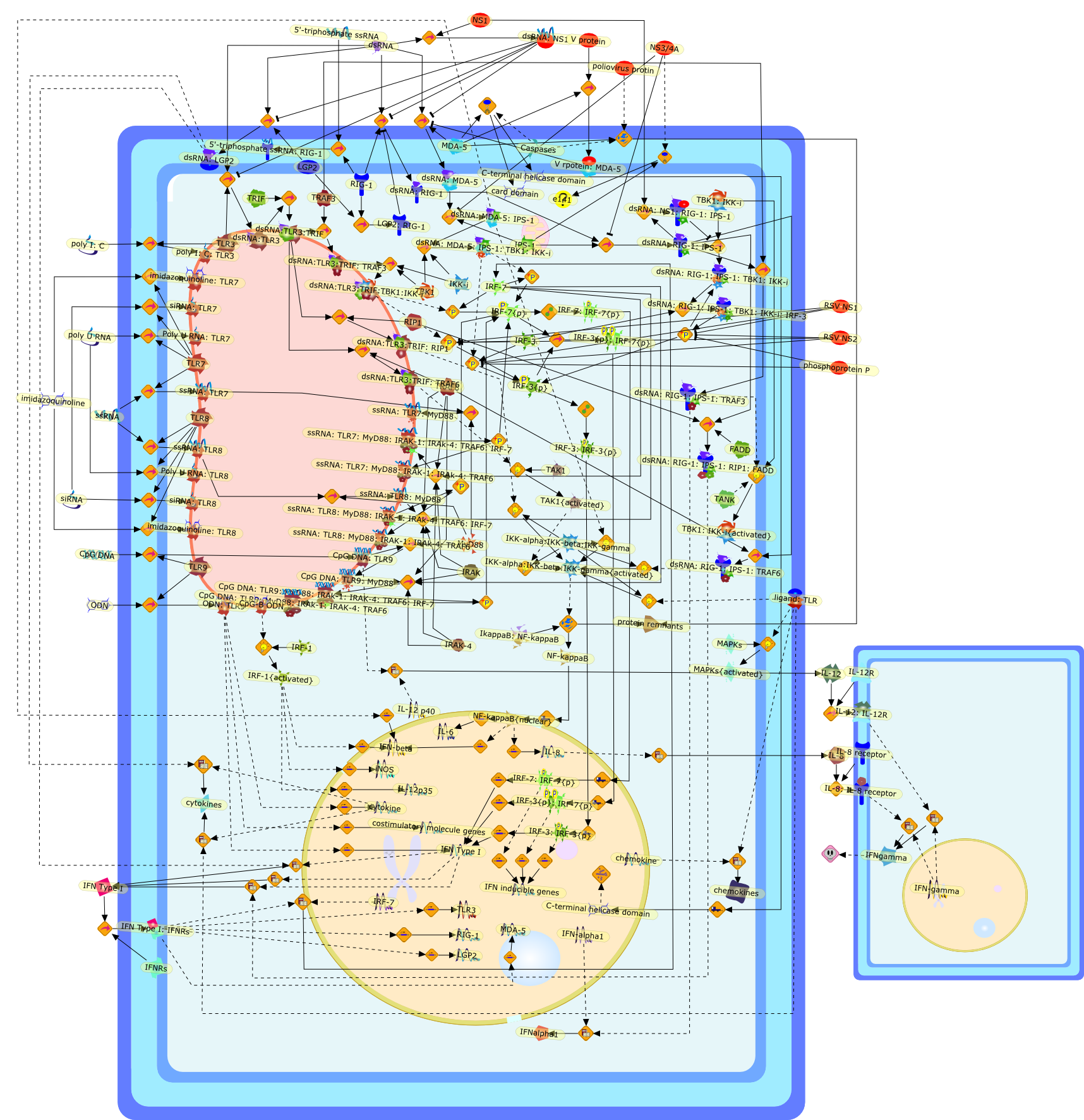{kind=link}
{kind=link}
5'-triphosphate ssRNA,5'-triphosphate ssRNA: RIG-1,C-terminal helicase domain,Caspases,CpG DNA,CpG DNA: TLR9,CpG DNA: TLR9: MyD88,CpG DNA: TLR9: MyD88: IRAk-1: IRAK-4: TRAF6,CpG DNA: TLR9: MyD88: IRAk-1: IRAK-4: TRAF6: IRF-7,CpG-B ODN,FADD,IFN Type I,IFN Type I: IFNRs,IFN inducible genes,IFN-alpha1,IFN-beta,IFN-gamma,IFNRs,IFNalpha1,IFNgamma,IKK-alpha:IKK-beta:IKK-gamma,IKK-alpha:IKK-beta:IKK-gamma{activated},IKK-i,IL-12,IL-12 p40,IL-12: IL-12R,IL-12R,IL-12p35,IL-6,IL-8,IL-8 receptor,IL-8: IL-8 receptor,IPS-1,IRAK,IRAK-4,IRF-1,IRF-1{activated},IRF-3,IRF-3: IRF-3{p},IRF-3{p},IRF-3{p}: IRF-7{p},IRF-7,IRF-7: IRF-7{p},IRF-7{p},IkappaB: NF-kappaB,LGP2,LGP2: RIG-1,MAPKs,MAPKs{activated},MDA-5,MyD88,NF-kappaB,NF-kappaB{nuclear},NS1,NS3/4A,ODN,ODN: TLR9,Poly U RNA: TLR7,Poly U RNA: TLR8,RIG-1,RIP1,RSV NS1,RSV NS2,TAK1,TAK1{activated},TANK,TBK1,TBK1: IKK-i,TBK1: IKK-i{activated},TLR3,TLR7,TLR8,TLR9,TRAF3,TRAF6,TRIF,V protein,V rpotein: MDA-5,card domain,chemokine,chemokines,costimulatory molecule genes,cytokine,cytokines,dsRNA,dsRNA: LGP2,dsRNA: MDA-5,dsRNA: MDA-5: IPS-1,dsRNA: MDA-5: IPS-1: TBK1: IKK-i,dsRNA: NS1,dsRNA: NS1: RIG-1: IPS-1,dsRNA: RIG-1,dsRNA: RIG-1: IPS-1,dsRNA: RIG-1: IPS-1: RIP1: FADD,dsRNA: RIG-1: IPS-1: TBK1: IKK-i,dsRNA: RIG-1: IPS-1: TBK1: IKK-i: IRF-3,dsRNA: RIG-1: IPS-1: TRAF3,dsRNA: RIG-1: IPS-1: TRAF6,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF: RIP1,dsRNA:TLR3:TRIF: TRAF3,dsRNA:TLR3:TRIF: TRAF6,dsRNA:TLR3:TRIF:TBK1:IKK-i,e141,iNOS,imidazoquinoline,imidazoquinoline: TLR7,imidazoquinoline: TLR8,ligand: TLR,phosphoprotein P,poliovirus protin,poly I: C,poly I: C: TLR3,poly U RNA,protein remnants,siRNA,siRNA: TLR7,siRNA: TLR8,ssRNA,ssRNA: TLR7,ssRNA: TLR7: MyD88,ssRNA: TLR7: MyD88: IRAK-1: IRAk-4: TRAF6,ssRNA: TLR7: MyD88: IRAK-1: IRAk-4: TRAF6: IRF-7,ssRNA: TLR8,ssRNA: TLR8: MyD88,ssRNA: TLR8: MyD88: IRAK-1: IRAk-4: TRAF6,ssRNA: TLR8: MyD88: IRAK-1: IRAk-4: TRAF6: IRF-7
Sepsis remains a serious clinical problem because of high patient morbidity andmortality. Despite significant advances in critical care, there is still noefficient causal therapy applicable to patients indicating the need to furtherelucidate the molecular pathways leading to the immunopathology of sepsis. Theimportance of Toll-like receptors (TLR) for the induction of immune responsesagainst sepsis was demonstrated in humans exhibiting polymorphisms in TLR genesand in animal models using genetically modified mouse strains. Because of theclinical heterogeneity in human sepsis and the complex pathomechanismsunderlying sepsis, several different animal models might be used to cover thediverse features of sepsis. TLR receptors induce signaling through the adapterproteins MyD88 and TRIF. TLR signaling is tightly controlled at different stepsof the signaling cascade by series of regulatory proteins. Using a model ofsevere polymicrobial septic peritonitis we could show that single TLRs aredispensable for the induction of innate immune responses under those conditions.However, genetic ablation of MyD88 or TRIF/type-I interferon signaling pathwaysprevented hyper-inflammation and attenuated the pathogenic consequences ofsepsis indicating that dampening common signaling pathways may create a moderatesignal strength which is associated with favorable immune responses. Therefore,broad knowledge about the regulation of TLR-induced signaling pathways mayfurther elucidate the immune mechanisms during sepsis and targeting of TLRadapter molecules may provide a new therapeutic strategy against severe sepsis.
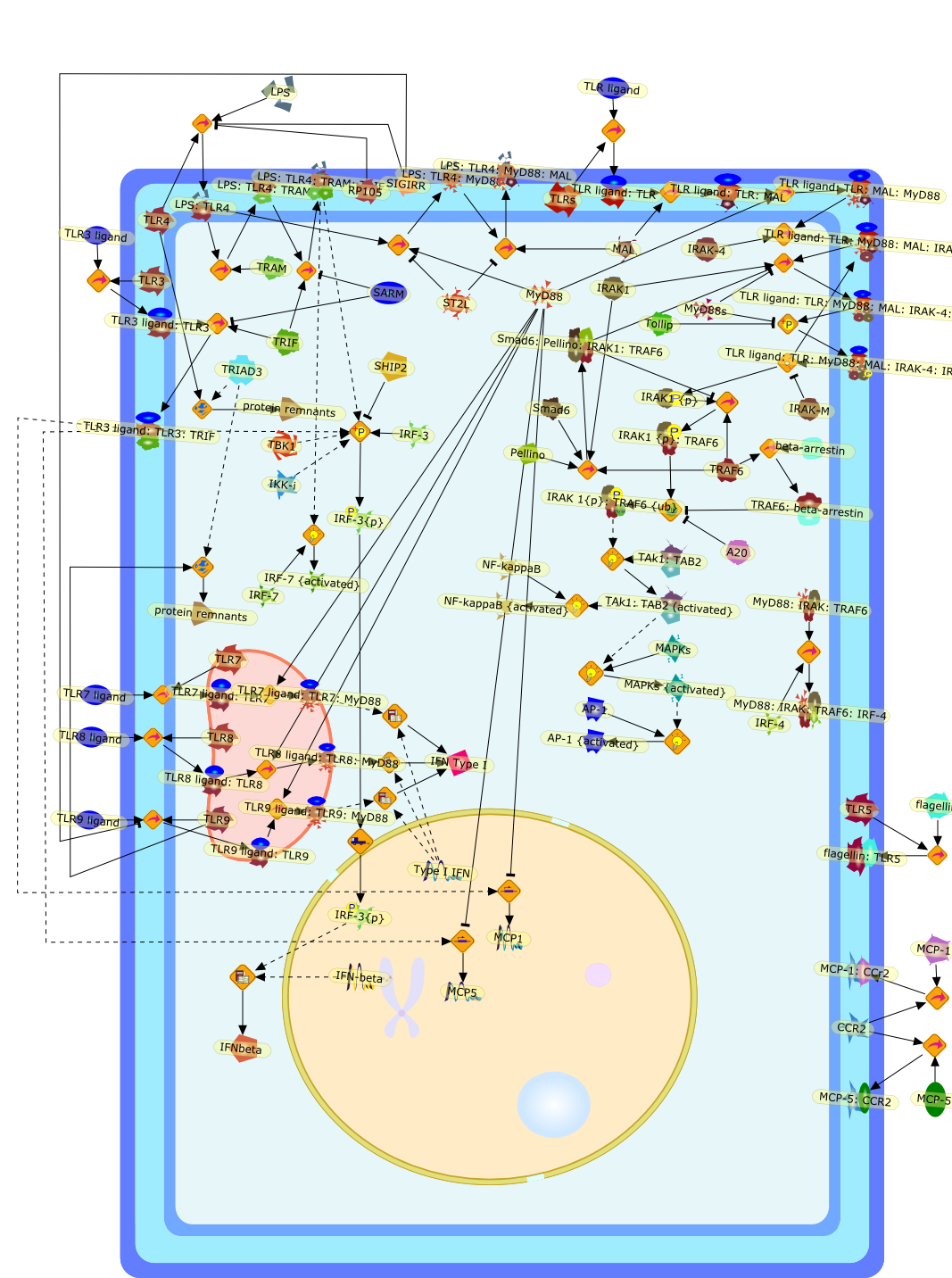{kind=link}
{kind=link}
A20,AP-1,AP-1 {activated},CCR2,IFN Type I,IFN-beta,IFNbeta,IKK-i,IRAK 1{p}: TRAF6 {ub},IRAK-4,IRAK-M,IRAK1,IRAK1 {p},IRAK1 {p}: TRAF6,IRF-3,IRF-3{p},IRF-4,IRF-7,IRF-7 {activated},LPS,LPS: TLR4,LPS: TLR4: MyD88,LPS: TLR4: MyD88: MAL,LPS: TLR4: TRAM,LPS: TLR4: TRAM: TRIF,MAL,MAPKs,MAPKs {activated},MCP-1,MCP-1: CCr2,MCP-5,MCP-5: CCR2,MCP1,MCP5,MyD88,MyD88: IRAK: TRAF6,MyD88: IRAK: TRAF6: IRF-4,MyD88s,NF-kappaB,NF-kappaB {activated},Pellino,RP105,SARM,SHIP2,SIGIRR,ST2L,Smad6,Smad6: Pellino: IRAK1: TRAF6,TAk1: TAB2,TAk1: TAB2 (activated},TBK1,TLR ligand,TLR ligand: TLR,TLR ligand: TLR: MAL,TLR ligand: TLR: MAL: MyD88,TLR ligand: TLR: MyD88: MAL: IRAK-4,TLR ligand: TLR: MyD88: MAL: IRAK-4: IRAK1,TLR ligand: TLR: MyD88: MAL: IRAK-4: IRAK1{p},TLR3,TLR3 ligand,TLR3 ligand: TLR3,TLR3 ligand: TLR3: TRIF,TLR4,TLR5,TLR7,TLR7 ligand,TLR7 ligand: TLR7,TLR7 ligand: TLR7: MyD88,TLR8,TLR8 ligand,TLR8 ligand: TLR8,TLR8 ligand: TLR8: MyD88,TLR9,TLR9 ligand,TLR9 ligand: TLR9,TLR9 ligand: TLR9: MyD88,TLRs,TRAF6,TRAF6: beta-arrestin,TRAM,TRIAD3,TRIF,Tollip,Type I IFN,beta-arrestin,flagellin,flagellin: TLR5,protein remnants
Interferons are pleiotropic cytokines with important proinflammatory functionsrequired in defence against infections with bacteria, viruses and multicellularparasites. In recent years, fundamental functions of interferons in otherprocesses such as cancer immunosurveillance, immune homeostasis andimmunosuppression have been established. In addition, anti-inflammatory roles ofinterferons are well-documented in several inflammatory disease models in themouse, most importantly in experimental autoimmune encephalomyelitis thatresembles multiple sclerosis in humans. While the beneficial effects ofinterferons in such disease models are known, the molecular mechanisms remainpoorly understood. Only recently a few molecular principles for theanti-inflammatory properties of interferons at the cellular level have beenrevealed. They include the ability of interferons to reduce the expression ofthe receptors for the inflammation-related cytokines IL-1 and IL-4, or toincrease the expression of the potent anti-inflammatory genes tristetraprolinand Twist. However, the individual contribution of these anti-inflammatoryresponses to the overall beneficial effects of interferons in inflammatorydiseases is still an open question. Also, the reason for the apparently limitednumber of tissues that are susceptible to the anti-inflammatory functions ofinterferons remains enigmatic. This review summarizes the present knowledge ofthe anti-inflammatory effects of interferons, and describes the currently knownmolecular mechanisms that may help explain the benefits of interferon signallingin several inflammatory diseases.
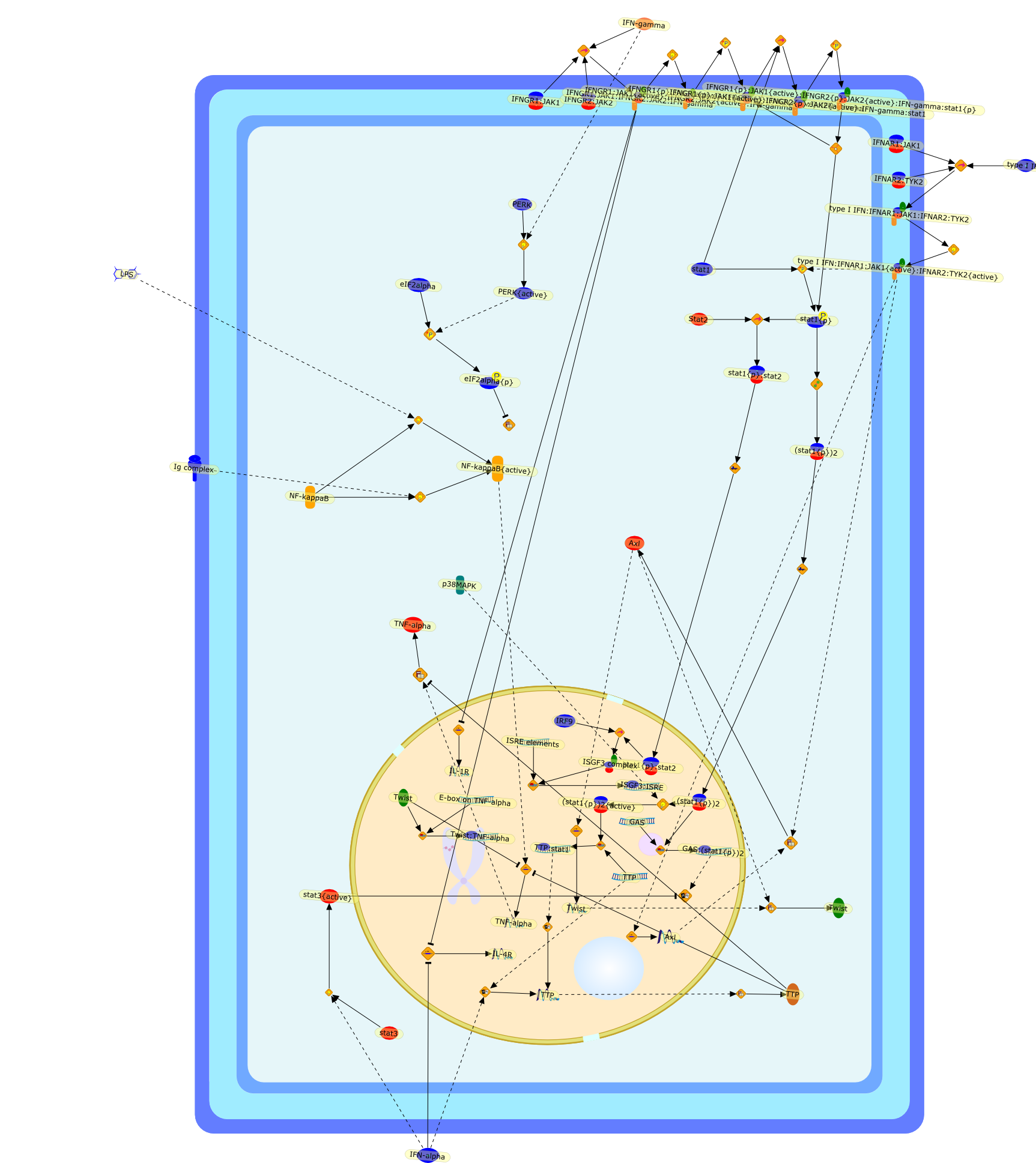{kind=link}
{kind=link}
(stat1{p})2,(stat1{p})2{active},Axl,E-box on TNF-alpha,GAS,GAS:(stat1{p})2,IFN-alpha,IFN-gamma,IFNAR1:JAK1,IFNAR2:TYK2,IFNGR1:JAK1,IFNGR1:JAK1:IFNGR2:JAK2:IFN-gamma,IFNGR1:JAK1{active}:IFNGR2:JAK2{active}:IFN-gamma,IFNGR1{p}:JAK1{active}:IFNGR2{p}:JAK2{active}:IFN-gamma,IFNGR1{p}:JAK1{active}:IFNGR2{p}:JAK2{active}:IFN-gamma:stat1,IFNGR1{p}:JAK1{active}:IFNGR2{p}:JAK2{active}:IFN-gamma:stat1{p},IFNGR2:JAK2,IL-1R,IL-4R,IRF9,ISGF3 complex,ISGF3:ISRE,ISRE elements,Ig complex,LPS,NF-kappaB,NF-kappaB{active},PERK,PERK{active},Stat2,TNF-alpha,TTP,TTP:stat1,Twist,Twist:TNF-alpha,eIF2alpha,eIF2alpha{p},p38MAPK,stat1,stat1{p},stat1{p}:stat2,stat3,stat3{active},type I IFN,type I IFN:IFNAR1:JAK1:IFNAR2:TYK2,type I IFN:IFNAR1:JAK1{active}:IFNAR2:TYK2{active}
Fungal infections represent a significant health burden, especially inimmunocompromised individuals, yet many of the underlying immunologicalmechanisms involved in the recognition and control of these pathogens areunclear. The identification of the Toll-like receptors (TLRs) has shed newinsights on innate microbial recognition and the initiation of immune responses;however, recent evidence indicates that the 'non-TLR' receptors also have asignificant role in these processes, particularly in antifungal immunity. Ofinterest are members of the C-type lectin-receptor family, including the mannosereceptor, dendritic cell-specific intercellular adhesion molecule-3(ICAM-3)-grabbing non-integrin (DC-SIGN), Dectin-1, Dectin-2 and the collectins.Here, we review the roles of each of these receptors, describing how theycontribute to fungal recognition, uptake and killing and also participate in theinduction and/or modulation of the host immune response.
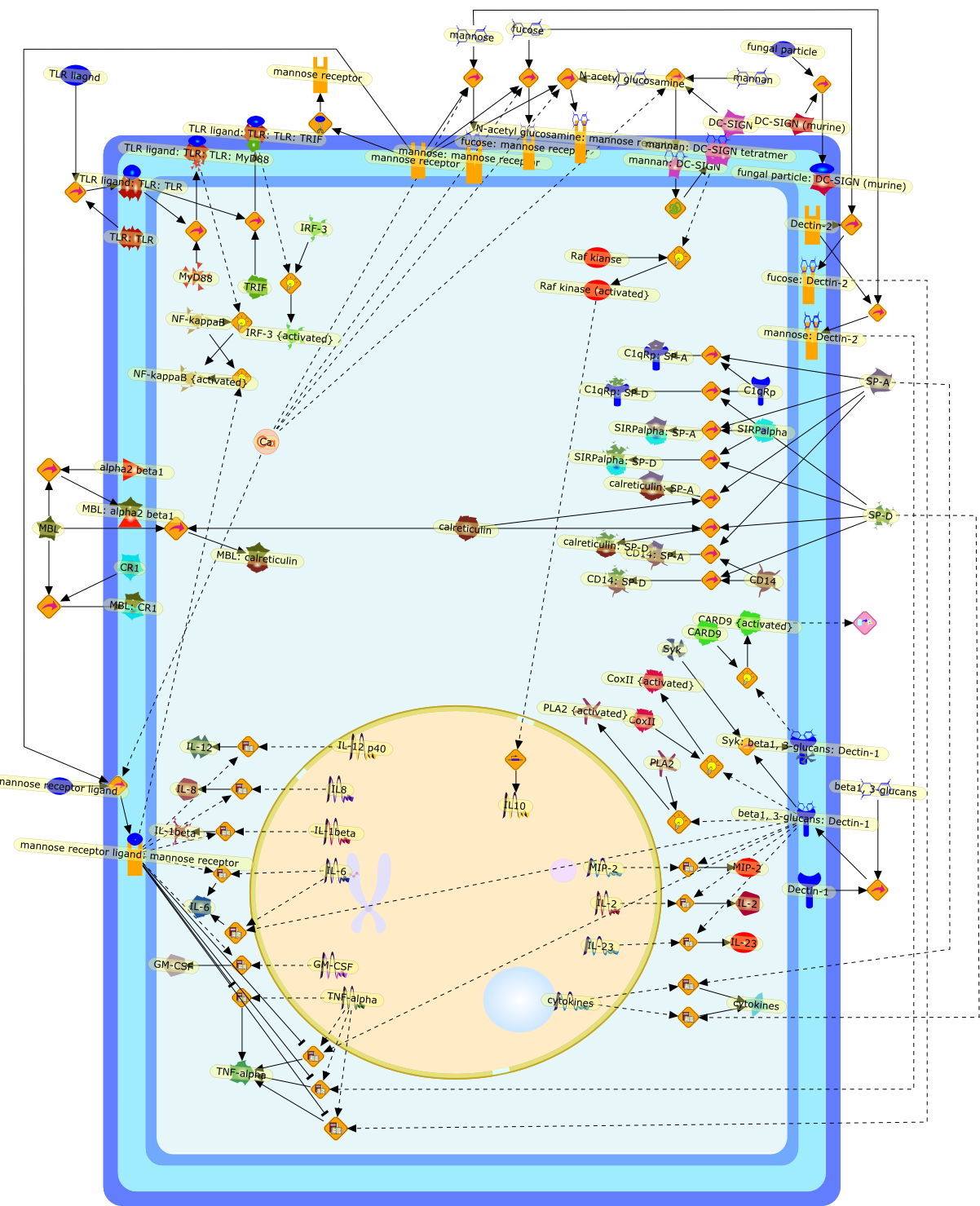{kind=link}
{kind=link}
C1qRp,C1qRp: SP-A,C1qRp: SP-D,CARD9,CARD9 {activated},CD14,CD14: SP-A,CD14: SP-D,CR1,Ca,CoxII,CoxII {activated},DC-SIGN,DC-SIGN (murine),Dectin-1,Dectin-2,GM-CSF,IL-12,IL-12 p40,IL-1beta,IL-2,IL-23,IL-6,IL-8,IL10,IL8,IRF-3,IRF-3 {activated},MBL,MBL: CR1,MBL: alpha2 beta1,MBL: calreticulin,MIP-2,MyD88,N-acetyl glucosamine,N-acetyl glucosamine: mannose receptor,NF-kappaB,NF-kappaB {activated},PLA2,PLA2 {activated},Raf kianse,Raf kinase (activated},SIRPalpha,SIRPalpha: SP-A,SIRPalpha: SP-D,SP-A,SP-D,Syk,Syk: beta1, 3-glucans: Dectin-1,TLR liagnd,TLR ligand: TLR: TLR,TLR ligand: TLR: TLR: MyD88,TLR ligand: TLR: TLR: TRIF,TLR: TLR,TNF-alpha,TRIF,alpha2 beta1,beta1, 3-glucans,beta1, 3-glucans: Dectin-1,calreticulin,calreticulin: SP-A,calreticulin: SP-D,cytokines,fucose,fucose: Dectin-2,fucose: mannose receptor,fungal particle,fungal particle: DC-SIGN (murine),mannan,mannan: DC-SIGN,mannan: DC-SIGN tetratmer,mannose,mannose receptor,mannose receptor ligand,mannose receptor ligand: mannose receptor,mannose: Dectin-2,mannose: mannose receptor
Macrophages are cells that function as a first line of defence against invadingmicroorganisms. One of the hallmarks of macrophages is their ability to becomeactivated in response to exogenous 'danger signals'. Most microbes havemolecular patterns (PAMPS) that are recognized by macrophages and trigger thisactivation response. There are many aspects of the activation response to PAMPSthat are recapitulated when macrophages encounter endogenous danger signals. Inresponse to damaged or stressed self, macrophages undergo physiological changesthat include the initiation of signal transduction cascades fromgermline-encoded receptors, resulting in the elaboration of chemokines,cytokines and toxic mediators. This response to endogenous mediators can enhanceinflammation, and thereby contribute to autoimmune pathologies. Often theoverall inflammatory response is the result of cooperative activation signalsfrom both exogenous and endogenous signals. Macrophage activation plays acritical role, not only in the initiation of the inflammatory response but alsoin the resolution of this response. The clearance of granulocytes and theelaboration of anti-inflammatory mediators by macrophages contribute to thedissolution of the inflammatory response. Thus, macrophages are a key player inthe initiation, propagation and resolution of inflammation. This reviewsummarizes our understanding of the role of macrophages in inflammation. We payparticular attention to the endogenous danger signals that macrophages mayencounter and the responses that these signals induce. The molecular mechanismsresponsible for these responses and the diseases that result frominappropriately controlled macrophage activation are also examined. 2007Pathological Society of Great Britain and Ireland
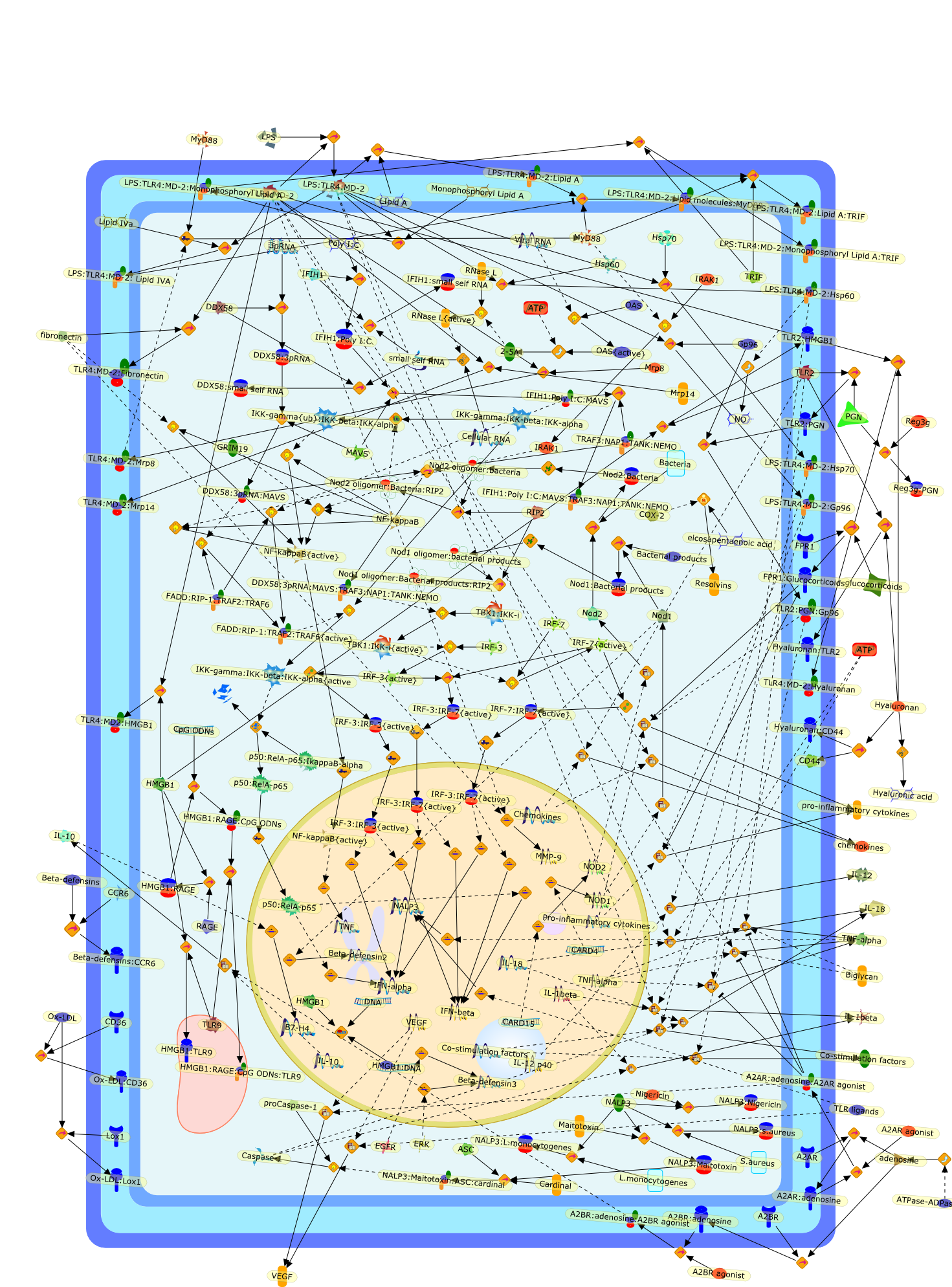{kind=link}
{kind=link}
2-5A,3pRNA,A2AR,A2AR agonist,A2AR:adenosine,A2AR:adenosine:A2AR agonist,A2BR,A2BR agonist,A2BR:adenosine,A2BR:adenosine:A2BR agonist,ASC,ATP,ATPase-ADPase system,B7-H4,Bacteria,Bacterial products,Beta-defensin2,Beta-defensin3,Beta-defensins,Beta-defensins:CCR6,Biglycan,CARD15 ,CARD4,CCR6,CD36,CD44,COX-2,Cardinal,Caspase-1,Cellular RNA,Chemokines,Co-stimulation factors,CpG ODNs,DDX58,DDX58:3pRNA,DDX58:3pRNA:MAVS,DDX58:3pRNA:MAVS:TRAF3:NAP1:TANK:NEMO,DDX58:small self RNA,DNA,EGFR,ERK,FADD:RIP-1:TRAF2:TRAF6,FADD:RIP-1:TRAF2:TRAF6{active},FPR1,FPR1:Glucocorticoids,GRIM19,Gp96,HMGB1,HMGB1:DNA,HMGB1:RAGE,HMGB1:RAGE:CpG ODNs,HMGB1:RAGE:CpG ODNs:TLR9,HMGB1:TLR9,Hsp60,Hsp70,Hyaluronan,Hyaluronan:CD44,Hyaluronan:TLR2,Hyaluronic acid,IFIH1,IFIH1:Poly I:C,IFIH1:Poly I:C:MAVS,IFIH1:Poly I:C:MAVS:TRAF3:NAP1:TANK:NEMO,IFIH1:small self RNA,IFN-alpha,IFN-beta,IKK-gamma:IKK-beta:IKK-alpha,IKK-gamma:IKK-beta:IKK-alpha{active,IKK-gamma{ub}:IKK-beta:IKK-alpha,IL-10,IL-12,IL-12 p40,IL-18,IL-1beta,IRAK1,IRF-3,IRF-3:IRF-3{active},IRF-3:IRF-7{active},IRF-3{active},IRF-7,IRF-7:IRF-7{active},IRF-7{active},L.monocytogenes,LIpid A,LPS,LPS:TLR4:MD-2,LPS:TLR4:MD-2: Lipid IVA,LPS:TLR4:MD-2:Gp96,LPS:TLR4:MD-2:Hsp60,LPS:TLR4:MD-2:Hsp70,LPS:TLR4:MD-2:Lipid A,LPS:TLR4:MD-2:Lipid A:TRIF,LPS:TLR4:MD-2:Lipid molecules:MyD88,LPS:TLR4:MD-2:Monophosphoryl Lipid A,LPS:TLR4:MD-2:Monophosphoryl Lipid A:TRIF,Lipid IVa,Lox1,MAVS,MMP-9,Maitotoxin,Monophosphoryl Lipid A,Mrp14,Mrp8,MyD88,NALP3,NALP3:L.monocytogenes,NALP3:Maitotoxin,NALP3:Maitotoxin:ASC:cardinal,NALP3:Nigericin,NALP3:s.aureus,NF-kappaB,NF-kappaB{active},NO,NOD1,NOD2,Nigericin,Nod1,Nod1 oligomer:Bacterial products:RIP2,Nod1 oligomer:bacterial products,Nod1:Bacterial products,Nod2,Nod2 oligomer:Bacteria:RIP2,Nod2 oligomer:bacteria,Nod2:Bacteria,OAS,OAS{active},Ox-LDL,Ox-LDL:CD36,Ox-LDL:Lox1,PGN,Poly I:C,Pro-inflammatory cytokines,RAGE,RIP2,RNase L,RNase L{active},Reg3g,Reg3g:PGN,Resolvins,S.aureus,TBK1:IKK-i,TBK1:IKK-i{active},TLR ligands,TLR2,TLR2:HMGB1,TLR2:PGN,TLR2:PGN:Gp96,TLR4:MD-2,TLR4:MD-2:Fibronectin,TLR4:MD-2:Hyaluronan,TLR4:MD-2:Mrp14,TLR4:MD-2:Mrp8,TLR4:MD2:HMGB1,TLR9,TNF,TNF-alpha,TRAF3:NAP1:TANK:NEMO,TRIF,VEGF,Viral RNA,adenosine,chemokines,degradants,eicosapentaenoic acid,fibronectin,glucocorticoids,p50:RelA-p65,p50:RelA-p65:IkappaB-alpha,pro-inflammatory cytokines,proCaspase-1,small self RNA
Cytokines like interferons (IFNs) play a central role in regulating innate andspecific immunities against the pathogens and neoplastic cells. A number ofsignaling pathways are induced in response to IFN in various cells. One classicmechanism employed by IFNs is the JAK-STAT signaling pathway for inducingcellular responses. Here we describe the non-STAT pathways that participate inIFN-induced responses. In particular, we will focus on the role played bytranscription factor C/EBP-beta in mediating these responses.
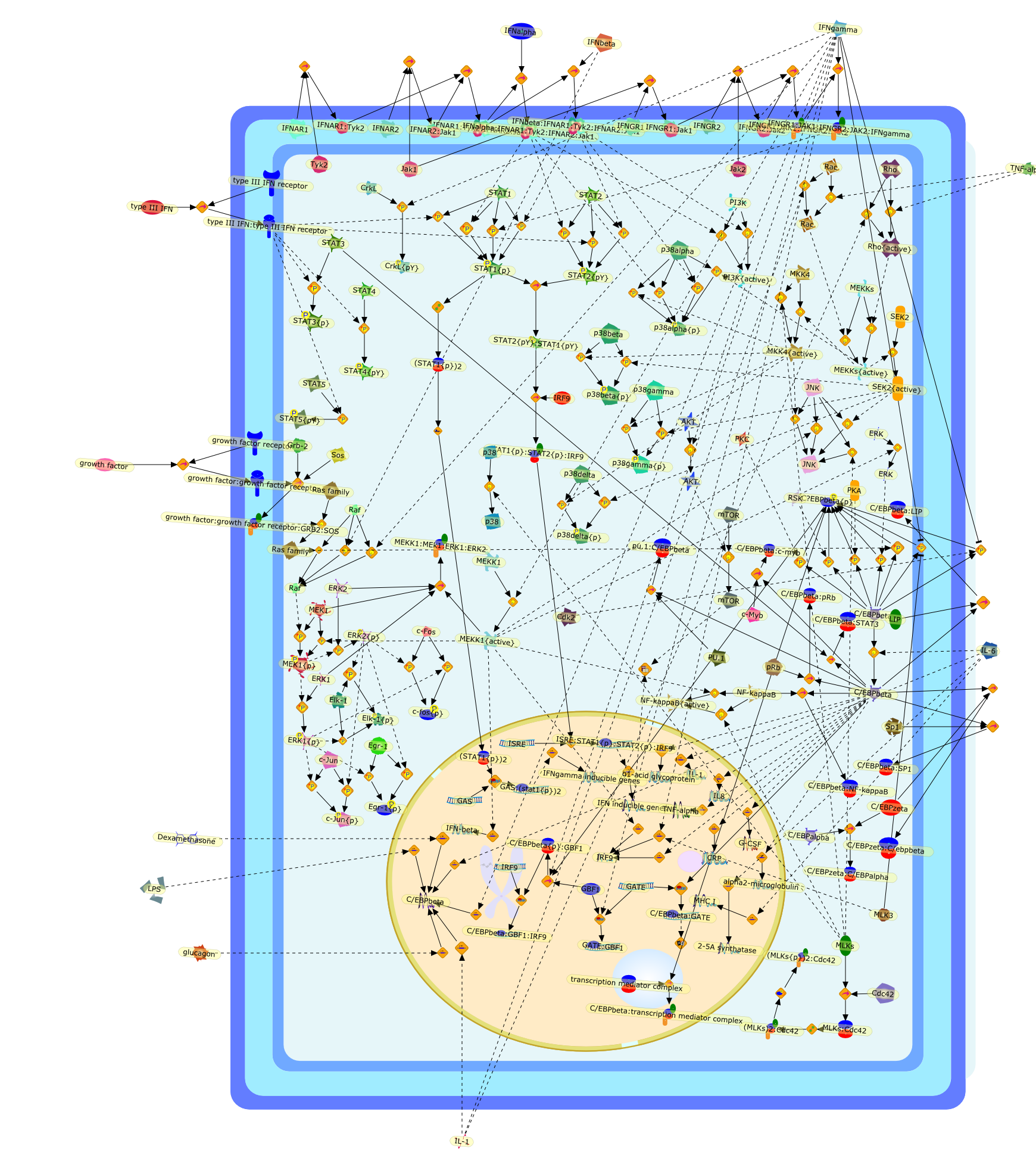{kind=link}
{kind=link}
(MLKs)2:Cdc42,(MLKs{p})2:Cdc42,(STAT1{p})2,2-5A synthatase,AKT,C/EBPalpha,C/EBPbeta,C/EBPbeta:GATE,C/EBPbeta:GBF1:IRF9,C/EBPbeta:LIP,C/EBPbeta:NF-kappaB,C/EBPbeta:SP1,C/EBPbeta:STAT3,C/EBPbeta:c-myb,C/EBPbeta:pRb,C/EBPbeta:transcription mediator complex,C/EBPbeta{p}:GBF1,C/EBPzeta,C/EBPzeta:C/EBPalpha,C/EBPzeta:C/ebpbeta,C?EBPbeta{p},CRP,Cdc42,Cdk2,CrkL,CrkL{pY},Dexamethasone,ERK,ERK1,ERK1{p},ERK2,ERK2{p},Egr-1,Egr-1{p},Elk-1,Elk-1{p},G-CSF,GAS,GAS:(stat1{p})2,GATE,GATE:GBF1,GBF1,Grb-2,IFN inducible genes,IFN-beta,IFNAR1,IFNAR1:Tyk2,IFNAR1:Tyk2:IFNAR2:Jak1,IFNAR2,IFNAR2:Jak1,IFNGR1,IFNGR1:JAK1:IFNGR2:JAK2,IFNGR1:JAK1:IFNGR2:JAK2:IFNgamma,IFNGR1:Jak1,IFNGR2,IFNGR2:Jak2,IFNalpha,IFNalpha:IFNAR1:Tyk2:IFNAR2:Jak1,IFNbeta,IFNbeta:IFNAR1:Tyk2:IFNAR2:Jak1,IFNgamma,IFNgamma inducible genes,IL-1,IL-6,IL8,IRF9,ISRE,ISRE:STAT1{p}:STAT2{p}:IRF9,JNK,Jak1,Jak2,LIP,LPS,MEK1,MEK1{p},MEKK1,MEKK1:MEk1:ERK1:ERK2,MEKK1{active},MEKKs,MEKKs{active},MHC I,MKK4,MKK4{active},MLK3,MLKs,MLKs:Cdc42,NF-kappaB,NF-kappaB{active},PI3K,PI3K{active},PKA,PKC,PU.1,RSK,Rac,Raf,Ras family,Rho,Rho{active},SEK2,SEK2{active},STAT1,STAT1{p},STAT1{p}:STAT2{p}:IRF9,STAT2,STAT2{pY},STAT2{pY}:STAT1{pY},STAT3,STAT3{p},STAT4,STAT4{pY},STAT5,STAT5{pY},Sos,Sp1,TNF-alpha,Tyk2,alpha2-microglobulin,c-Fos,c-Jun,c-Jun{p},c-Myb,c-fos{p},glucagon,growth factor,growth factor receptor,growth factor:growth factor receptor,growth factor:growth factor receptor:GRB2:SOS,mTOR,p38,p38alpha,p38alpha{p},p38beta,p38beta{p},p38delta,p38delta{p},p38gamma,p38gamma{p},pRb,pu.1:C/EBPbeta,transcription mediator complex,type III IFN,type III IFN receptor,type III IFN:type III IFN receptor,�Ύ�1-acid glycoprotein
Recent studies indicate that TNF (tumor necrosis factor) receptor signalling isa key player in HIV infection. HIV proteins have been shown to target TNFreceptor signalling, leading both to apoptosis of uninfected bystander T cellsand to sustained viral replication in infected T cells and macrophages. Thisarticle proposes a model that highlights the role of HIV proteins in themodulation of TNF receptor signalling and could explain both immune suppressionand the formation of viral reservoirs during HIV infection.
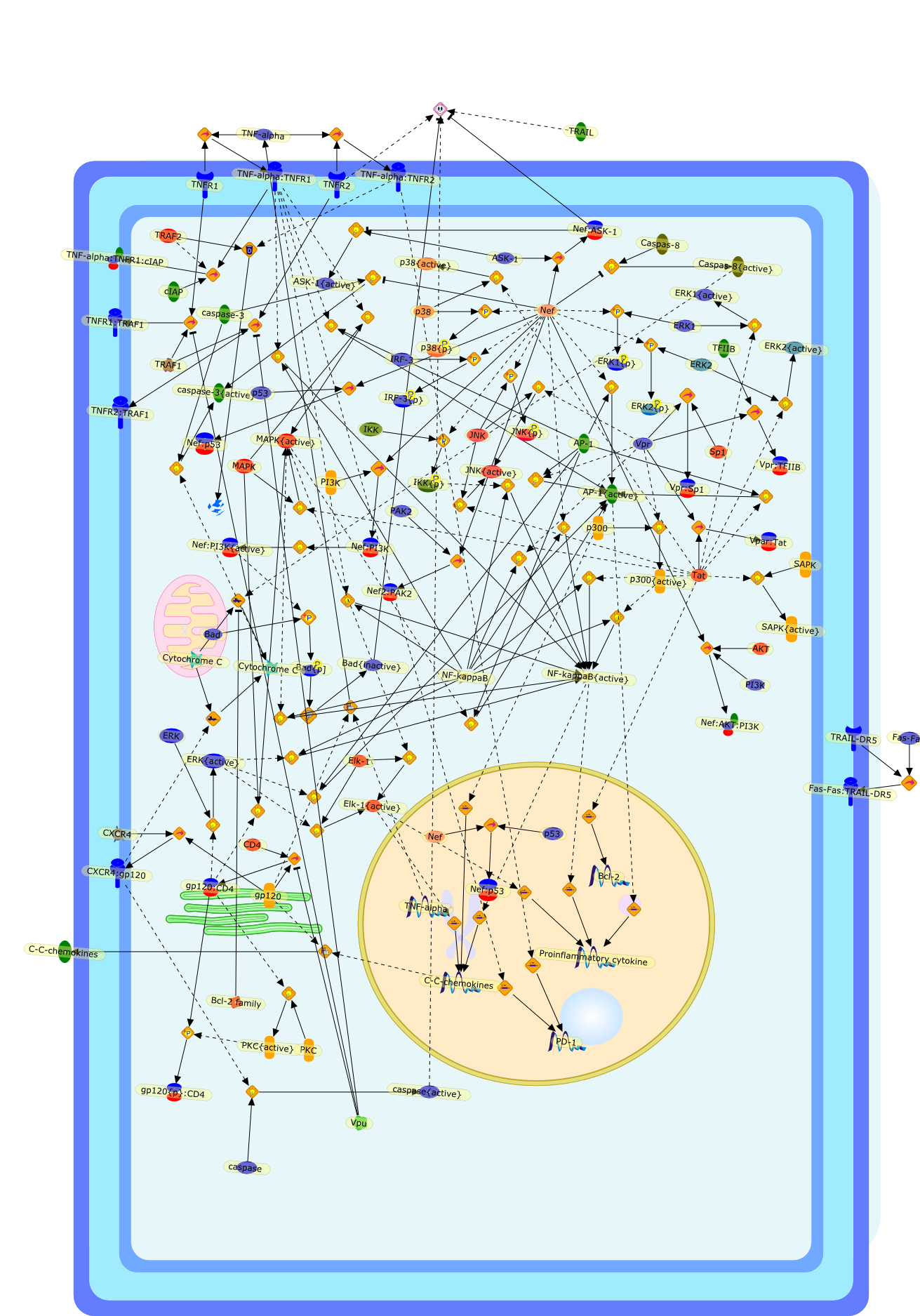{kind=link}
{kind=link}
AKT,AP-1,AP-1{active},ASK-1,ASK-1{active},Bad,Bad{inactive},Bad{p],Bcl-2,Bcl-2 family,C-C-chemokines,CD4,CXCR4,CXCR4:gp120,Caspas-8,Caspas-8{active},Cytochrome C,ERK,ERK1,ERK1{active},ERK1{p},ERK2,ERK2{active},ERK2{p},ERK{active},Elk-1,Elk-1{active},Fas-Fas,Fas-Fas:TRAIL-DR5,IKK,IKK{p},IRF-3,IRF-3{p},JNK,JNK{active},JNK{p},MAPK,MAPK{active},NF-kappaB,NF-kappaB{active},Nef,Nef2:PAK2,Nef:AKT:PI3K,Nef:ASK-1,Nef:PI3K,Nef:PI3K{active},Nef:p53,PAK2,PD-1,PI3K,PKC,PKC{active},Proinflammatory cytokine,SAPK,SAPK{active},Sp1,TFIIB,TNF-alpha,TNF-alpha:TNFR1,TNF-alpha:TNFR1:cIAP,TNF-alpha:TNFR2,TNFR1,TNFR1:TRAF1,TNFR2,TNFR2:TRAF1,TRAF1,TRAF2,TRAIL,TRAIL-DR5,Tat,Vpar:Tat,Vpr,Vpr:Sp1,Vpr:TFIIB,Vpu,cIAP,caspase,caspase-3,caspase-3{active},caspase{active},degradants,gp120,gp120:CD4,gp120{p}:CD4,p300,p300{active},p38,p38{active},p38{p},p53
Controversy exists concerning the role of Toll-like receptors and MyD88 inimmunity to tuberculosis (TB). This mini-review argues that (i) Toll-likereceptors are not essential for an effective immune response against TB, (ii)MyD88 is essential, but not because it transduces signals from TLRs, (iii)adaptive immunity to TB is largely TLR/MyD88-independent. Some of thediscrepancies may be resolved by cogent attribution of distinct immune functionsto the individual components of the TLR/MyD88 system. In mice, TLRs and MyD88are fully dispensable in sensing Mtb infection and instructing T cell-mediatedadaptive immunity, and while TLRs are also redundant during macrophage effectorimmunity, MyD88 is essential for efficient killing of mycobacteria. Thisdistinction should help to molecularly pinpoint the MyD88-dependent, yetTLR-independent critical mechanisms of macrophage activation involved inintracellular growth restriction of Mtb. Disrupted IL-1R and/or IFN-gammasignaling pathways likely play a much more prominent role in explaining theexquisite susceptibility of MyD88-deficient mice to TB than the function ofMyD88 as a TLR adaptor.
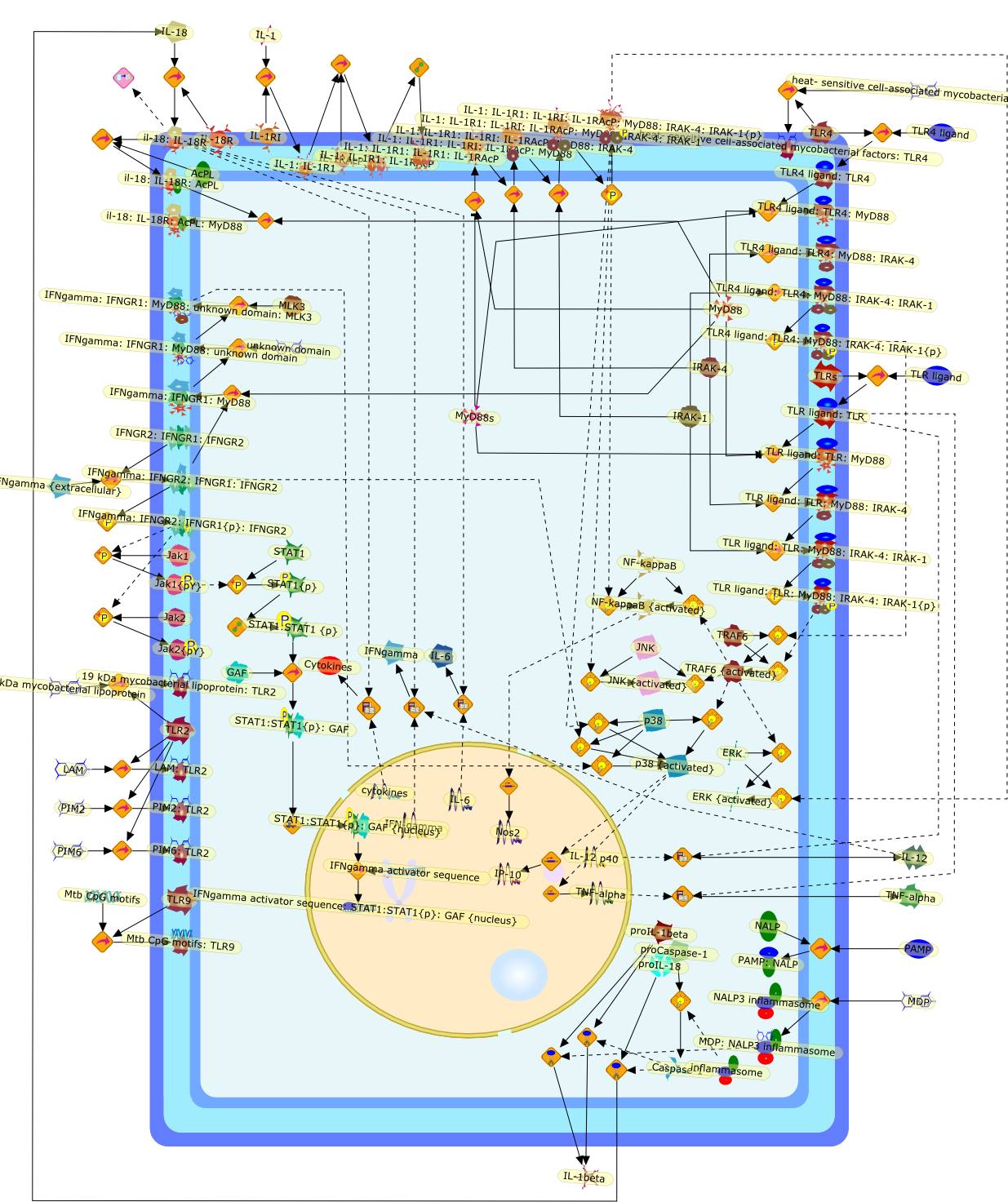{kind=link}
{kind=link}
19 kDa mycobacterial lipoprotein,19 kDa mycobacterial lipoprotein: TLR2,AcPL,Caspase-1,Cytokines,ERK,ERK {activated},GAF,IFN-gamma,IFNGR2: IFNGR1: IFNGR2,IFNgamma,IFNgamma activator sequence,IFNgamma activator sequence: STAT1:STAT1{p}: GAF {nucleus},IFNgamma {extracellular},IFNgamma: IFNGR1: MyD88,IFNgamma: IFNGR1: MyD88: unknown domain,IFNgamma: IFNGR1: MyD88: unknown domain: MLK3,IFNgamma: IFNGR2: IFNGR1: IFNGR2,IFNgamma: IFNGR2: IFNGR1{p}: IFNGR2,IL-1,IL-12,IL-12 p40,IL-18,IL-18R,IL-1: IL-1R1,IL-1: IL-1R1: IL-1R1: IL-1RAcP,IL-1: IL-1R1: IL-1RAcP,IL-1: IL-1R1: IL-1RI: IL-1RAcP: MyD88,IL-1: IL-1R1: IL-1RI: IL-1RAcP: MyD88: IRAK-4,IL-1: IL-1R1: IL-1RI: IL-1RAcP: MyD88: IRAK-4: IRAK-1,IL-1: IL-1R1: IL-1RI: IL-1RAcP: MyD88: IRAK-4: IRAK-1{p},IL-1RAcP,IL-1RI,IL-1beta,IL-6,IP-10,IRAK-1,IRAK-4,JNK,JNK {activated},Jak1,Jak1{pY},Jak2,Jak2{pY},LAM,LAM: TLR2,MDP,MDP: NALP3 inflammasome,MLK3,Mtb CpG motifs,Mtb CpG motifs: TLR9,MyD88,MyD88s,NALP,NALP3 inflammasome,NF-kappaB,NF-kappaB {activated},Nos2,PAMP,PAMP: NALP,PIM2,PIM2: TLR2,PIM6,PIM6: TLR2,STAT1,STAT1:STAT1 {p},STAT1:STAT1{p}: GAF ,STAT1:STAT1{p}: GAF {nucleus},STAT1{p},TLR ligand,TLR ligand: TLR,TLR ligand: TLR: MyD88,TLR ligand: TLR: MyD88: IRAK-4,TLR ligand: TLR: MyD88: IRAK-4: IRAK-1,TLR ligand: TLR: MyD88: IRAK-4: IRAK-1{p},TLR2,TLR4,TLR4 ligand,TLR4 ligand: TLR4,TLR4 ligand: TLR4: MyD88,TLR4 ligand: TLR4: MyD88: IRAK-4,TLR4 ligand: TLR4: MyD88: IRAK-4: IRAK-1,TLR4 ligand: TLR4: MyD88: IRAK-4: IRAK-1{p},TLR9,TLRs,TNF-alpha,TRAF6,TRAF6 {activated},cytokines,heat- sensitive cell-associated mycobacterial factors,heat- sensitive cell-associated mycobacterial factors: TLR4,il-18: IL-18R,il-18: IL-18R: AcPL,il-18: IL-18R: AcPL: MyD88,inflammasome,p38,p38 {activated},proCaspase-1,proIL-18,proIL-1beta,unknown domain
Inhalation of the crystalline form of silica is associated with a variety ofpathologies, from acute lung inflammation to silicosis, in addition toautoimmune disorders and cancer. Basic science investigators looking at themechanisms involved with the earliest initiators of disease are focused on howthe alveolar macrophage interacts with the inhaled silica particle and theconsequences of silica-induced toxicity on the cellular level. Based onexperimental results, several rationales have been developed for exactly howcrystalline silica particles are toxic to the macrophage cell that isfunctionally responsible for clearance of the foreign particle. For example,silica is capable of producing reactive oxygen species (ROS) either directly (onthe particle surface) or indirectly (produced by the cell as a response tosilica), triggering cell-signaling pathways initiating cytokine release andapoptosis. With murine macrophages, reactive nitrogen species are produced inthe initial respiratory burst in addition to ROS. An alternative explanation forsilica toxicity includes lysosomal permeability, by which silica disrupts thenormal internalization process leading to cytokine release and cell death. Stillother research has focused on the cell surface receptors (collectively known asscavenger receptors) involved in silica binding and internalization. Thesilica-induced cytokine release and apoptosis are described as the function ofreceptor-mediated signaling rather than free radical damage. Current researchideas on silica toxicity and binding in the alveolar macrophage are reviewed anddiscussed.
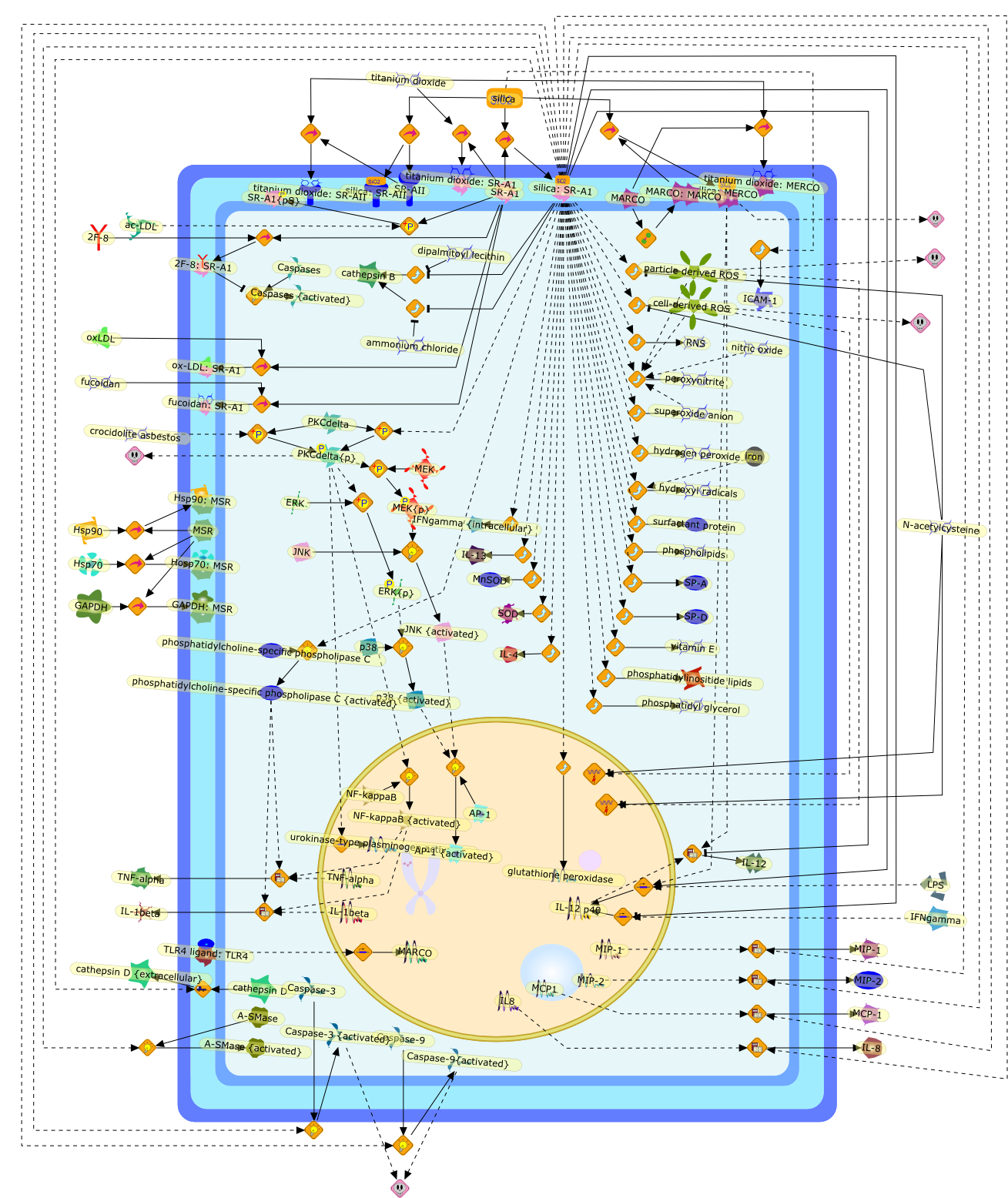{kind=link}
{kind=link}
2F-8,2F-8: SR-A1,A-SMase,A-SMase {activated},AP-1,AP-1 {activated},Caspase-3,Caspase-3 {activated},Caspase-9,Caspase-9{activated},Caspases,Caspases {activated},ERK,ERK{p},GAPDH,GAPDH: MSR,Hosp70: MSR,Hsp70,Hsp90,Hsp90: MSR,ICAM-1,IFNgamma,IFNgamma {intracellular},IL-12,IL-12 p40,IL-13,IL-1beta,IL-4,IL-8,IL8,JNK,JNK {activated},LPS,MARCO,MARCO: MARCO,MCP-1,MCP1,MEK,MEK{p},MIP-1,MIP-2,MSR,MnSOD,N-acetylcysteine,NF-kappaB,NF-kappaB {activated},PKCdelta,PKCdelta{p},RNS,SOD,SP-A,SP-D,SR-A1,SR-A1{pS},SR-AII,TLR4 ligand: TLR4,TNF-alpha,ac-LDL,ammonium chloride,cathepsin B,cathepsin D,cathepsin D {extracellular},cell-derived ROS,crocidolite asbestos,dipalmitoyl lecithin,fucoidan,fucoidan: SR-A1,glutathione peroxidase,hydrogen peroxide,hydroxyl radicals,iron,nitric oxide,ox-LDL: SR-A1,oxLDL,p38,p38 {activated},particle derived ROS,peroxynitrite,phosphatidyl glycerol,phosphatidylcholine-specific phospholipase C,phosphatidylcholine-specific phospholipase C {activated},phosphatidylinositide lipids,phospholipids,silica,silica: MERCO,silica: SR-A1,silica: SR-AII,superoxide anion,surfactant protein,titanium dioxide,titanium dioxide: MERCO,titanium dioxide: SR-A1,titanium dioxide: SR-AII,urokinase-type plasminogen activator,vitamin E
The immune response induced by the pathogen-associated-pattern recognitionreceptor toll-like receptor 9 (TLR9) upon binding of CpG motif-containing DNAhas been widely accepted as an important pathway in the immune defense againstmicrobial pathogens. In contrast, the role of TLR9 in anti-DNA antibodygeneration and the pathogenesis of systemic lupus erythematosus (SLE) remainscontroversial. Indeed, the in vivo situation might consist of a delicate balancebetween B-cell receptor and DNA receptor signaling. Most surprisingly, TLR9deletion does not ameliorate but rather exacerbates pathology in murine models.Such observations warrant caution with therapeutic efforts to treat autoimmunediseases, especially SLE, via TLR modulation.
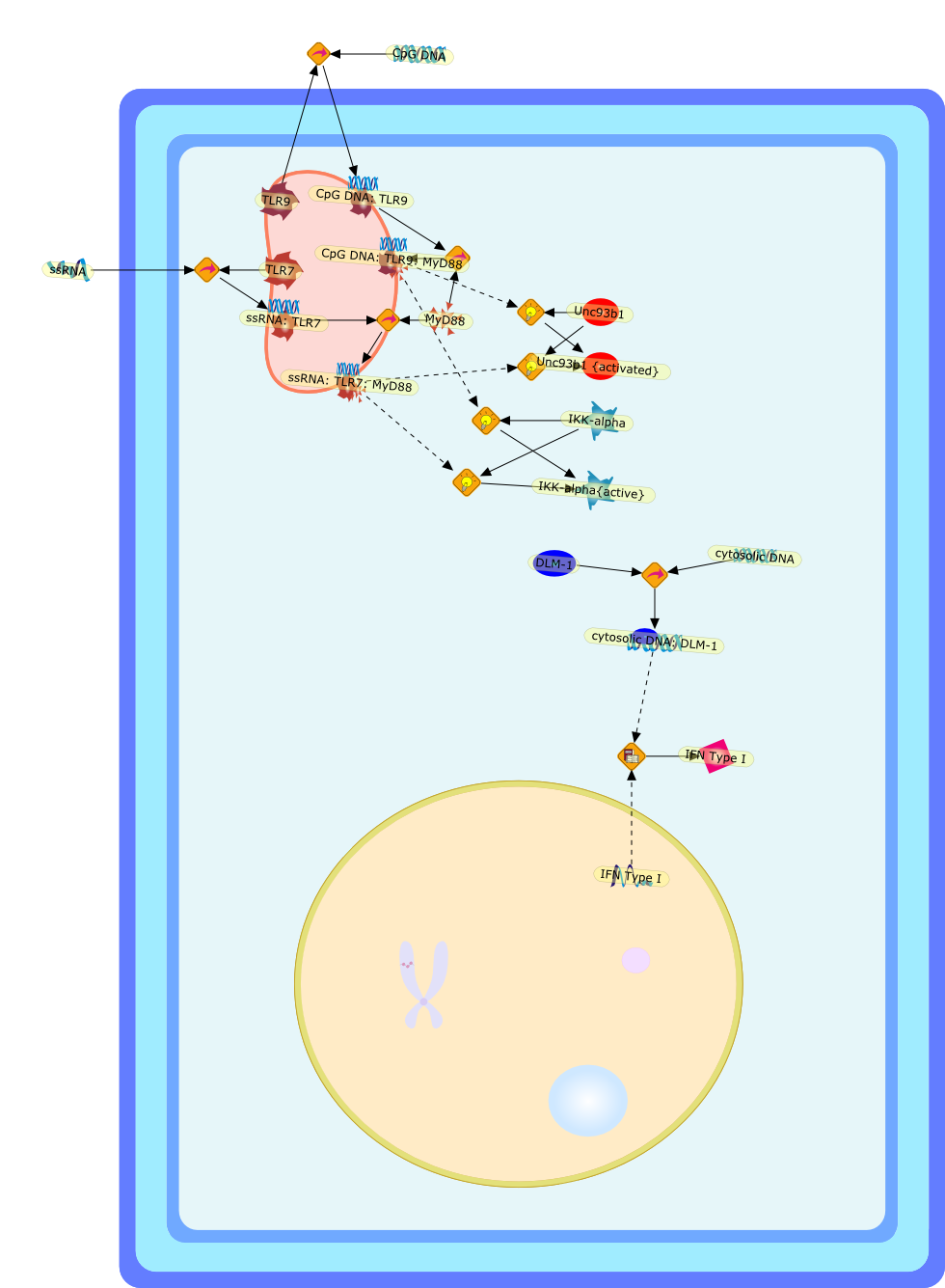{kind=link}
{kind=link}
CpG DNA,CpG DNA: TLR9,CpG DNA: TLR9: MyD88,DLM-1,IFN Type I,IKK-alpha,IKK-alpha{active},MyD88,TLR7,TLR9,Unc93b1,Unc93b1 {activated},cytosolic DNA,cytosolic DNA: DLM-1,ssRNA,ssRNA: TLR7,ssRNA: TLR7: MyD88
Classically, the extracellular matrix (ECM) was viewed as a supporting structurefor stabilizing the location of cells in tissues and for preserving thearchitecture of tissues. This conception has changed dramatically over the pastfew decades with discoveries that ECM has profound influences on the structure,viability, and functions of cells. Much of the data supporting this new paradigmhas been obtained from studies of normal and pathological structural cells suchas fibroblasts, smooth muscle cells, and malignant cells, as, for example,breast cancer epithelial cells. However, there has also been recognition thateffects of ECM on cells extend to inflammatory cells. In this context, attentionhas been drawn to fragments of ECM components. In this review, we presentinformation supporting the concept that proteolytic fragments of ECM affectmultiple functions and properties of inflammatory and immune cells. Our focus isparticularly upon neutrophils, monocytes, and macrophages and fragments derivedfrom collagens, elastin, and laminins. Hyaluronan fragments, although they arenot products of proteolysis, are also discussed, as they are a notable exampleof ECM fragments that exhibit important effects on inflammatory cells. Further,we summarize some exciting recent developments in this field as a result ofmouse models in which defined ECM fragments and their receptors are clearlyimplicated in inflammation in vivo. Thus, this review underscores the idea thatproteolysis of ECM may well have implications that go beyond modifying thestructural environment of cells and tissues.
{kind=link}
{kind=link}
ASKVKV,ASKVKV receptor,ASKVKV: ASKVKV receptor,CD44,DNA,EBP,IFN-gamma,IFNgamma,IL-1,IL-12,IL-6,IL-8,IL12,IL8,KC,MCP-1,MCP1,MIP-1alpha,MIP-1beta,MMP-9,MMP12,MMP14,MMP9,MT1-MMP,NEU1,NF-kappaB,NF-kappaB_DNA,PAI-1,PPCA,SIKVAV,SIKVAV :SIKVAV receptor,SIKVAV receptor,TNF-alpha,TNFR-II,VGVAPG,XGXXPG,XGXXPG: EBP,XGXXPG: EBP: PPCA,XGXXPG: EBP: PPCA: NEU1,fibronectin fragmant,fibronectin fragmant : fibronectin fragment recptor,fibronectin fragment recptor,hyaluronan fragment,hyaluronan fragment: CD44,uPA
C-type lectins play important roles in both innate and adaptive immuneresponses. In contrast to the mannose- or fucose-specific C-type lectins DC-SIGNand mannose receptor, the galactose-type lectins, of which only macrophagegalactose-type lectin (MGL) is found within the immune system, are less wellknown. MGL is selectively expressed by immature dendritic cells and macrophageswith elevated levels on tolerogenic or alternatively activated subsets. HumanMGL has an exclusive specificity for rare terminal GalNAc structures, which arerevealed on the tumor-associated mucin MUC1 and CD45 on effector T cells. Thesefindings implicate MGL in the homeostatic control of adaptive immunity. Wediscuss here the functional similarities and differences between MGL orthologsand compare MGL to its closest homolog, the liver-specific asialoglycoproteinreceptor (ASGP-R).
{kind=link}
{kind=link}
ASGP-R,ASGP-R:Tetra-antennary glycans,ASGP-R:Tri-antennary glycans,CD45AB,CD45ABC,CD45B,CD45B:MGL,CD45BC,Ca2+,DC-SIGN,DC-SIGN:Fucose,DC-SIGN:Mannose,Fucose,GD2,GM1,Galactose,Glycans,Glycoproteins,Glycosphingolipids,MGL,MGL:CD45AB,MGL:CD45ABC,MGL:CD45BC,MGL:GD2,MGL:GM1,MGL:Glycans,MGL:Glycoproteins,MGL:Glycosphingolipids,MGL:MUC1,MR,MUC1,Mannose,Mannose:MR,Tri-antennary glycans,mMGL,mMGL:sialoadhesin,rMGL,rMGL:Galactose,sialoadhesin,tetra-antennary glycans
Innate immunity and inflammation plays a key role in host defense and woundhealing. However, Excessive or altered inflammatory processes can contribute tosevere and diverse human diseases including cardiovascular disease, diabetes andcancer. The interleukin-1 receptor-associated kinases (IRAKs) are criticallyinvolved in the regulation of intracellular signaling networks controllinginflammation. Collective studies indicate that IRAKs are present in many celltypes, and can mediate signals from various cell receptors includingtoll-like-receptors (TLRs). Consequently, diverse downstream signaling processescan be elicited following the activation of various IRAKs. Given the criticaland complex roles IRAK proteins play, it is not surprising that geneticvariations in human IRAK genes have been found to be linked with various humaninflammatory diseases. This review intends to summarize the recent advancesregarding the regulations of various IRAK proteins and their cellular functionsin mediating inflammatory signaling processes.
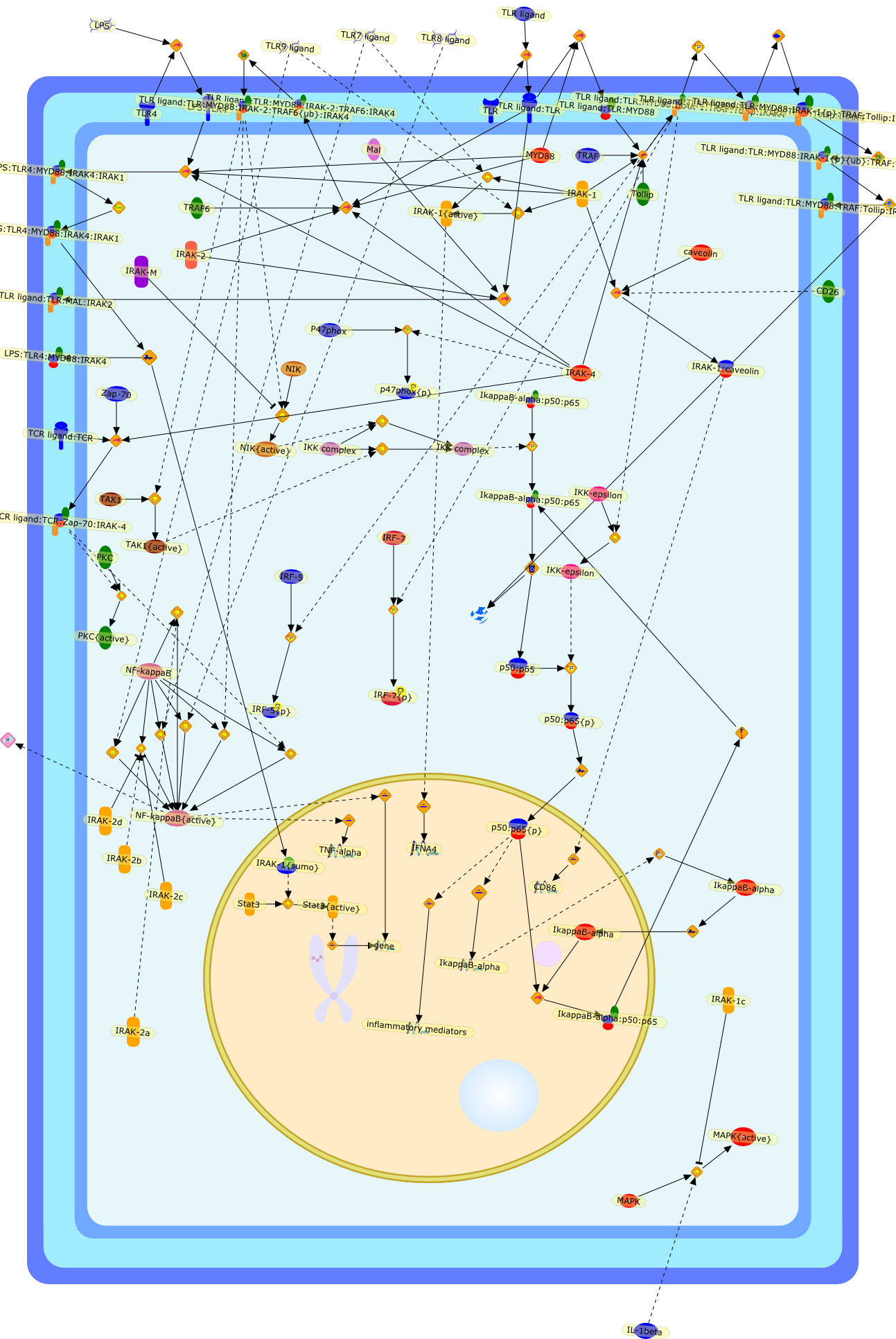{kind=link}
{kind=link}
CD26,CD86,IFNA4,IKK complex,IKK-epsilon,IL-1beta,IRAK-1,IRAK-1:caveolin,IRAK-1c,IRAK-1{active},IRAK-1{sumo},IRAK-2,IRAK-2a,IRAK-2b,IRAK-2c,IRAK-2d,IRAK-4,IRAK-M,IRF-5,IRF-5{p},IRF-7,IRF-7{p},IkappaB-alpha,IkappaB-alpha:p50:p65,LPS,LPS:TLR4,LPS:TLR4:MYD88:IRAK4,LPS:TLR4:MYD88:IRAK4:IRAK1,MAPK,MAPK{active},MYD88,Mal,NF-kappaB,NF-kappaB{active},NIK,NIK{active},P47phox,PKC,PKC{active},Stat3,Stat3{active},TAK1,TAK1{active},TCR ligand:TCR,TCR ligand:TCR:Zap-70:IRAK-4,TLR,TLR ligand,TLR ligand:TLR,TLR ligand:TLR:MAL:IRAK2,TLR ligand:TLR:MYD88,TLR ligand:TLR:MYD88:IRAK-1:TRAF:Tollip:IRAK4,TLR ligand:TLR:MYD88:IRAK-1{p}:TRAF:Tollip:IRAK4,TLR ligand:TLR:MYD88:IRAK-1{p}{ub}:TRAF:Tollip:IRAK4,TLR ligand:TLR:MYD88:IRAK-2:TRAF6:IRAK4,TLR ligand:TLR:MYD88:IRAK-2:TRAF6{ub}:IRAK4,TLR ligand:TLR:MYD88:TRAF:Tollip:IRAK4,TLR4,TLR7 ligand,TLR8 ligand,TLR9 ligand,TNF-alpha,TRAF,TRAF6,Tollip,Zap-70,caveolin,gene,inflammatory mediators,p47phox{p},p50:p65,p50:p65{p},protein remnants
Macrophages exist in almost all animals. In some invertebrates, mesenchymalcells, endothelial cells or fibroblast-like cells can transform intomacrophages. In vertebrates, primitive macrophages first develop in yolk sachematopoiesis and differentiate into fetal macrophages. Monocytes aredifferentiated from hematopoietic stem cells in the late stage of fetalhematopoietic organs and bone marrow. Macrophages serve as an effector inmetabolism and host defense. Depletion of macrophages severely reduced bilirubinproduction and host resistance to infection. Macrophage scavenger receptors areinvolved in host defense. Macrophage growth factors are critical for macrophagedifferentiation and function. In macrophage colony-stimulating factor(M-CSF)-deficient osteopetrotic mice, monocytes, tissue macrophages andosteoclasts are deficient. Granulocyte-macrophage colony-stimulating factor(GM-CSF)-deficient mice develop alveolar proteinosis due to impaired surfactantcatabolism by alveolar macrophages. Accumulation of glucocerebroside inmacrophages due to the deficiency of glucocerebrosidase in lysosomes producesGaucher cells. Macrophages in the arterial wall incorporate chemically modifiedlow-density lipoprotein (LDL) and transform into foam cells. Binding oxidizedLDL to liver X receptor alpha (LXRalpha) upregulates the expression of itstarget genes, which act as cholesterol removers from macrophages. Inflammatorysignals downregulate the expression of LXRalpha and enhance lipid accumulation.Thus, macrophages play a pivotal role in metabolism and host defense.
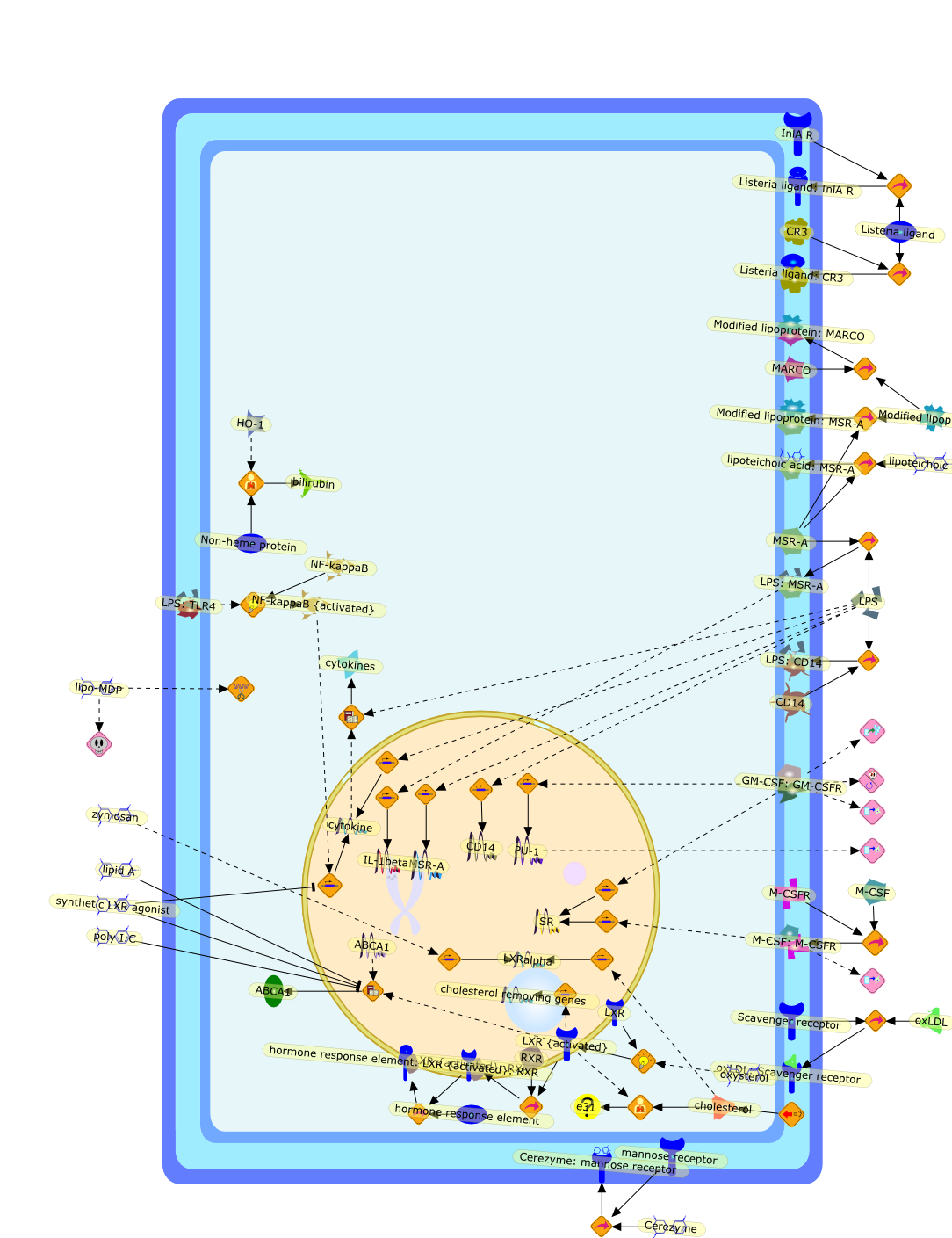{kind=link}
{kind=link}
ABCA1,CD14,CR3,Cerezyme,Cerezyme: mannose receptor,GM-CSF: GM-CSFR,HO-1,IL-1beta,InlA R,LPS,LPS: CD14,LPS: MSR-A,LPS: TLR4,LXR,LXR {activated},LXR {activated}: RXR,LXRalpha,Listeria ligand,Listeria ligand: CR3,Listeria ligand: InlA R,M-CSF,M-CSF: M-CSFR,M-CSFR,MARCO,MSR-A,MSR-A ,Modified lipoprotein,Modified lipoprotein: MARCO,Modified lipoprotein: MSR-A,NF-kappaB,NF-kappaB {activated},Non-heme protein,PU-1,RXR,SR,Scavenger receptor,bilirubin,cholesterol,cholesterol removing genes,cytokine,cytokines,e31,hormone response element,hormone response element: LXR {activated}: RXR,lipid A,lipo-MDP,lipoteichoic acid,lipoteichoic acid: MSR-A,mannose receptor,oxLDL,oxLDL: Scavenger receptor,oxysterol,poly I:C,synthetic LXR agonist,zymosan
Unmethylated DNA with CpG-motifs is recognized by Toll-like receptor 9 (TLR9)and pleiotropic immune responses are elicited. Macrophages and conventionaldendritic cells (cDCs) produce proinflammatory cytokines to B/K-type CpG-DNA,whereas plasmacytoid DCs induce type I interferons to A/D-type CpG-DNA and DNAviruses. The TLR9 mediated signaling pathway is not only responsible foractivation of innate immune cells, but also for mounting acquired responses.Thus, it has been attempted to exploit TLR9 ligands as a vaccine adjuvant foranti-cancer immunotherapy. Further, TLR9 mediated signaling is implicated in thepathogenesis of autoimmune diseases such as systemic lupus erythematosus.Nevertheless, recent studies revealed that double-stranded DNA can be recognizedby intracellular receptor(s) in a TLR9-independent manner. This review willfocus on the roles of TLR9 in immune responses, and its signaling pathways.
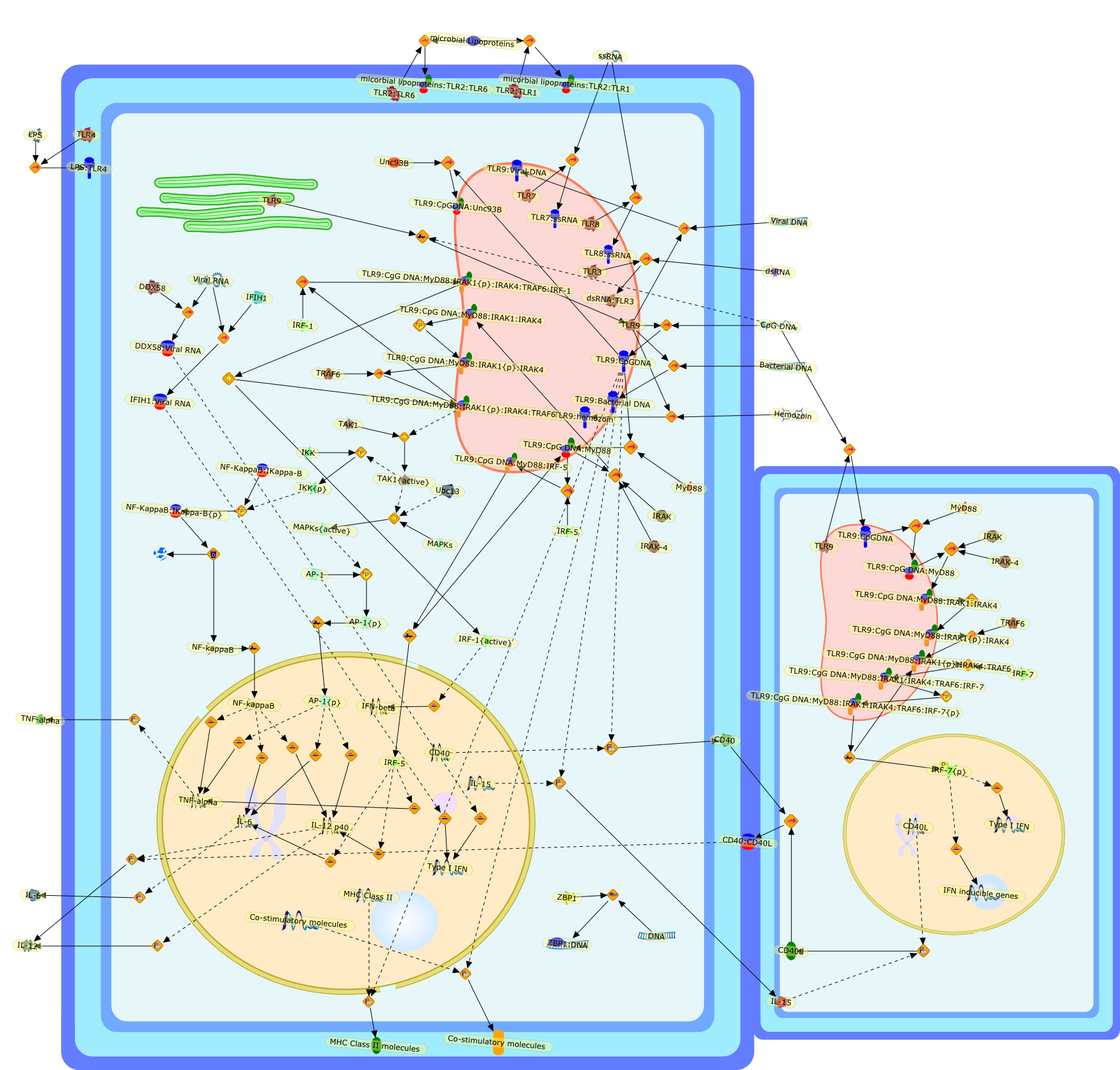{kind=link}
{kind=link}
AP-1,AP-1{p},Bacterial DNA,CD40,CD40:CD40L,CD40L,Co-stimulatory molecules,CpG DNA,DDX58,DDX58:Viral RNA,DNA,Hemozoin,IFIH1,IFIH1:Viral RNA,IFN inducible genes,IFN-beta,IKK,IKK{p},IL-12,IL-12 p40,IL-15,IL-6,IRAK,IRAK-4,IRF-1,IRF-1{active},IRF-5,IRF-7,IRF-7{p},LPS,LPS:TLR4,MAPKs,MAPKs{active},MHC Class II,MHC Class II molecules,MyD88,NF-KappaB:IKappa-B,NF-KappaB:IKappa-B{p},NF-kappaB,TAK1,TAK1{active},TLR2:TLR1,TLR2:TLR6,TLR3,TLR4,TLR7,TLR7:ssRNA,TLR8,TLR8:ssRNA,TLR9,TLR9:Bacterial DNA,TLR9:CgG DNA:MyD88:IRAK1:IRAK4:TRAF6:IRF-7,TLR9:CgG DNA:MyD88:IRAK1:IRAK4:TRAF6:IRF-7{p},TLR9:CgG DNA:MyD88:IRAK1{p}:IRAK4,TLR9:CgG DNA:MyD88:IRAK1{p}:IRAK4:TRAF6,TLR9:CgG DNA:MyD88:IRAK1{p}:IRAK4:TRAF6:IRF-1,TLR9:CpG DNA:MyD88,TLR9:CpG DNA:MyD88:IRAK1:IRAK4,TLR9:CpG DNA:MyD88:IRF-5,TLR9:CpGDNA,TLR9:CpGDNA:Unc93B,TLR9:Viral DNA,TLR9:hemozoin,TNF-alpha,TRAF6,Type I IFN,Ubc13,Unc93B,Viral DNA,Viral RNA,ZBP1,ZBP1:DNA,degradants,dsRNA,dsRNA:TLR3,micorbial lipoproteins:TLR2:TLR1,micorbial lipoproteins:TLR2:TLR6,microbial Lipoproteins,ssRNA
Toll-like receptor 3 (TLR3) recognizes viral double-stranded RNA and itssynthetic analog polyriboinosinic:polyribocytidylic acid (poly(I:C)) and inducestype I interferon (IFN), inflammatory cytokine/chemokine production anddendritic cell (DC) maturation via the adaptor protein TICAM-1 (also calledTRIF). TLR3 is expressed both intracellularly and on the cell surface offibroblasts and epithelial cells, but is localized to the endosomal compartmentof myeloid DCs. Several studies in TLR3-deficient mice demonstrate that TLR3participates in the generation of protective immunity against some viralinfections. Involvement of TLR3-TICAM-1 in activation of NK cells and CTLs bymyeloid DCs suggests that TLR3 serves as an inducer of cellular immunity sensingviral infection rather than a simple IFN inducer. In this review, we summarizethe current knowledge on TLR3 and discuss its possible role in innate andadaptive immunity.
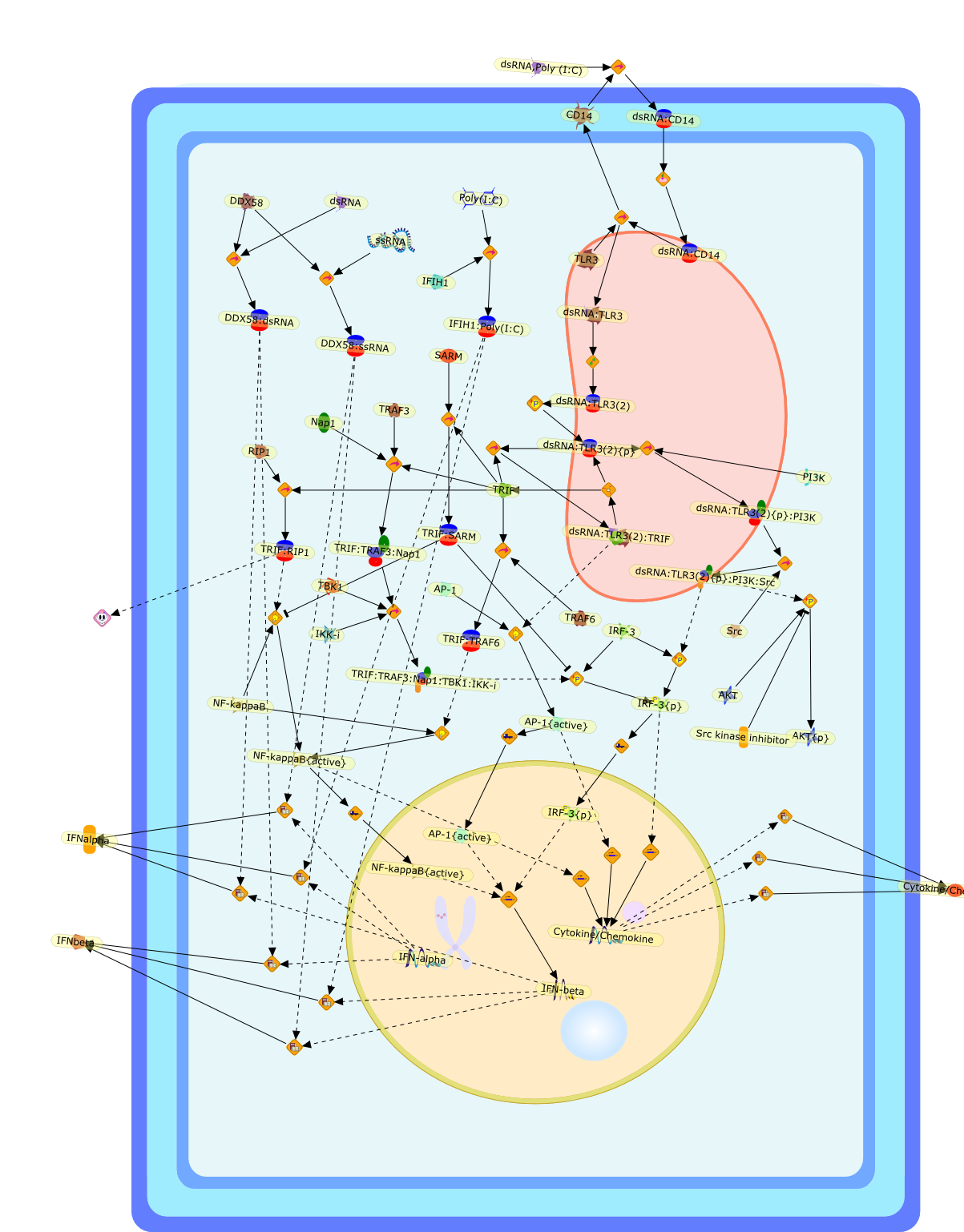{kind=link}
{kind=link}
AKT,AKT{p},AP-1,AP-1{active},CD14,Cytokine/Chemokine,DDX58,DDX58:dsRNA,DDX58:ssRNA,IFIH1,IFIH1:Poly(I:C),IFN-alpha,IFN-beta,IFNalpha,IFNbeta,IKK-i,IRF-3,IRF-3{p},NF-kappaB,NF-kappaB{active},Nap1,PI3K,Poly(I:C),RIP1,SARM,Src,Src kinase inhibitor,TBK1,TLR3,TRAF3,TRAF6,TRIF,TRIF:RIP1,TRIF:SARM,TRIF:TRAF3:Nap1,TRIF:TRAF3:Nap1:TBK1:IKK-i,TRIF:TRAF6,dsRNA,dsRNA,Poly (I:C),dsRNA:CD14,dsRNA:TLR3,dsRNA:TLR3(2),dsRNA:TLR3(2):TRIF,dsRNA:TLR3(2){p},dsRNA:TLR3(2){p}:PI3K,dsRNA:TLR3(2){p}:PI3K:Src,ssRNA
The transcription factor NF-kappaB has served as a standard for inducibletranscription factors for more than 20 years. The numerous stimuli that activateNF-kappaB, and the large number of genes regulated by NF-kappaB, ensure thatthis transcription factor is still the subject of intense research. Here, weattempt to synthesize some of the basic principles that have emerged fromstudies of NF-kappaB, and we aim to generate a more unified view of NF-kappaBregulation.
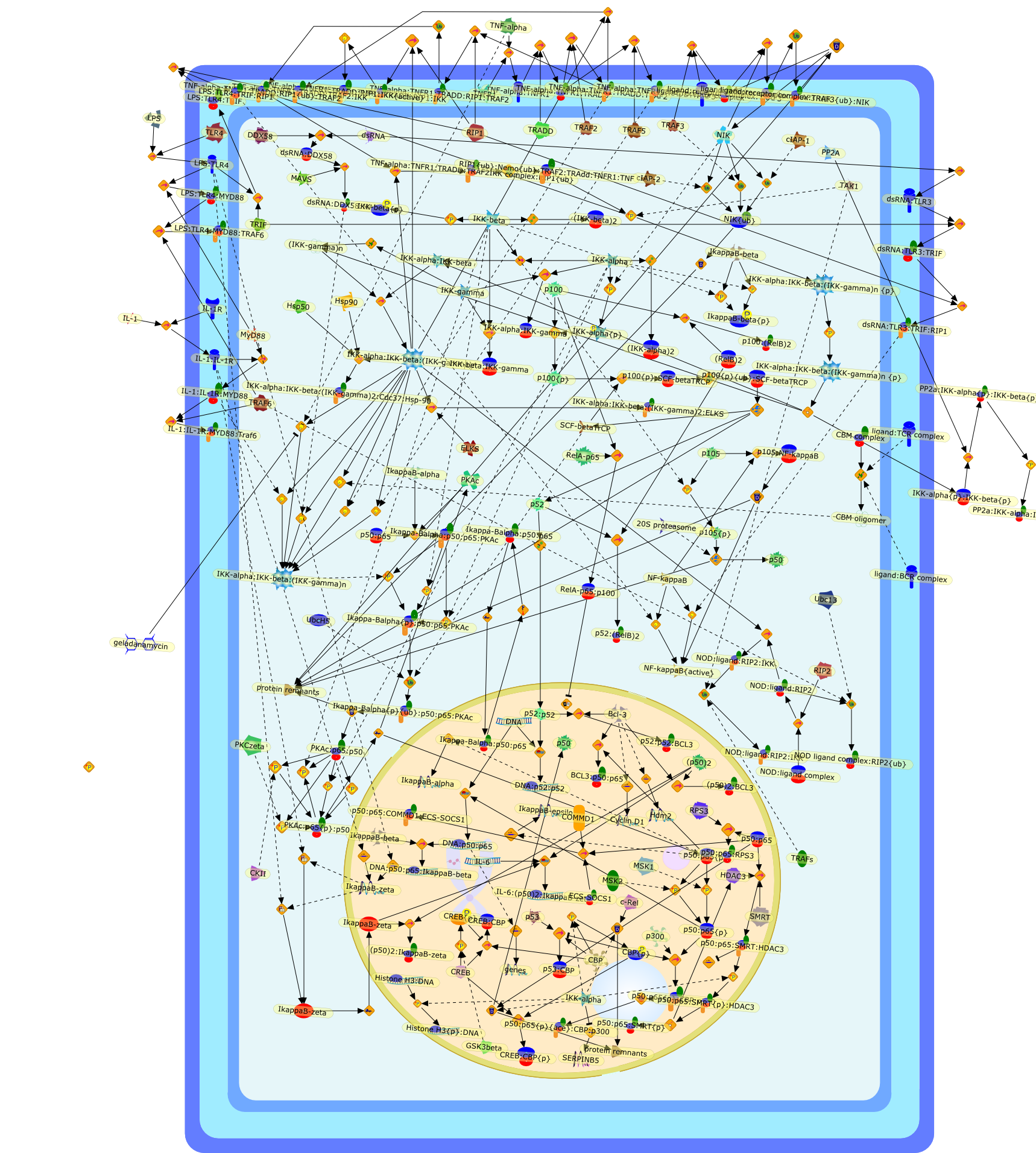{kind=link}
{kind=link}
(IKK-alpha)2,(IKK-beta)2,(IKK-gamma)n,(RelB)2,(p50)2,(p50)2:BCL3,(p50)2:IkappaB-zeta,20S proteasome,BCL3:p50:p65,Bcl-3,CBM complex,CBM oligomer,CBP,CBP{p},CKII,COMMD1,CREB,CREB:CBP,CREB:CBP{p},CREB{p},Cyclin D1,DDX58,DNA,DNA:p50:p65,DNA:p50:p65:IkappaB-beta,DNA:p52:p52,ECS-SOCS1,ELKS,GSK3beta,HDAC3,Hdm2,Histone H3:DNA,Histone H3{p}:DNA,Hsp50,Hsp90,IKK-alpha,IKK-alpha:IKK-beta,IKK-alpha:IKK-beta:(IKK-gamma)2:Cdc37:Hsp-90,IKK-alpha:IKK-beta:(IKK-gamma)2:ELKS,IKK-alpha:IKK-beta:(IKK-gamma)n,IKK-alpha:IKK-beta:(IKK-gamma)n {p},IKK-alpha:IKK-gamma,IKK-alpha{p},IKK-alpha{p}:IKK-beta{p},IKK-beta,IKK-beta:IKK-gamma,IKK-beta{p},IKK-gamma,IL-1,IL-1:IL-1R,IL-1:IL-1R:MYD88,IL-1:IL-1R:MYD88:Traf6,IL-1R,IL-6,IL-6:(p50)2:IkappaB-zeta,Ikappa-Balpha:p50:p65,Ikappa-Balpha:p50:p65:PKAc,Ikappa-Balpha{p}:p50:p65:PKAc,Ikappa-Balpha{p}{ub}:p50:p65:PKAc,IkappaB-alpha,IkappaB-beta,IkappaB-beta{p},IkappaB-epsilon,IkappaB-zeta,LPS,LPS:TLR4,LPS:TLR4:MYD88,LPS:TLR4:MYD88:TRAF6,LPS:TLR4:TRIF,LPS:TLR4:TRIF:RIP1,MAVS,MSK1,MSK2,MyD88,NF-kappaB,NF-kappaB{active},NIK,NIK{ub},NOD ligand complex:RIP2{ub},NOD:ligand complex,NOD:ligand:RIP2,NOD:ligand:RIP2:IKK,PKAc,PKAc:p65:p50,PKAc:p65{p}:p50,PKCzeta,PP2A,PP2a:IKK-alpha:IKK-beta,PP2a:IKK-alpha{p}:IKK-beta{p},RIP1,RIP1{ub}:Nemo{ub}:TRAF2:TRAdd:TNFR1:TNF,RIP2,RPS3,RelA-p65,RelA-p65:p100,SCF-betaTrCP,SERPINB5,SMRT,TAK1,TLR4,TNF-alpha,TNF-alpha:TNFR1,TNF-alpha:TNFR1:TRADD,TNF-alpha:TNFR1:TRADD:RIP1:IKK,TNF-alpha:TNFR1:TRADD:RIP1:IKK{active},TNF-alpha:TNFR1:TRADD:RIP1:TRAF2,TNF-alpha:TNFR1:TRADD:RIP1{ub}:TRAF2,TNF-alpha:TNFR1:TRADD:TRAF2,TNF-alpha:TNFR1:TRADD:TRAF2:IKK,TNF-alpha:TNFR1:TRADD:TRAF2IKK complex:RIP1{ub},TNF-alpha:TNFR1:TRADD:TRAF5,TNFR1,TRADD,TRAF2,TRAF3,TRAF5,TRAF6,TRAFs,TRIF,Ubc13,UbcH5,c-Rel,cIAP-1,cIAP-2,dsRNA,dsRNA:DDX58,dsRNA:DDX58:MAVS,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:RIP1,geladanamycin,genes,ligand:BCR complex,ligand:TCR complex,ligand:receptor complex,ligand:receptor complex:TRAF3,ligand:receptor complex:TRAF3:NIK,ligand:receptor complex:TRAF3{ub}:NIK,p100,p100:(RelB)2,p100{p},p100{p}:SCF-betaTRCP,p100{p}{ub}:SCF-betaTRCP,p105,p105:NF-kappaB,p105{p},p300,p50,p50:p65,p50:p65:COMMD1:ECS-SOCS1,p50:p65:RPS3,p50:p65:SMRT:HDAC3,p50:p65:SMRT{p},p50:p65:SMRT{p}:HDAC3,p50:p65{p},p50:p65{p}:CBP:p300,p50:p65{p}{ace}:CBP:p300,p52,p52:(RelB)2,p52:p52,p52:p52:BCL3,p53,p53:CBP,protein remnants
The innate immune system initially recognizes RNA virus infection and evokesantiviral responses by producing type I interferons (IFNs). Toll-like receptors(TLRs) and cytoplasmic retinoic acid-inducible gene I (RIG-I)-like helicases(RLHs) are the two major receptor systems for detecting RNA viruses. The RLHsignaling pathways play essential roles in the recognition of RNA viruses invarious cells, with the exception of plasmacytoid dendritic cells, which utilizeTLRs for virus recognition. The route of infection determines the cell typesresponsible for type I IFN production. Recent studies have suggested that TLRsare critical for activation of adaptive immune responses against several virusinfections, although it may be premature to draw such a conclusion for virusinfections in general. In this review, we will discuss recent advances towardclarifying the signaling pathways activated by RLHs and TLRs.
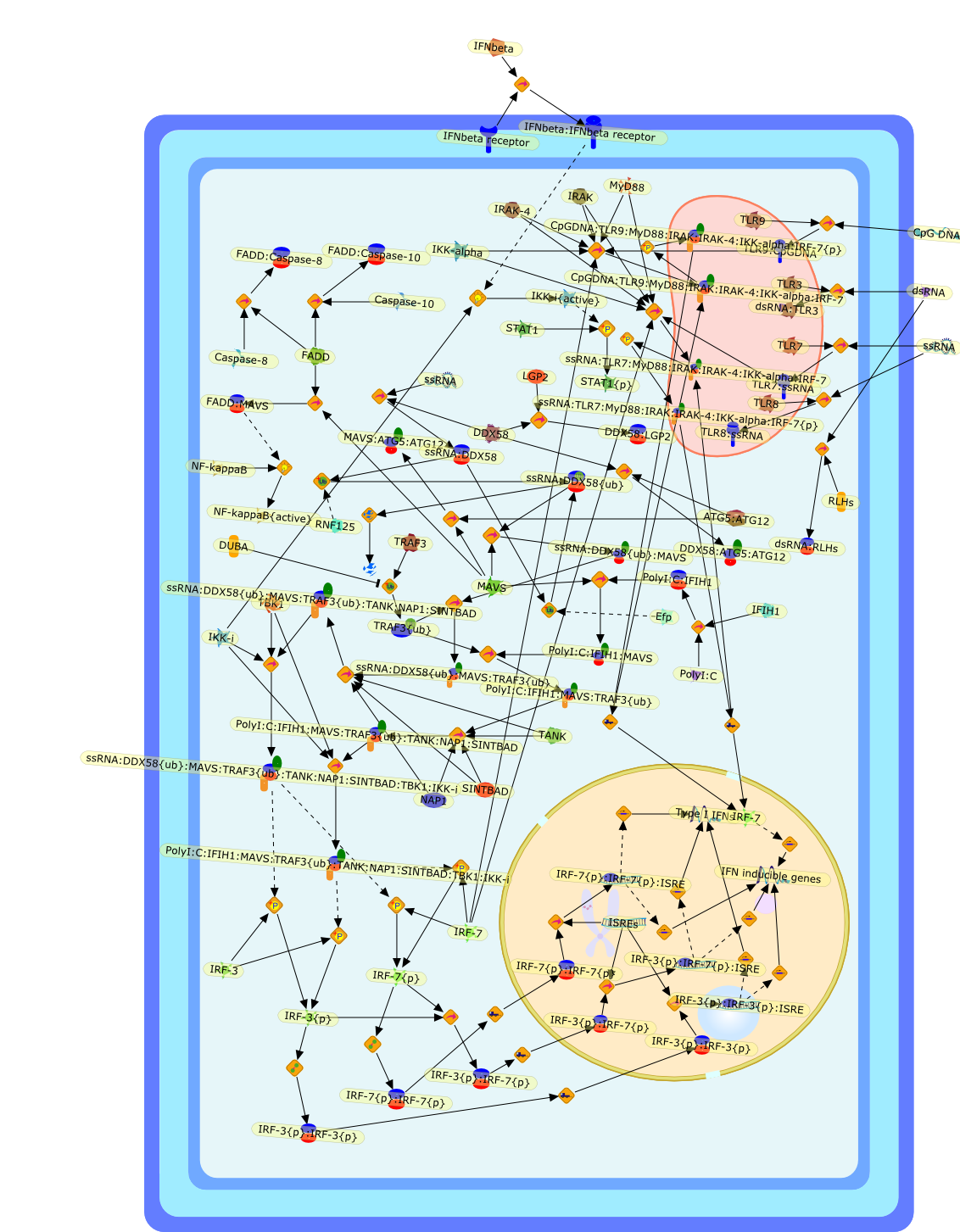{kind=link}
{kind=link}
ATG5:ATG12,Caspase-10,Caspase-8,CpG DNA,CpGDNA:TLR9:MyD88:IRAK:IRAK-4:IKK-alpha:IRF-7,CpGDNA:TLR9:MyD88:IRAK:IRAK-4:IKK-alpha:IRF-7{p},DDX58,DDX58:ATG5:ATG12,DDX58:LGP2,DUBA,Efp,FADD,FADD:Caspase-10,FADD:Caspase-8,FADD:MAVS,IFIH1,IFN inducible genes,IFNbeta,IFNbeta receptor,IFNbeta:IFNbeta receptor,IKK-alpha,IKK-i,IKK-i{active},IRAK,IRAK-4,IRF-3,IRF-3{p},IRF-3{p}:IRF-3{p},IRF-3{p}:IRF-3{p}:ISRE,IRF-3{p}:IRF-7{p},IRF-3{p}:IRF-7{p}:ISRE,IRF-7,IRF-7{p},IRF-7{p}:IRF-7{p},IRF-7{p}:IRF-7{p}:ISRE,ISREs,LGP2,MAVS,MAVS:ATG5:ATG12,MyD88,NAP1,NF-kappaB,NF-kappaB{active},PolyI:C,PolyI:C:IFIH1,PolyI:C:IFIH1:MAVS,PolyI:C:IFIH1:MAVS:TRAF3{ub},PolyI:C:IFIH1:MAVS:TRAF3{ub}:TANK:NAP1:SINTBAD,PolyI:C:IFIH1:MAVS:TRAF3{ub}:TANK:NAP1:SINTBAD:TBK1:IKK-i,RLHs,RNF125,SINTBAD,STAT1,STAT1{p},TANK,TBK1,TLR3,TLR7,TLR7:ssRNA,TLR8,TLR8:ssRNA,TLR9,TLR9:CpGDNA,TRAF3,TRAF3{ub},Type I IFNs,degradants,dsRNA,dsRNA:RLHs,dsRNA:TLR3,ssRNA,ssRNA:DDX58,ssRNA:DDX58{ub},ssRNA:DDX58{ub}:MAVS,ssRNA:DDX58{ub}:MAVS:TRAF3{ub},ssRNA:DDX58{ub}:MAVS:TRAF3{ub}:TANK:NAP1:SINTBAD,ssRNA:DDX58{ub}:MAVS:TRAF3{ub}:TANK:NAP1:SINTBAD:TBK1:IKK-i,ssRNA:TLR7:MyD88:IRAK:IRAK-4:IKK-alpha:IRF-7,ssRNA:TLR7:MyD88:IRAK:IRAK-4:IKK-alpha:IRF-7{p}
In recent years, it has become increasingly evident that mammalian Toll-likereceptors (TLRs) play a critical role in determining the outcome of virusinfection. TLRs have evolved to recognize viral nucleic acids, and promote thestimulation of innate and adaptive immune responses. Interestingly, the study ofmice harboring deficiencies in various TLR proteins and their adaptors suggeststhat TLR activation promotes protective anti-viral immunity in some cases, whileexacerbating virus-induced disease in others. In this report we describe theinteractions of viruses with both the TLR system and the intracellularrecognition system and highlight the role of TLRs in shaping the outcome ofvirus infection in both a positive and negative manner.
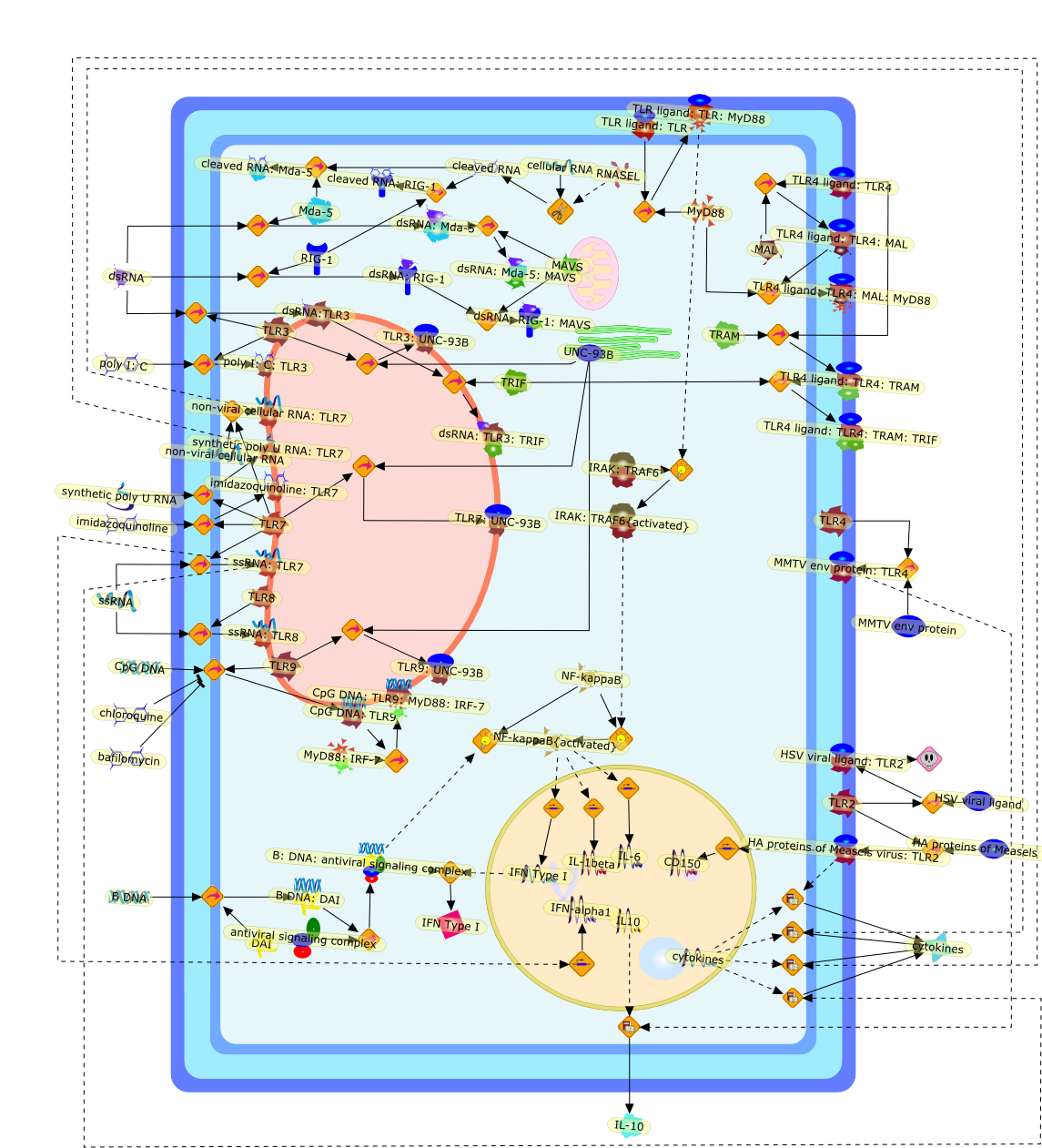{kind=link}
{kind=link}
B DNA,B DNA: DAI,B: DNA: antiviral signaling complex,CD150,CpG DNA,CpG DNA: TLR9,CpG DNA: TLR9: MyD88: IRF-7,DAI,HA proteins of Measels virus,HA proteins of Measels virus: TLR2,HSV viral ligand,HSV viral ligand: TLR2,IFN Type I,IFN-alpha1,IL-10,IL-1beta,IL-6,IL10,IRAK: TRAF6,IRAK: TRAF6{activated},MAL,MAVS,MMTV env protein,MMTV env protein: TLR4,Mda-5,MyD88,MyD88: IRF-7,NF-kappaB,NF-kappaB{activated},RIG-1,RNASEL,TLR ligand: TLR,TLR ligand: TLR: MyD88,TLR2,TLR3,TLR3: UNC-93B,TLR4,TLR4 ligand: TLR4,TLR4 ligand: TLR4: MAL,TLR4 ligand: TLR4: MAL: MyD88,TLR4 ligand: TLR4: TRAM,TLR4 ligand: TLR4: TRAM: TRIF,TLR7,TLR7: UNC-93B,TLR8,TLR9,TLR9: UNC-93B,TRAM,TRIF,UNC-93B,antiviral signaling complex,bafilomycin,cellular RNA,chloroquine,cleaved RNA,cleaved RNA: Mda-5,cleaved RNA: RIG-1,cytokines,dsRNA,dsRNA: Mda-5,dsRNA: Mda-5: MAVS,dsRNA: RIG-1,dsRNA: RIG-1: MAVS,dsRNA: TLR3: TRIF,dsRNA:TLR3,imidazoquinoline,imidazoquinoline: TLR7,non-viral cellular RNA,non-viral cellular RNA: TLR7,poly I: C,poly I: C: TLR3,ssRNA,ssRNA: TLR7,ssRNA: TLR8,synthetic poly U RNA,synthetic poly U RNA: TLR7
The detection of microbial components by pattern recognition receptors (PRRs)and the subsequent triggering of innate immune responses constitute the firstline of defense against infections. Recently, much attention has been focused oncytosolic nucleic acid receptors; the activation of these receptors commonlyevokes a robust innate immune response, the hallmark of which is the inductionof type I interferon (IFN) genes. In addition to receptors for RNA, receptorsthat detect DNA exposed in the cytosol and activate innate immune responses havelong been thought to exist. Recently, DAI (DLM-1/ZBP1) has been identified as acandidate cytosolic DNA sensor. Cytosolic signaling by DNA-activated DAI(DLM-1/ZBP1) signaling results in activation of the two pathways of genetranscription critical to innate immune responses, the IRF and NF-kappaBpathways. In this review, we summarize our current view of activation mechanismand immunological roles of DAI (DLM-1/ZBP1) and related molecules. In addition,we also discuss the issue of self vs. non-self DNA recognition by DAI(DLM-1/ZBP1) and other DNA sensors in terms of the possible involvement inautoimmune abnormalities.
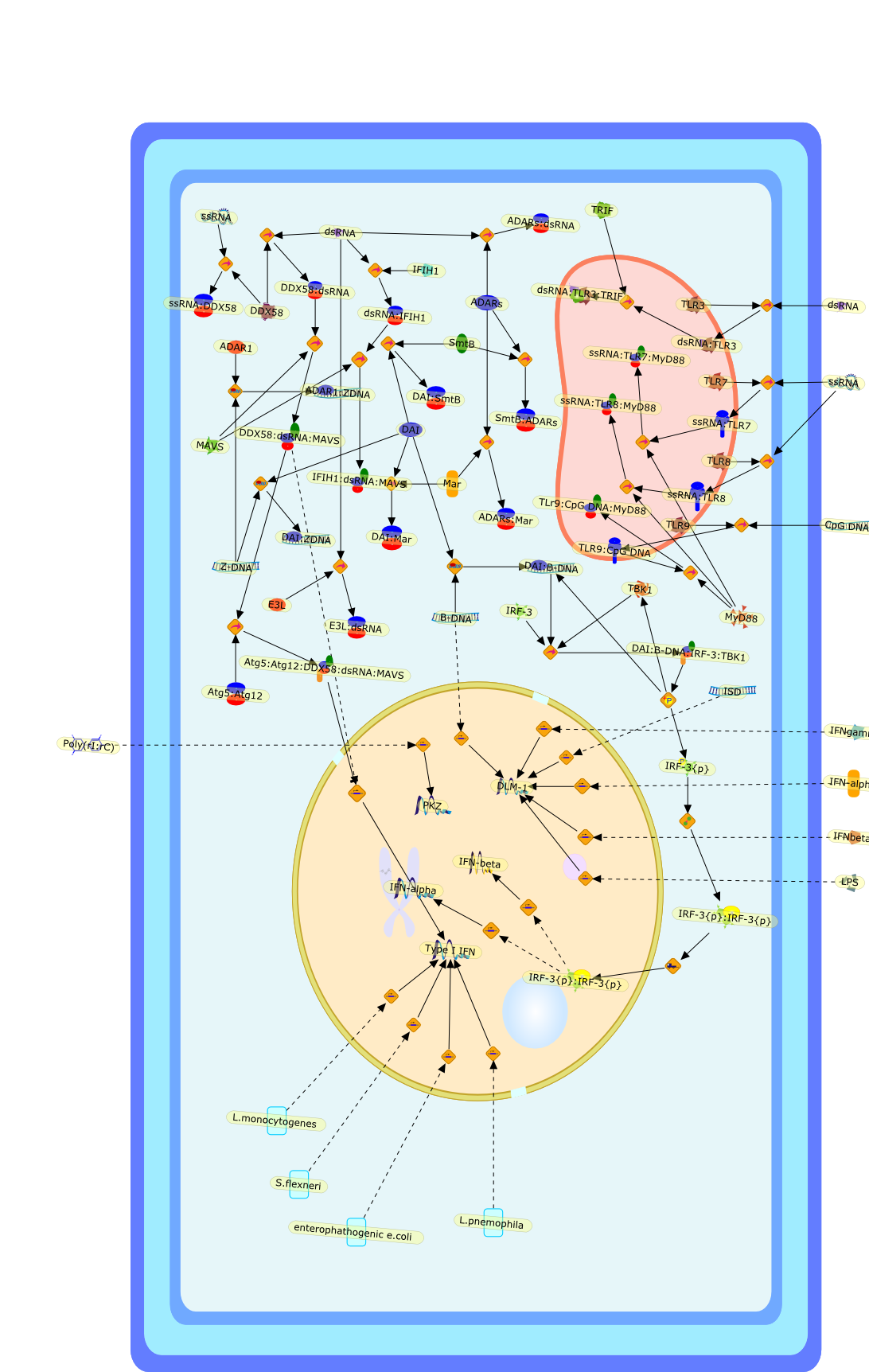{kind=link}
{kind=link}
ADAR1,ADAR1:ZDNA,ADARs,ADARs:Mar,ADARs:dsRNA,Atg5:Atg12,Atg5:Atg12:DDX58:dsRNA:MAVS,B-DNA,CpG DNA,DAI,DAI:B-DNA,DAI:B-DNA:IRF-3:TBK1,DAI:Mar,DAI:SmtB,DAI:ZDNA,DDX58,DDX58:dsRNA,DDX58:dsRNA:MAVS,DLM-1,E3L,E3L:dsRNA,IFIH1,IFIH1:dsRNA:MAVS,IFN-alpha,IFN-beta,IFNbeta,IFNgamma,IRF-3,IRF-3{p},IRF-3{p}:IRF-3{p},ISD,L.monocytogenes,L.pnemophila,LPS,MAVS,Mar,MyD88,PKZ,Poly(rI:rC),S.flexneri,SmtB,SmtB:ADARs,TBK1,TLR3,TLR7,TLR8,TLR9,TLR9:CpG DNA,TLr9:CpG DNA:MyD88,TRIF,Type I IFN,Z-DNA,dsRNA,dsRNA:IFIH1,dsRNA:TLR3,dsRNA:TLR3:TRIF,enterophathogenic e.coli,ssRNA,ssRNA:DDX58,ssRNA:TLR7,ssRNA:TLR7:MyD88,ssRNA:TLR8,ssRNA:TLR8:MyD88
The stimulation of Toll-like receptor 4 (TLR4) by lipopolysaccharide (LPS)induces the release of critical proinflammatory cytokines that are necessary toactivate potent immune responses. LPS/TLR4 signaling has been intensivelystudied in the past few years. Here we review molecules involved inTLR4-mediated signaling, including players that are involved in the negativeregulation of this important pathway.
{kind=link}
{kind=link}
AP-1,AP-1{active},CD14,CpGDNA,Envelope protein,F protein,IKK,IKK-alpha,IKK-beta,IKK-gamma,IKK{active},IRAK-4,IRAK-M,IRAK1,IRAK1 degradants,IRF-3,IRF-3{active},IRF3:IRF3,Ikki,LBP,LPS,LPS:LBP,LPS:LBP:CD14,LPS:TLR4:TRAM,LPS:TLR4:TRAM:TRIF,LPS:TLR4:TRAM:TRIF:RIP1,LPS:TLR4:TRAM:TRIF:TRAF3,LPS:TLR4:TRAM:TRIF:TRAF3:TANK:IKKi:TBK1,LTA,MAPKs,MAPKs{active},MD-1,MD-2,MyD88,NF-KappaB:IKappaB,NF-KappaB:IKappaB{p},NF-kappaB,NF-kappaB{active},PGN,RIP1,RP105,RP105:MD-1,SIGIRR,SOCS-1,ST2L,ST2L:MyD88,ST2L:TIRAP,TAK1,TAK1{active},TANK,TBK1,TIRAP,TIRAP{ub},TLR2,TLR2:TLR6,TLR2:TLR6:LTA,TLR2:TLR6:PGN,TLR3,TLR4,TLR4:F protein,TLR4:LPS oligomer,TLR4:LPS oligomer:TIRAP,TLR4:LPS oligomer:TIRAP:MyD88,TLR4:LPS:TIRAP:MyD88:IRAK-4{active],TLR4:LPS:TIRAP:MyD88:IRAK-4{active},TLR4:LPS:TIRAP:MyD88:IRAK-4{active}:IRAK1{active},TLR4:MD-2,TLR4:MD-2:LPS:LBP:CD14,TLR4:MD-2:RP105:MD-1,TLR4:envelope protein,TLR6,TLR7,TLR7:ssRNA,TLR8,TLR8:ssRNA,TLR9,TLR9:CpGDNA,TRAF1,TRAF1cleaved fragments,TRAF3,TRAF4,TRAF4:TRAF6,TRAF6,TRAF6{active},TRAF6{active}:Ubc13:UVE1A,TRAM,TRIAD3A,TRIAD3A:RIP1,TRIAD3A:TIRAP,TRIAD3A:TRIF,TRIF,TRIF:TRAF4,Type I interferons,UVE1A,Ubc13,caspases,degradants,dsRNA,dsRNA:TLR3,proinflammatory cytokines,ssRNA
Nucleic acids from microbes were known to be immunogenic for a long time, buttheir sensors have been only recently identified. Two types of sensors,Toll-like receptors (TLR) and the RIG-I-like receptors (RLR), principallyrespond to nucleic acid. Discovery of these sensors have opened basic researchas to how our immune system handles microbial nucleic acids. Recent resultsdemonstrate that functional interaction between pathogen sensors is criticallyinvolved in the extent and type of immune responses. Moreover, responsiveness ofnucleic acid-sensing TLRs is profoundly influenced by molecules associated withTLRs or with nucleic acids. Our understanding of mechanisms by which TLRresponses were controlled would contribute to development of drugs such asimmunostimulatory nucleic acids or small chemicals antagonizing TLRs.
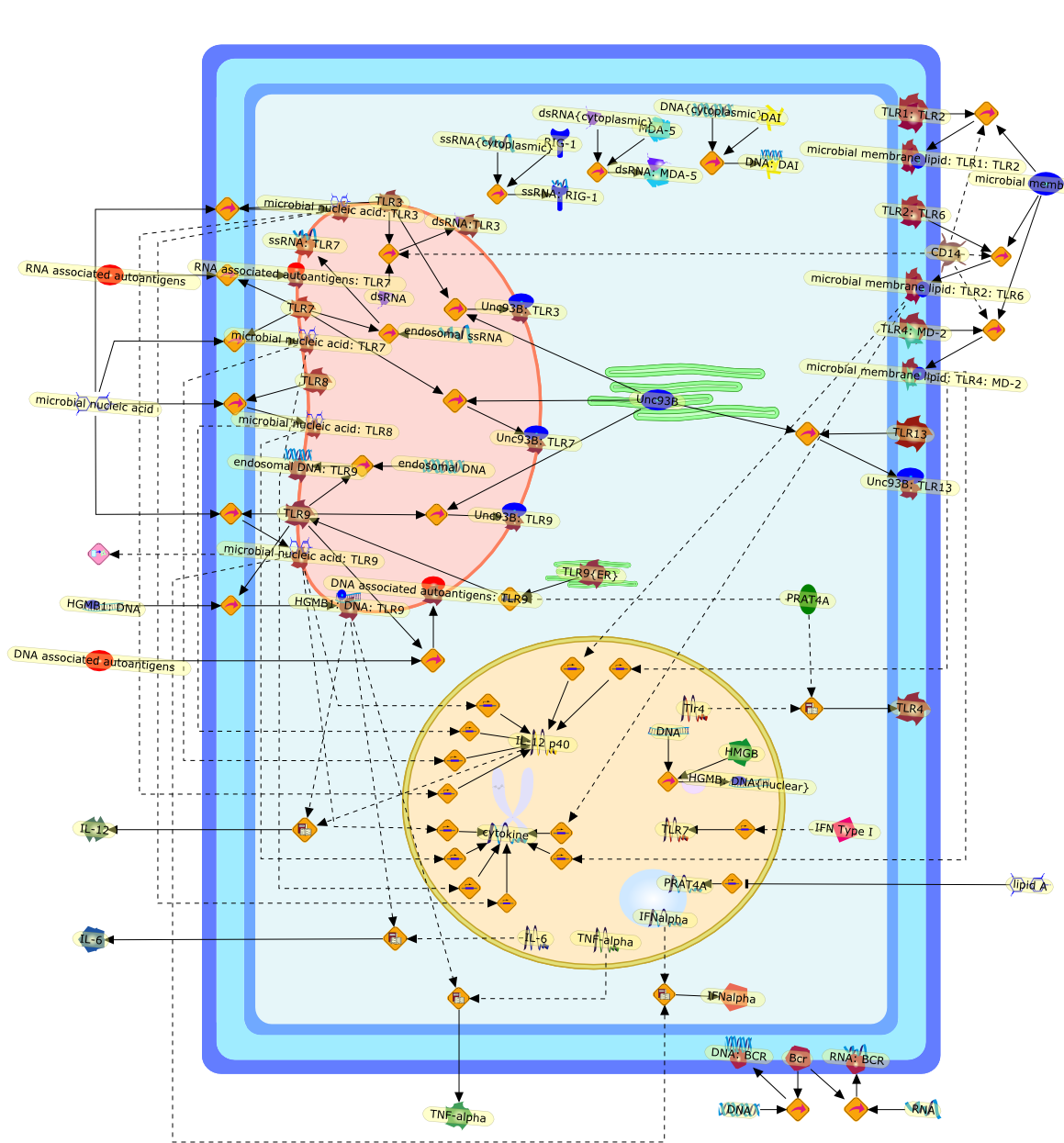{kind=link}
{kind=link}
Bcr,CD14,DAI,DNA,DNA associated autoantigens,DNA associated autoantigens: TLR9,DNA: BCR,DNA: DAI,DNA{cytoplasmic},HGMB1: DNA,HGMB1: DNA: TLR9,HGMB: DNA{nuclear},HMGB,IFN Type I,IFNalpha,IL-12,IL-12 p40,IL-6,MDA-5,PRAT4A,RIG-1,RNA,RNA associated autoantigens,RNA associated autoantigens: TLR7,RNA: BCR,TLR13,TLR1: TLR2,TLR2: TLR6,TLR3,TLR4,TLR4: MD-2,TLR7,TLR8,TLR9,TLR9{ER},TNF-alpha,Tlr4,Unc93B,Unc93B: TLR13,Unc93B: TLR3,Unc93B: TLR7,Unc93B: TLR9,cytokine,dsRNA,dsRNA: MDA-5,dsRNA:TLR3,dsRNA{cytoplasmic},endosomal DNA,endosomal DNA: TLR9,endosomal ssRNA,lipid A,microbial membrane lipid,microbial membrane lipid: TLR1: TLR2,microbial membrane lipid: TLR2: TLR6,microbial membrane lipid: TLR4: MD-2,microbial nucleic acid,microbial nucleic acid: TLR3,microbial nucleic acid: TLR7,microbial nucleic acid: TLR8,microbial nucleic acid: TLR9,ssRNA: RIG-1,ssRNA: TLR7,ssRNA{cytoplasmic}
Chronic and persistent inflammation contributes to cancer development and canpredispose to carcinogenesis. Infection-driven inflammations are involved in thepathogenesis of approximately 15-20% of human tumors. However, even tumors thatare not epidemiologically linked to pathogens are characterized by the presenceof an inflammatory component in their microenvironment. Hallmarks ofcancer-associated inflammation include the presence of infiltrating leukocytes,cytokines, chemokines, growth factors, lipid messengers, and matrix-degradingenzymes. Schematically, two interrelated pathways link inflammation and cancer:(1) genetic events leading to neoplastic transformation promote the constructionof an inflammatory milieu; (2) tumor-infiltrating leukocytes, in particularmacrophages, are prime regulators of cancer inflammation. Thus, an intrinsicpathway of inflammation (driven in tumor cells), as well as an extrinsic pathway(in tumor-infiltrating leukocytes) have been described and both contribute totumor progression.
{kind=link}
{kind=link}
Braf,CCL17,CCL17:CCR4,CCL18,CCL2,CCL20,CCL22,CCL22:CCR4,CCR4,COX-2,CXCR4,Chemokines,Cytokine,EGF,FGF,Hsp60,Hyaluronan,IL-1,IL-10,IL-13,IL-1R:ligand:MyD88,IL-1R:ligand:MyD88:IRAK:TRAF6,IL-1R:ligand:MyD88:SIGIRR,IL-4,IL-6,IL-8,IRAK,L-selectin,MMP2,MMP9,NF-kappaB,NF-kappaB{active},PDGF,ROI,Ras,SIGIRR,TGFbeta,TLR4,TLR4:Hsp60,TLR4:Hyaluronan,TLR4:fibronectin,TLR4:ligand:MyD88,TLR4:ligand:MyD88:IRAK:TRAF6,TLR4:ligand:MyD88:SIGIRR,TNF,TPA,TRAF6,VEGF,cytokines,fibronectin,tPA,uPAR,urokinase plaminogen activator,urokinase plasminogen activator
Adiponectin is an adipokine with potent anti-inflammatory properties. Thedevelopment of alcoholic liver disease is thought to involve increasedpro-inflammatory activity, mediated in part by the activation of hepaticmacrophages (Kupffer cells). Chronic ethanol feeding sensitizes hepaticmacrophages to activation by lipopolysaccharide (LPS), leading to increasedproduction of reactive oxygen species and tumor necrosis factor-alpha(TNF-alpha). Adiponectin can normalize Toll-like receptor-4 (TLR-4) mediatedsignaling in hepatic macrophages after ethanol feeding, likely contributing tothe hepatoprotective effect of adiponectin in the progression of alcoholic liverdisease. However, the mechanisms by which adiponectin suppress TLR-4 mediatedresponses are not well understood. Using the macrophage-like cell line,RAW264.7, we have investigated the molecular mechanisms by which adiponectinsuppresses LPS-stimulated TNF-alpha production. Globular adiponectin(gAcrp)-mediated desensitization of LPS-stimulated responses in RAW264.7macrophages was dependent on the production of the anti-inflammatory cytokineinterleukin (IL)-10. gAcrp initially increased TNF-alpha expression in RAW264.7macrophages; this TNF-alpha then contributed to increased expression of IL-10.This initial gAcrp-mediated increase in TNF-alpha production by macrophages wasmediated via activation of ERK1/2-->Egr-1 and nuclear factor(NF)-kappaB-dependent mechanisms. gAcrp-stimulated IL-10 expression was alsodependent on the phosphorylation of cAMP response element-binding protein andthe cAMP response element in the IL-10 promoter. In summary, these studiesreveal a complex, integrated response of macrophages to gAcrp. gAcrp initiallyactivated signaling pathways considered to be pro-inflammatory, with asubsequent increase in the expression of the potent, anti-inflammatory cytokine,IL-10. Increased IL-10 expression was ultimately required for the suppression ofTLR4-mediated signaling by gAcrp.
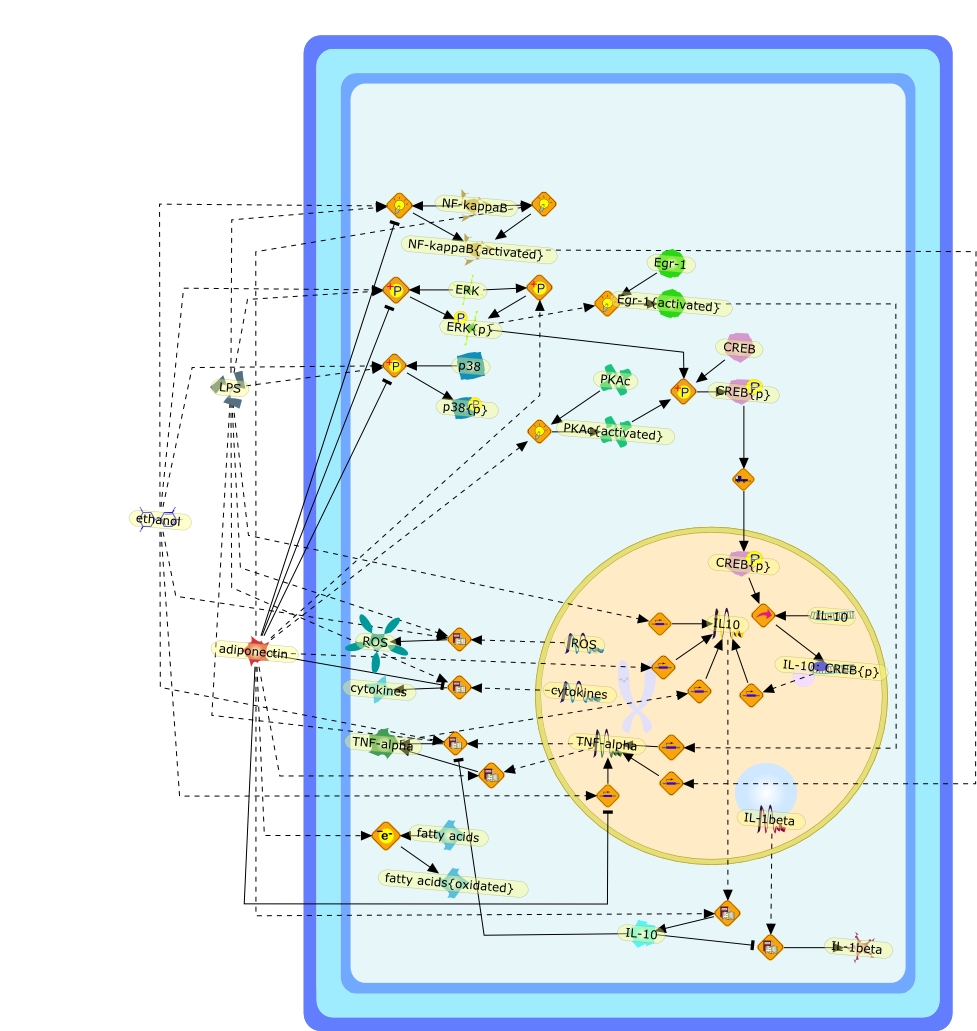{kind=link}
{kind=link}
CREB,CREB{p},ERK,ERK{p},Egr-1,Egr-1{activated},IL-10,IL-10: CREB{p},IL-1beta,IL10,LPS,NF-kappaB,NF-kappaB{activated},PKAc,PKAc{activated},ROS,TNF-alpha,adiponectin,cytokines,ethanol,fatty acids,fatty acids{oxidated},p38,p38{p}
The IkappaB kinases (IKKs) IKK-alpha and IKK-beta, and the IKK-related kinasesTBK1 and IKK-epsilon, have essential roles in innate immunity throughsignal-induced activation of NF-kappaB, IRF3 and IRF7, respectively. Althoughthe signaling events within these pathways have been extensively studied, themechanisms of IKK and IKK-related complex assembly and activation remain poorlydefined. Recent data provide insight into the requirement for scaffold proteinsin complex assembly; NF-kappaB essential modulator coordinates some IKKcomplexes, whereas TANK, NF-kappaB-activating kinase-associated protein 1 (NAP1)or similar to NAP1 TBK1 adaptor (SINTBAD) assemble TBK1 and IKK-epsiloncomplexes. The different scaffold proteins undergo similar post-translationalmodifications, including phosphorylation and non-degradative polyubiquitylation.Moreover, increasing evidence indicates that distinct scaffold proteins assembleIKK, and potentially TBK1 and IKK-epsilon subcomplexes, in a stimulus-specificmanner, which might be a mechanism to achieve specificity.
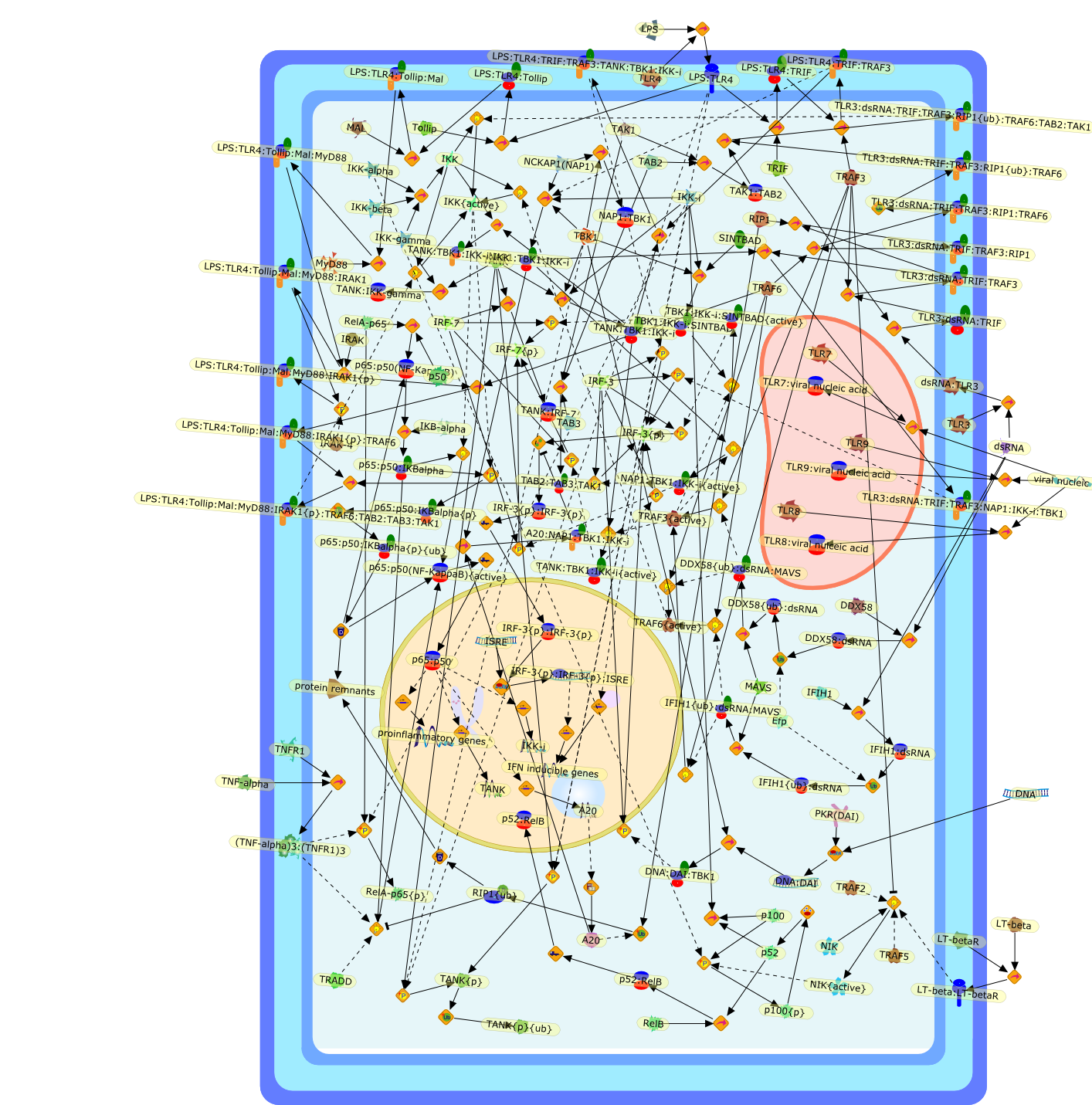{kind=link}
{kind=link}
(TNF-alpha)3:(TNFR1)3,A20,A20:NAP1:TBK1:IKK-i,DDX58,DDX58:dsRNA,DDX58{ub}:dsRNA,DDX58{ub}:dsRNA:MAVS,DNA,DNA:DAI,DNA:DAI:TBK1,Efp,IFIH1,IFIH1:dsRNA,IFIH1{ub}:dsRNA,IFIH1{ub}:dsRNA:MAVS,IFN inducible genes,IKB-alpha,IKK,IKK-alpha,IKK-beta,IKK-gamma,IKK-i,IKK{active},IRAK,IRAK-4,IRF-3,IRF-3{p},IRF-3{p}:IRF-3{p},IRF-3{p}:IRF-3{p}:ISRE,IRF-7,IRF-7{p},ISRE,LPS,LPS:TLR4,LPS:TLR4:TRIF,LPS:TLR4:TRIF:TRAF3,LPS:TLR4:TRIF:TRAF3:TANK:TBK1:IKK-i,LPS:TLR4:Tollip,LPS:TLR4:Tollip:Mal,LPS:TLR4:Tollip:Mal:MyD88,LPS:TLR4:Tollip:Mal:MyD88:IRAK1,LPS:TLR4:Tollip:Mal:MyD88:IRAK1{p},LPS:TLR4:Tollip:Mal:MyD88:IRAK1{p}:TRAF6,LPS:TLR4:Tollip:Mal:MyD88:IRAK1{p}:TRAF6:TAB2:TAB3:TAK1,LT-beta,LT-beta:LT-betaR,LT-betaR,MAL,MAVS,MyD88,NAP1:TBK1,NAP1:TBK1:IKK-i,NAP1:TBK1:IKK-i{active},NCKAP1(NAP1),NIK,NIK{active},PKR(DAI),RIP1,RIP1{ub},RelA-p65,RelA-p65{p},RelB,SINTBAD,TAB2,TAB2:TAB3:TAK1,TAB3,TAK1,TAK1:TAB2,TANK,TANK:IKK-gamma,TANK:IRF-7,TANK:TBK1:IKK-i,TANK:TBK1:IKK-i:IKK,TANK:TBK1:IKK-i{active},TANK{p},TANK{p}{ub},TBK1,TBK1:IKK-i:SINTBAD,TBK1:IKK-i:SINTBAD{active},TLR3,TLR3:dsRNA:TRIF,TLR3:dsRNA:TRIF:TRAF3,TLR3:dsRNA:TRIF:TRAF3:NAP1:IKK-i:TBK1,TLR3:dsRNA:TRIF:TRAF3:RIP1,TLR3:dsRNA:TRIF:TRAF3:RIP1:TRAF6,TLR3:dsRNA:TRIF:TRAF3:RIP1{ub}:TRAF6,TLR3:dsRNA:TRIF:TRAF3:RIP1{ub}:TRAF6:TAB2:TAK1,TLR4,TLR7,TLR7:viral nucleic acid,TLR8,TLR8:viral nulceic acid,TLR9,TLR9:viral nucleic acid,TNF-alpha,TNFR1,TRADD,TRAF2,TRAF3,TRAF3{active},TRAF5,TRAF6,TRAF6{active},TRIF,Tollip,dsRNA,dsRNA:TLR3,p100,p100{p},p50,p52,p52:RelB,p65:p50,p65:p50(NF-KappaB),p65:p50(NF-KappaB){active},p65:p50:IKBalpha,p65:p50:IKBalpha{p},p65:p50:IKBalpha{p}{ub},proinflammatory genes,protein remnants,viral nucleic acid
Although the essentiality of zinc for plants and animals has been known for manydecades, the essentiality of zinc for humans was recognized only 40 years ago inthe Middle East. The zinc-deficient patients had severe immune dysfunctions,inasmuch as they died of intercurrent infections by the time they were 25 yearsof age. In our studies in an experimental human model of zinc deficiency, wedocumented decreased serum testosterone level, oligospermia, severe immunedysfunctions mainly affecting T helper cells, hyperammonemia, neurosensorydisorders, and decreased lean body mass. It appears that zinc deficiency isprevalent in the developing world and as many as two billion subjects may begrowth retarded due to zinc deficiency. Besides growth retardation and immunedysfunctions, cognitive impairment due to zinc deficiency also has been reportedrecently. Our studies in the cell culture models showed that the activation ofmany zinc-dependent enzymes and transcription factors were adversely affecteddue to zinc deficiency. In HUT-78 (T helper 0 [Th(0)] cell line), we showed thata decrease in gene expression of interleukin-2 (IL-2) and IL-2 receptoralpha(IL-2Ralpha) were due to decreased activation of nuclear factor-kappaB(NF-kappaB) in zinc deficient cells. Decreased NF-kappaB activation in HUT-78due to zinc deficiency was due to decreased binding of NF-kappaB to DNA,decreased level of NF-kappaB p105 (the precursor of NF-kappaB p50) mRNA,decreased kappaB inhibitory protein (IkappaB) phosphorylation, and decreasedIkappa kappa. These effects of zinc were cell specific. Zinc also is anantioxidant and has anti-inflammatory actions. The therapeutic roles of zinc inacute infantile diarrhea, acrodermatitis enteropathica, prevention of blindnessin patients with age-related macular degeneration, and treatment of common coldwith zinc have been reported. In HL-60 cells (promyelocytic leukemia cell line),zinc enhances the up-regulation of A20 mRNA, which, via TRAF pathway, decreasesNF-kappaB activation, leading to decreased gene expression and generation oftumor necrosis factor-alpha (TNF-alpha), IL-1beta, and IL-8. We have reportedrecently that in both young adults and elderly subjects, zinc supplementationdecreased oxidative stress markers and generation of inflammatory cytokines.
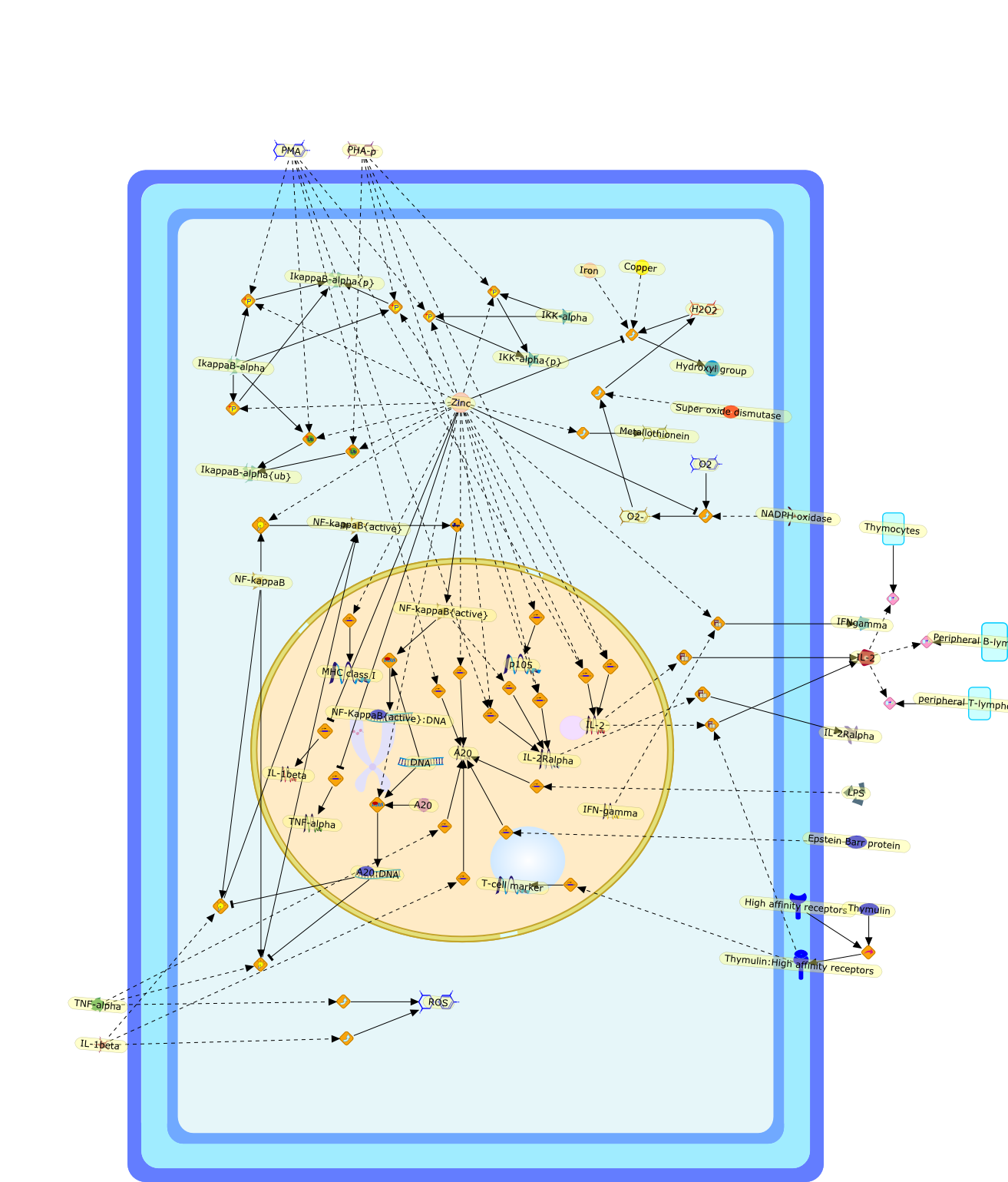{kind=link}
{kind=link}
A20,A20:DNA,Copper,DNA,Epstein Barr protein,H2O2,High affinity receptors,Hydroxyl group,IFN-gamma,IFNgamma,IKK-alpha,IKK-alpha{p},IL-1beta,IL-2,IL-2Ralpha,IkappaB-alpha,IkappaB-alpha{p},IkappaB-alpha{ub},Iron,LPS,MHC class I,Metallothionein,NADPH oxidase,NF-KappaB{active}:DNA,NF-kappaB,NF-kappaB{active},O2,O2-,PHA-p,PMA,Peripheral B-lymphocytes,ROS,Super oxide dismutase,T-cell marker,TNF-alpha,Thymocytes,Thymulin,Thymulin:High affinity receptors,Zinc,p105,peripheral T-lymphocytes
Src-family kinases (SFKs) regulate different granulocyte and monocyte/macrophageresponses. Accumulating evidence suggests that members of this family areimplicated in signal transduction pathways regulating phagocytic cell migrationand recruitment into inflammatory sites. Macrophages with a genetic deficiencyof SFKs display marked alterations in cytoskeleton dynamics, polarization andmigration. This same phenotype is found in cells with either a lack of SFKsubstrates and/or interacting proteins such as Pyk2/FAK, c-Cbl and p190RhoGAP.Notably, SFKs and their downstream targets also regulate monocyte recruitmentinto inflammatory sites. Depending on the type of assay used, neutrophilmigration in vitro may be either dependent on or independent of SFKs. Alsoneutrophil recruitment in in vivo models of inflammation may be regulateddifferently by SFKs depending on the tissue involved. In this review we willdiscuss possible mechanisms by which SFKs may regulate phagocytic cell migratoryabilities.
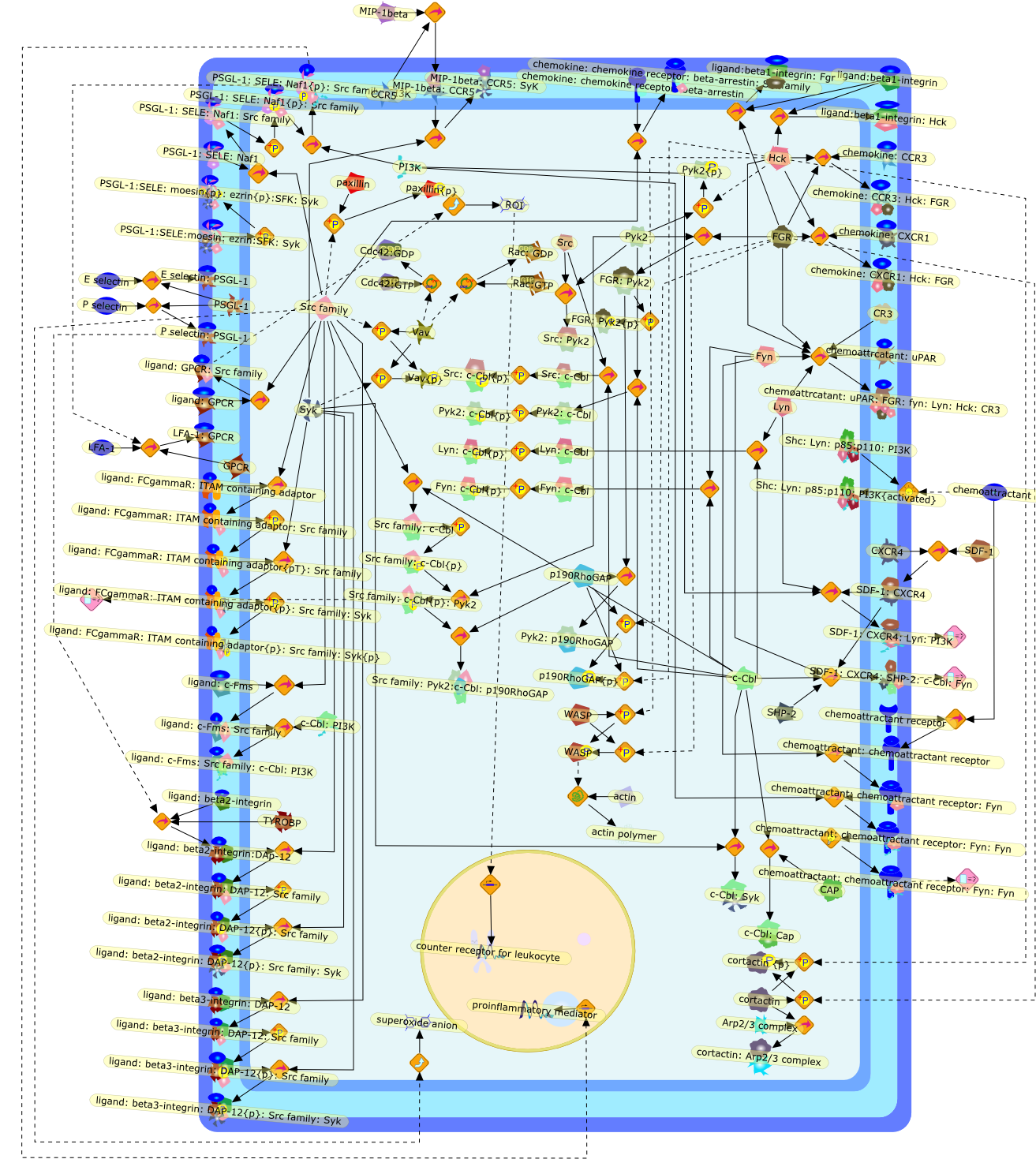{kind=link}
{kind=link}
Arp2/3 complex,CAP,CCR5,CR3,CXCR4,Cdc42:GDP,Cdc42:GTP,E selectin,E selectin: PSGL-1,FGR,FGR: Pyk2,FGR: Pyk2{p},Fyn,Fyn: c-Cbl,Fyn: c-Cbl{p},GPCR,Hck,LFA-1,LFA-1: GPCR,Lyn,Lyn: c-Cbl,Lyn: c-Cbl{p},MIP-1beta,MIP-1beta: CCR5,MIP-1beta: CCR5: SyK,P selectin,P selectin: PSGL-1,PI3K,PSGL-1,PSGL-1: SELE: Naf1,PSGL-1: SELE: Naf1: Src family,PSGL-1: SELE: Naf1{p}: Src family,PSGL-1: SELE: Naf1{p}: Src family: PI3K,PSGL-1:SELE: moesin{p}: ezrin{p}:SFK: Syk,PSGL-1:SELE:moesin: ezrin:SFK: Syk,Pyk2,Pyk2: c-Cbl,Pyk2: c-Cbl{p},Pyk2: p190RhoGAP,Pyk2{p},ROI,Rac: GDP,Rac:GTP,SDF-1,SDF-1: CXCR4,SDF-1: CXCR4: Lyn: PI3K,SDF-1: CXCR4: SHP-2: c-Cbl: Fyn,SHP-2,Shc: Lyn: p85:p110: PI3K,Shc: Lyn: p85:p110: PI3K{activated},Src,Src family,Src family: Pyk2:c-Cbl: p190RhoGAP,Src family: c-Cbl,Src family: c-Cbl{p},Src family: c-Cbl{p}: Pyk2,Src: Pyk2,Src: c-Cbl,Src: c-Cbl{p},Syk,TYROBP,Vav,Vav{p},WASP,actin,actin polymer,c-Cbl,c-Cbl: Cap,c-Cbl: PI3K,c-Cbl: Syk,chemoattractant,chemoattractant receptor,chemoattractant: chemoattractant receptor,chemoattractant: chemoattractant receptor: Fyn,chemoattractant: chemoattractant receptor: Fyn: Fyn,chemoattrcatant: uPAR,chemoattrcatant: uPAR: FGR: fyn: Lyn: Hck: CR3,chemokine: CCR3,chemokine: CCR3: Hck: FGR,chemokine: CXCR1,chemokine: CXCR1: Hck: FGR,chemokine: chemokine receptor: beta-arrestin,chemokine: chemokine receptor: beta-arrestin: Src family,cortactin,cortactin {p},cortactin: Arp2/3 complex,counter receptor for leukocyte,ligand: FCgammaR: ITAM containing adaptor,ligand: FCgammaR: ITAM containing adaptor: Src family,ligand: FCgammaR: ITAM containing adaptor{pT}: Src family,ligand: FCgammaR: ITAM containing adaptor{p}: Src family: Syk,ligand: FCgammaR: ITAM containing adaptor{p}: Src family: Syk{p},ligand: GPCR,ligand: GPCR: Src family,ligand: beta2-integrin,ligand: beta2-integrin: DAP-12: Src family,ligand: beta2-integrin: DAP-12{p}: Src family,ligand: beta2-integrin: DAP-12{p}: Src family: Syk,ligand: beta2-integrin:DAp-12,ligand: beta3-integrin: DAP-12,ligand: beta3-integrin: DAP-12: Src family,ligand: beta3-integrin: DAP-12{p}: Src family,ligand: beta3-integrin: DAP-12{p}: Src family: Syk,ligand: c-Fms,ligand: c-Fms: Src family,ligand: c-Fms: Src family: c-Cbl: PI3K,ligand:beta1-integrin,ligand:beta1-integrin: Fgr,ligand:beta1-integrin: Hck,p190RhoGAP,p190RhoGAP{p},paxillin,paxillin{p},proinflammatory mediator,superoxide anion
Apolipoprotein E has critical roles in the protection against atherosclerosisand is understood to follow the classical constitutive secretion pathway. Recentstudies have indicated that the secretion of apoE from macrophages is aregulated process of unexpected complexity. Cholesterol acceptors such asapolipoprotein A-I, high density lipoprotein, and phospholipid vesicles canstimulate apoE secretion. The ATP binding cassette transporter ABCA1 is involvedin basal apoE secretion and in lipidating apoE-containing particles secreted bymacrophages. However, the stimulation of apoE secretion by apoA-I isABCA1-independent, indicating the existence of both ABCA1-dependent and-independent pathways of apoE secretion. The release of apoE under basalconditions is also regulated, requiring intact protein kinase A activity,intracellular calcium, and an intact microtubular network. Mathematical modelingof apoE turnover indicates that whereas some pools of apoE are committed toeither secretion or degradation, other pools can be diverted from degradationtoward secretion. Targeted inhibition or stimulation of specific apoEtrafficking pathways will provide unique opportunities to regulate the biologyof this important molecule.
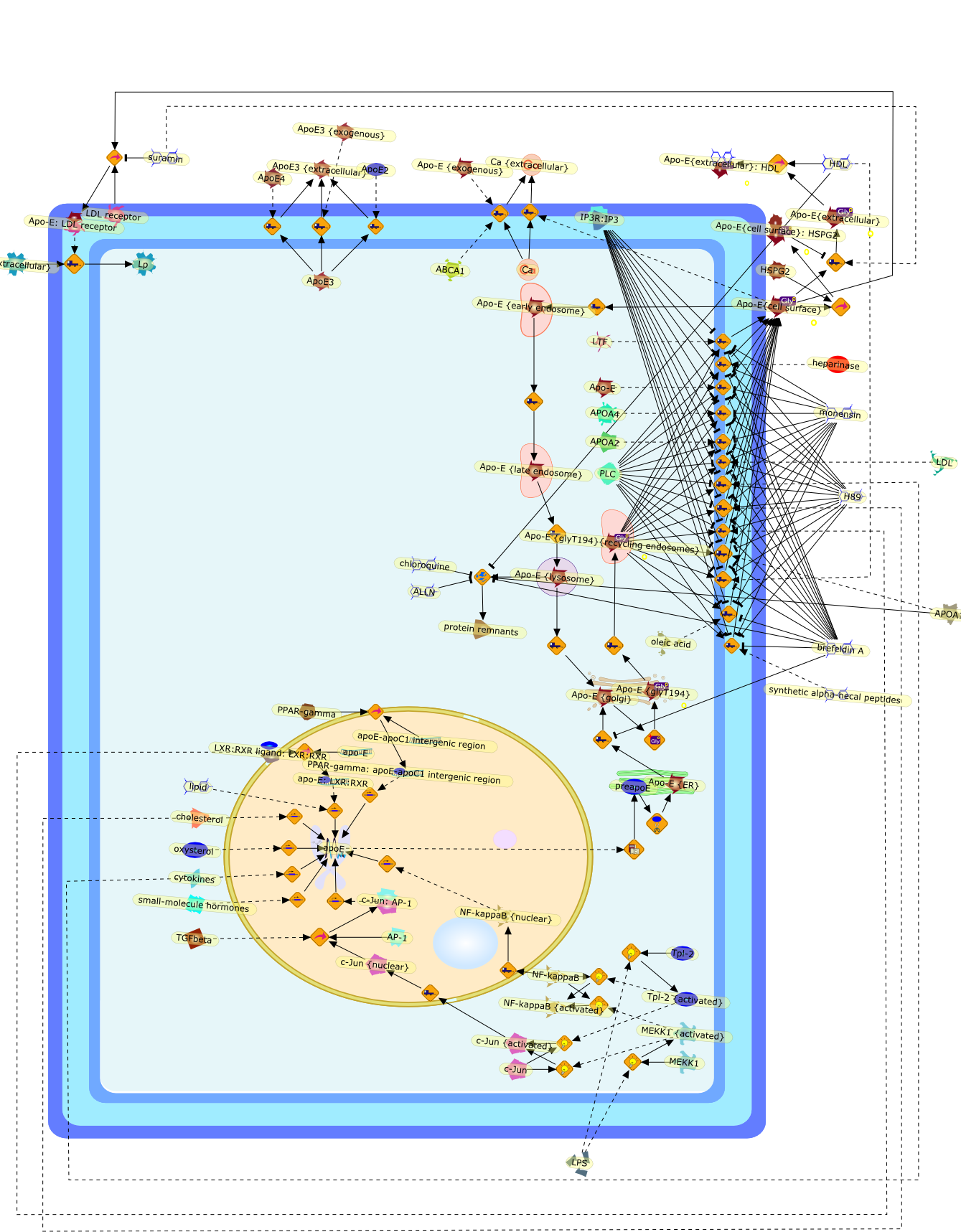{kind=link}
{kind=link}
ABCA1,ALLN,AP-1,APOA1,APOA2,APOA4,Apo-E,Apo-E {ER},Apo-E {early endosome},Apo-E {exogenous},Apo-E {glyT194},Apo-E {glyT194}{recycling endosomes},Apo-E {golgi},Apo-E {late endosome},Apo-E {lysosome},Apo-E: LDL receptor,Apo-E{cell surface},Apo-E{cell surface}: HSPG2,Apo-E{extracellular},Apo-E{extracellular}: HDL,ApoE2,ApoE3,ApoE3 {exogenous},ApoE3 {extracellular},ApoE4,Ca,Ca {extracellular},H89,HDL,HSPG2,IP3R:IP3,LDL,LDL receptor,LPS,LTF,LXR:RXR ligand: LXR:RXR,Lp,Lp {extracellular},MEKK1,MEKK1 {activated},NF-kappaB,NF-kappaB {activated},NF-kappaB {nuclear},PLC,PPAR-gamma,PPAR-gamma: apoE-apoC1 intergenic region,TGFbeta,Tpl-2,Tpl-2 {activated},apo-E,apo-E: LXR:RXR,apoE,apoE-apoC1 intergenic region,brefeldin A,c-Jun,c-Jun {activated},c-Jun {nuclear},c-Jun: AP-1,chloroquine,cholesterol,cytokines,heparinase,lipid,monensin,oleic acid,oxysterol,preapoE,protein remnants,small-molecule hormones,suramin,synthetic alpha-hecal peptides
Cytokine signaling via a restricted number of Jak-Stat pathways positively andnegatively regulates all cell types involved in the initiation, propagation, andresolution of inflammation. Here, we focus on Jak-Stat signaling in three majorcell types involved in inflammatory responses: T cells, neutrophils, andmacrophages. We summarize how the Jak-Stat pathways in these cells arenegatively regulated by the Suppressor of cytokine signaling (Socs) proteins. Weemphasize that common Jak-Stat-Socs signaling modules can have diversedevelopmental, pro- and anti-inflammatory outcomes depending on the cytokinereceptor activated and which genes are accessible at a given time in a cell'slife. Because multiple components of Jak-Stat-Socs pathways are mutated orclosely associated with human inflammatory diseases, and cytokine-basedtherapies are increasingly deployed to treat inflammation, understandingcytokine signaling will continue to advance our ability to manipulate chronicand acute inflammatory diseases.
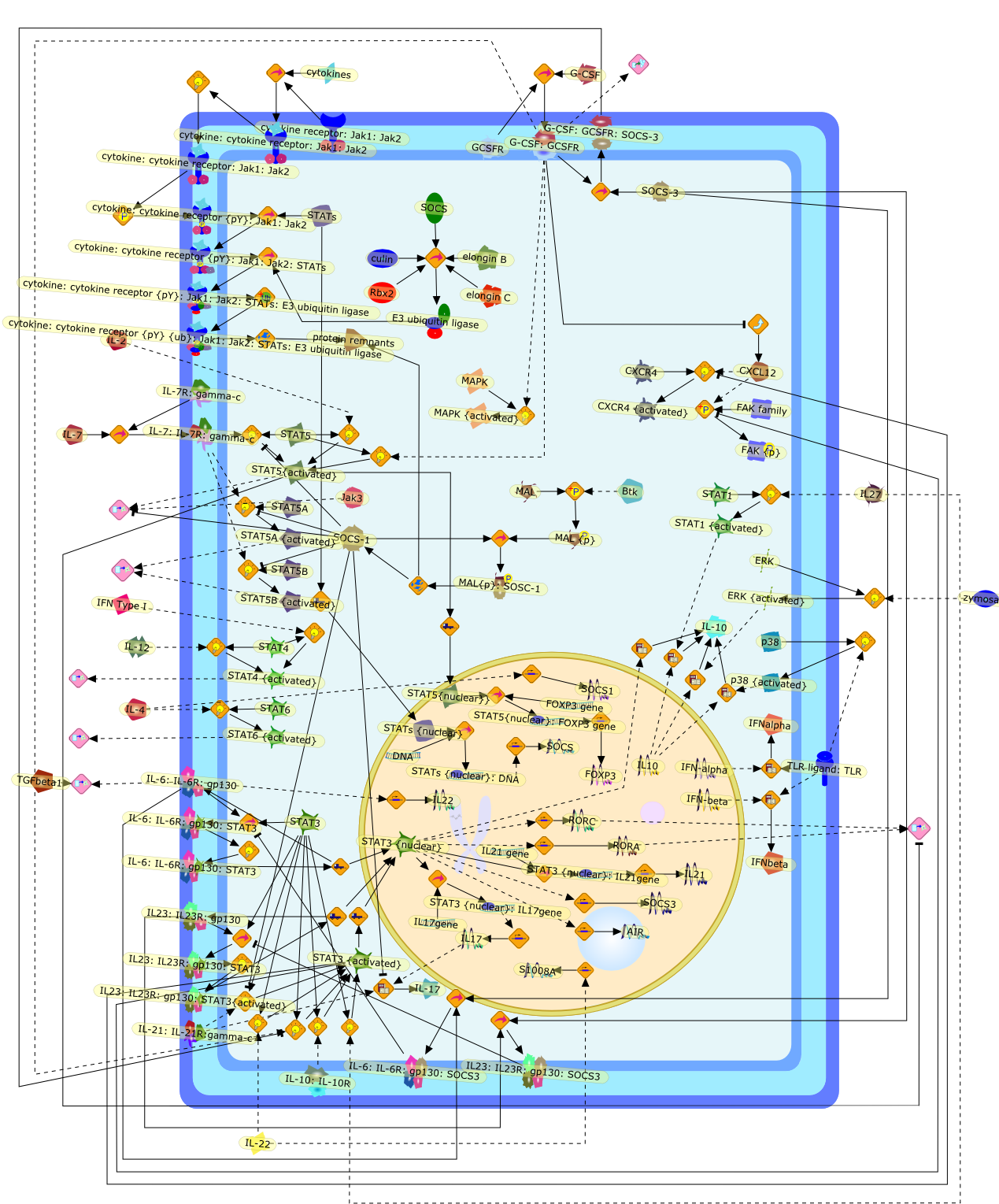{kind=link}
{kind=link}
AIR,Btk,CXCL12,CXCR4,CXCR4 {activated},DNA,E3 ubiquitin ligase,ERK,ERK {activated},FAK family,FAK {p},FOXP3,FOXP3 gene,G-CSF,G-CSF: GCSFR,G-CSF: GCSFR: SOCS-3,GCSFR,IFN Type I,IFN-alpha,IFN-beta,IFNalpha,IFNbeta,IL-10,IL-10: IL-10R,IL-12,IL-17,IL-2,IL-21: IL-21R:gamma-c,IL-22,IL-4,IL-6: IL-6R: gp130,IL-6: IL-6R: gp130: SOCS3,IL-6: IL-6R: gp130: STAT3,IL-7,IL-7: IL-7R: gamma-c,IL-7R: gamma-c,IL10,IL17,IL17gene,IL21,IL21 gene,IL22,IL23: IL23R: gp130,IL23: IL23R: gp130: SOCS3,IL23: IL23R: gp130: STAT3,IL23: IL23R: gp130: STAT3{activated},IL27,Jak3,MAL,MAL {p},MAL{p}: SOSC-1,MAPK,MAPK {activated},RORA,RORC,Rbx2,S1008A,SOCS,SOCS-1,SOCS-3,SOCS1,SOCS3,STAT1,STAT1 {activated},STAT3,STAT3 {activated},STAT3 {nuclear},STAT3 {nuclear}: IL17gene,STAT3 {nuclear}: IL21gene,STAT4,STAT4 {activated},STAT5,STAT5A,STAT5A {activated},STAT5B,STAT5B {activated},STAT5{activated},STAT5{nuclear}: FOXP3 gene,STAT5{nuclear}},STAT6,STAT6 {activated},STATs,STATs {nuclear},STATs {nuclear}: DNA,TGFbeta1,TLR ligand: TLR,culin,cytokine receptor: Jak1: Jak2,cytokine: cytokine receptor {pY} {ub}: Jak1: Jak2: STATs: E3 ubiquitin ligase,cytokine: cytokine receptor {pY}: Jak1: Jak2,cytokine: cytokine receptor {pY}: Jak1: Jak2: STATs,cytokine: cytokine receptor {pY}: Jak1: Jak2: STATs: E3 ubiquitin ligase,cytokine: cytokine receptor: Jak1: Jak2,cytokines,elongin B,elongin C,p38,p38 {activated},protein remnants,zymosan
In this review, we summarize our investigations concerning the differentialimportance of CD14 and LBP in toll-like receptor 4 (TLR4)/myeloiddifferentiation protein-2 (MD-2)-mediated signaling by smooth and rough-formlipopolysaccharide (LPS) chemotypes and include the results obtained in studieswith murine and human TLR4-transgenic mice. Furthermore, we present more recentdata on the mechanisms involved in the induction of LPS hypersensitivity bybacterial and viral infections and on the reactivity of the hypersensitive hostto non-LPS microbial ligands and endogenous mediators. Finally, the effects ofpre-existing hypersensitivity on the course and outcome of a super-infectionwith Salmonella typhimurium or Listeria monocytogenes are summarized.
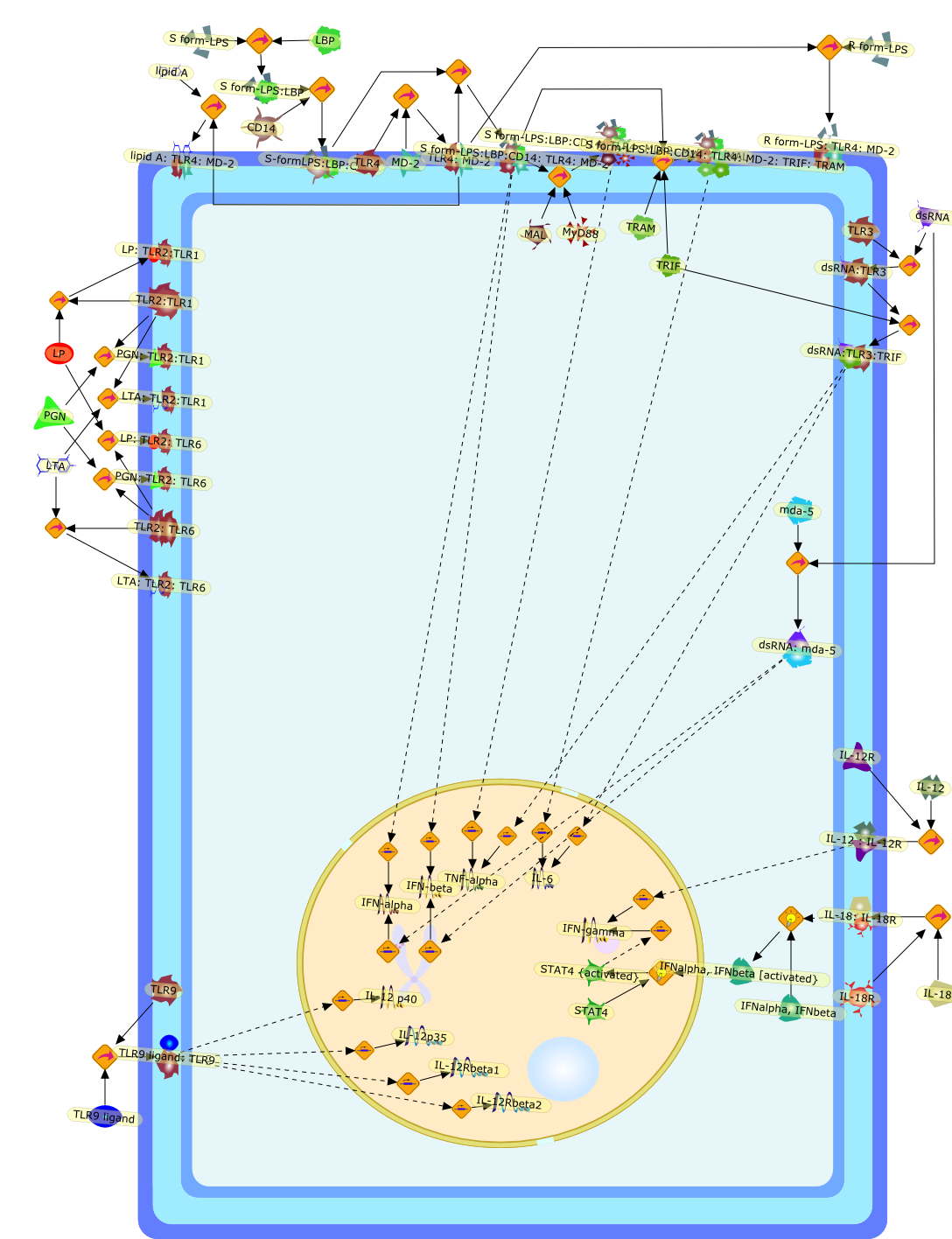{kind=link}
{kind=link}
CD14,IFN-alpha,IFN-beta,IFN-gamma,IFNalpha, IFNbeta,IFNalpha, IFNbeta [activated},IL-12,IL-12 : IL-12R,IL-12 p40,IL-12R,IL-12Rbeta1,IL-12Rbeta2,IL-12p35,IL-18,IL-18: IL-18R,IL-18R,IL-6,LBP,LP,LP: TLR2: TLR6,LP: TLR2:TLR1,LTA,LTA: TLR2: TLR6,LTA: TLR2:TLR1,MAL,MD-2,MyD88,PGN,PGN: TLR2: TLR6,PGN: TLR2:TLR1,R form-LPS,R form-LPS: TLR4: MD-2,S form-LPS,S form-LPS:LBP,S form-LPS:LBP:CD14: TLR4: MD-2,S form-LPS:LBP:CD14: TLR4: MD-2: MyD88: MAL,S form-LPS:LBP:CD14: TLR4: MD-2: TRIF: TRAM,S-formLPS:LBP:CD14,STAT4,STAT4 {activated},TLR2: TLR6,TLR2:TLR1,TLR3,TLR4,TLR4: MD-2,TLR9,TLR9 ligand,TLR9 ligand: TLR9,TNF-alpha,TRAM,TRIF,dsRNA,dsRNA: mda-5,dsRNA:TLR3,dsRNA:TLR3:TRIF,lipid A,lipid A: TLR4: MD-2,mda-5
Innate immunity represents the first line of defense against invading pathogens.Toll-like receptors (TLRs) are important for activation of innate immunity.Moreover, cytokines mediate communication of cells and are necessary to mount anappropriately regulated immune response. However, activation of innate immunityhas to be tightly controlled to avoid overshooting immune reactions. Suppressorof cytokine signaling (SOCS) proteins have been identified as inducible feedbackinhibitors of cytokine receptors and have been shown to be of crucial importancefor the limitation of inflammatory responses. In this review, we describe therole of SOCS proteins in macrophages and dendritic cells (DCs). Based on our ownfindings, we show that SOCS proteins are directly induced by stimulation ofTLRs. However, SOCS proteins do not interfere with direct TLR signaling, butavoid overshooting activation by regulating paracrine IFN-beta signaling. Inaddition, SOCS proteins in macrophages and DCs regulate the sensitivity towardsIFN-gamma and GM-CSF, thereby modulating anti-microbial activity of macrophagesand differentiation of DCs. We discuss that SOCS induction can also be used bymicrobes to evade immune defense, and this is exemplified by the parasiteToxoplasma gondii which induces SOCS1 to inhibit IFN-gamma-mediated macrophageactivation. Taken together, the findings indicate that SOCS proteins play animportant role in the balanced activation of innate immunity during infectiousencounter.
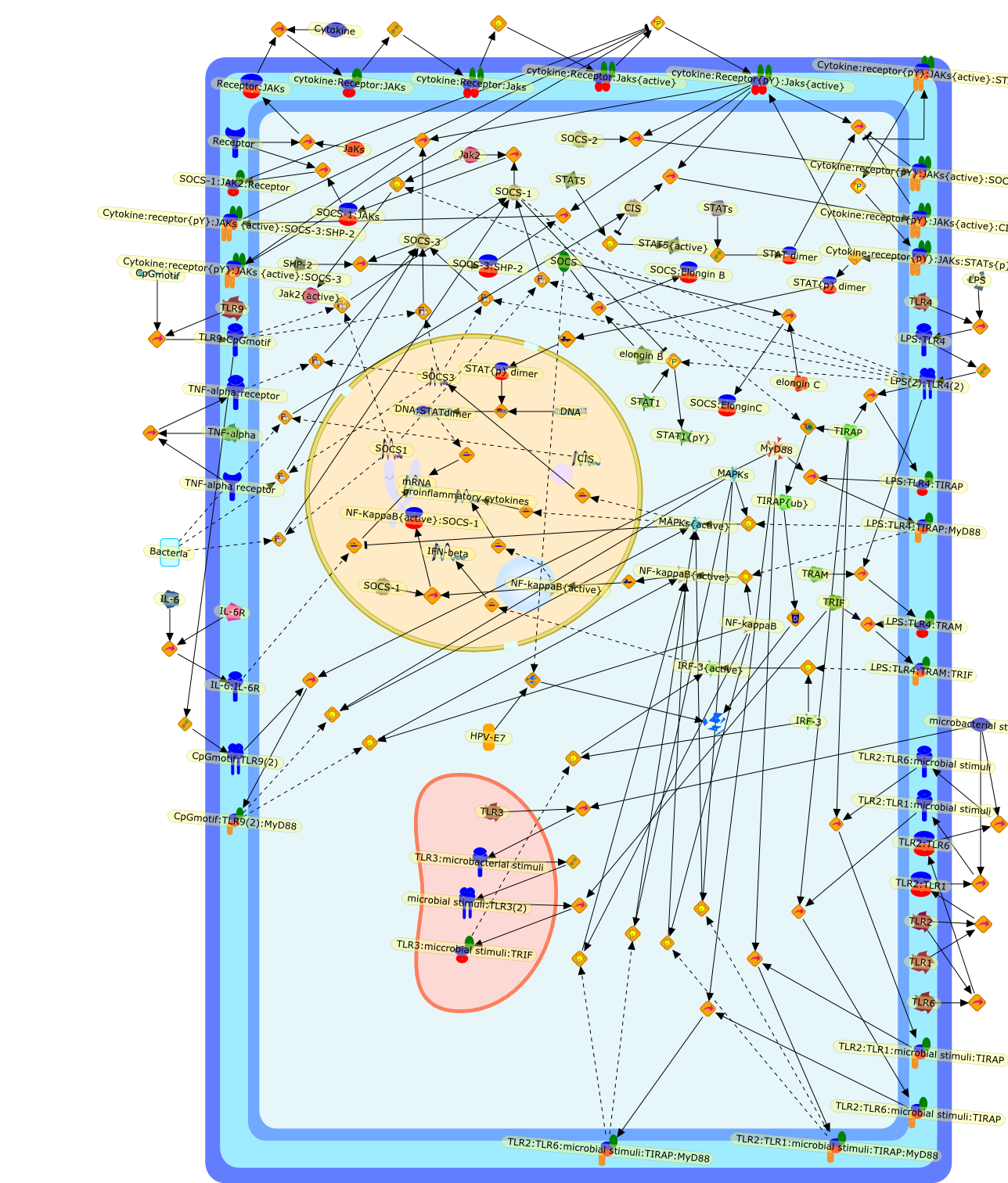{kind=link}
{kind=link}
Bacteria,CIS,CpGmotif,CpGmotif:TLR9(2),CpGmotif:TLR9(2):MyD88,Cytokine,Cytokine:receptor{pY}:JAKs {active}:SOCS-3,Cytokine:receptor{pY}:JAKs {active}:SOCS-3:SHP-2,Cytokine:receptor{pY}:JAKs:STATs{p},Cytokine:receptor{pY}:JAKs{active}:CIS,Cytokine:receptor{pY}:JAKs{active}:SOCS-2,Cytokine:receptor{pY}:JAKs{active}:STATs,DNA,DNA:STATdimer,HPV-E7,IFN-beta,IL-6,IL-6:IL-6R,IL-6R,IRF-3,IRF-3{active},JaKs,Jak2,Jak2{active},LPS,LPS(2):TLR4(2),LPS:TLR4,LPS:TLR4:TIRAP,LPS:TLR4:TIRAP:MyD88,LPS:TLR4:TRAM,LPS:TLR4:TRAM:TRIF,MAPKs,MAPKs{active},MyD88,NF-KappaB{active}:SOCS-1,NF-kappaB,NF-kappaB{active},Receptor,Receptor:JAKs,SHP-2,SOCS,SOCS-1,SOCS-1:JAK2:Receptor,SOCS-1:JAKs,SOCS-2,SOCS-3,SOCS-3:SHP-2,SOCS1,SOCS3,SOCS:Elongin B,SOCS:ElonginC,STAT dimer,STAT1,STAT1{pY},STAT5,STAT5{active},STATs,STAT{p} dimer,TIRAP,TIRAP{ub},TLR1,TLR2,TLR2:TLR1,TLR2:TLR1:microbial stimuli,TLR2:TLR1:microbial stimuli:TIRAP,TLR2:TLR1:microbial stimuli:TIRAP:MyD88,TLR2:TLR6,TLR2:TLR6:microbial stimuli,TLR2:TLR6:microbial stimuli:TIRAP,TLR2:TLR6:microbial stimuli:TIRAP:MyD88,TLR3,TLR3:miccrobial stimuli:TRIF,TLR3:microbacterial stimuli,TLR4,TLR6,TLR9,TLR9:CpGmotif,TNF-alpha,TNF-alpha receptor,TNF-alpha:receptor,TRAM,TRIF,cytokine:Receptor:JAKs,cytokine:Receptor:Jaks,cytokine:Receptor:Jaks{active},cytokine:Receptor{pY}:Jaks{active},dergradants,elongin B,elongin C,mRNA,microbacterial stimuli,microbial stimuli:TLR3(2),proinflammatory cytokines
One-third of the world's population is infected with Mycobacterium tuberculosis,one of the most effective human pathogens, whose success is attributed to thedeployment of remarkably sophisticated immune evasion mechanisms. In this issueof Cell Host & Microbe, a new study unravels a novel strategy of immune evasionand enhanced bacterial intracellular survival, which is dependent on inhibitionof inflammasome activation by an M. tuberculosis-encoded metalloprotease.
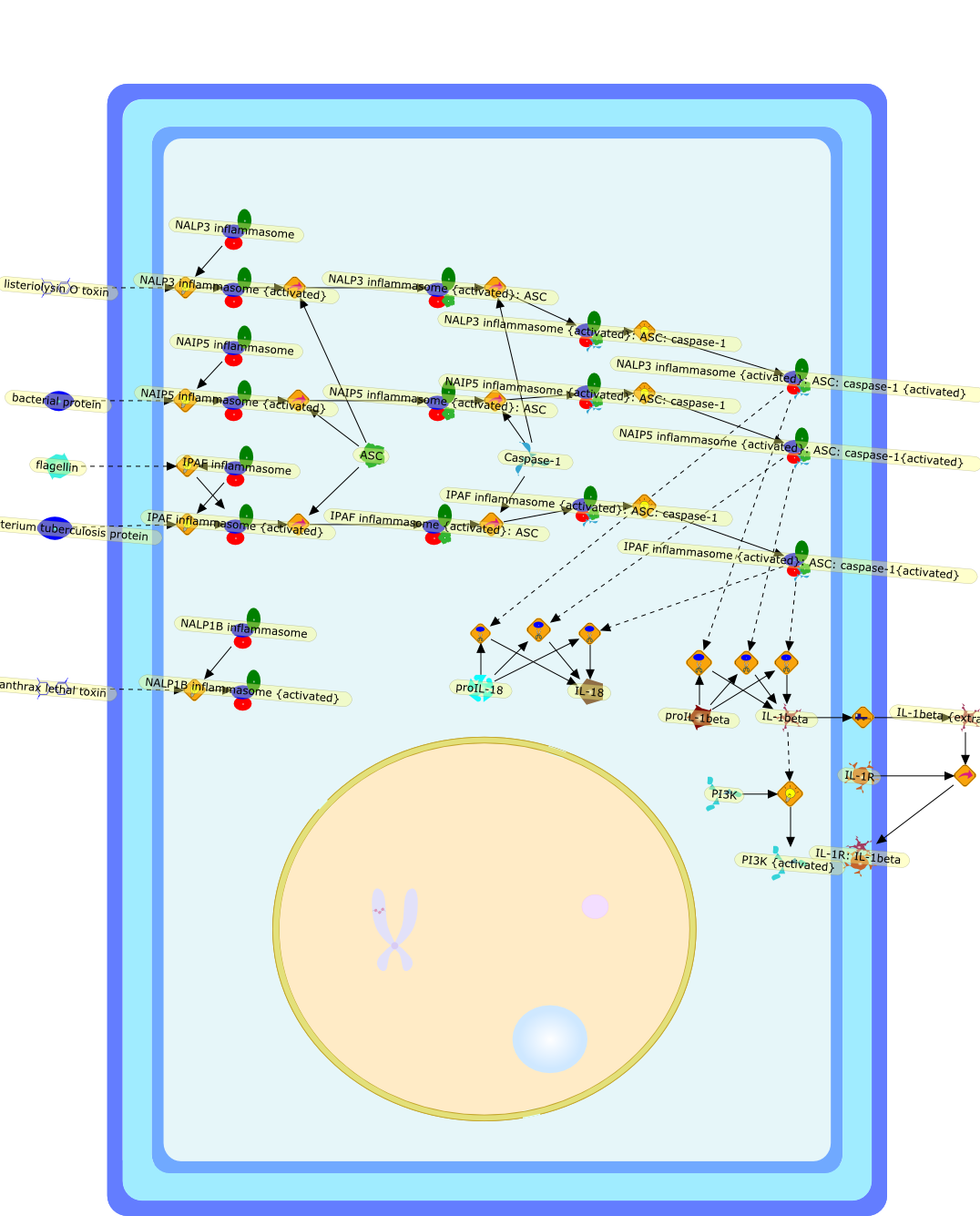{kind=link}
{kind=link}
ASC,Caspase-1,IL-18,IL-1R,IL-1R: IL-1beta,IL-1beta,IL-1beta {extracellular},IPAF inflammasome,IPAF inflammasome {activated},IPAF inflammasome {activated}: ASC,IPAF inflammasome {activated}: ASC: caspase-1,IPAF inflammasome {activated}: ASC: caspase-1{activated},Mycobacterium tuberculosis protein,NAIP5 inflammasome,NAIP5 inflammasome {activated},NAIP5 inflammasome {activated}: ASC,NAIP5 inflammasome {activated}: ASC: caspase-1,NAIP5 inflammasome {activated}: ASC: caspase-1{activated},NALP1B inflammasome,NALP1B inflammasome {activated},NALP3 inflammasome,NALP3 inflammasome {activated},NALP3 inflammasome {activated}: ASC,NALP3 inflammasome {activated}: ASC: caspase-1,NALP3 inflammasome {activated}: ASC: caspase-1 {activated},PI3K,PI3K {activated},anthrax lethal toxin,bacterial protein,flagellin,listeriolysin O toxin,proIL-18,proIL-1beta
Experimental perturbation of signaling or transcription factor networks has beenused to study the developmental potential of lymphoid progenitors,lineage-committed precursors and mature lymphocytes. Common lymphoid progenitorsand uncommitted pro-T cells can be efficiently diverted into myeloid orerythroid lineages by ectopic cytokine signaling or retroviral expression of themyeloid C/EBPalpha or erythroid GATA1 transcription factor. Forced C/EBPalphaexpression furthermore induces direct transdifferentiation of immaturethymocytes or B cells into macrophages. Notably, conditional inactivation of theB cell commitment factor Pax5 is sufficient to convert mature B cells intofunctional T cells via dedifferentiation to uncommitted progenitors. Togetherthese experiments have uncovered an unanticipated developmental plasticity oflymphocytes, which may account for lineage switches observed in humanmalignancies.
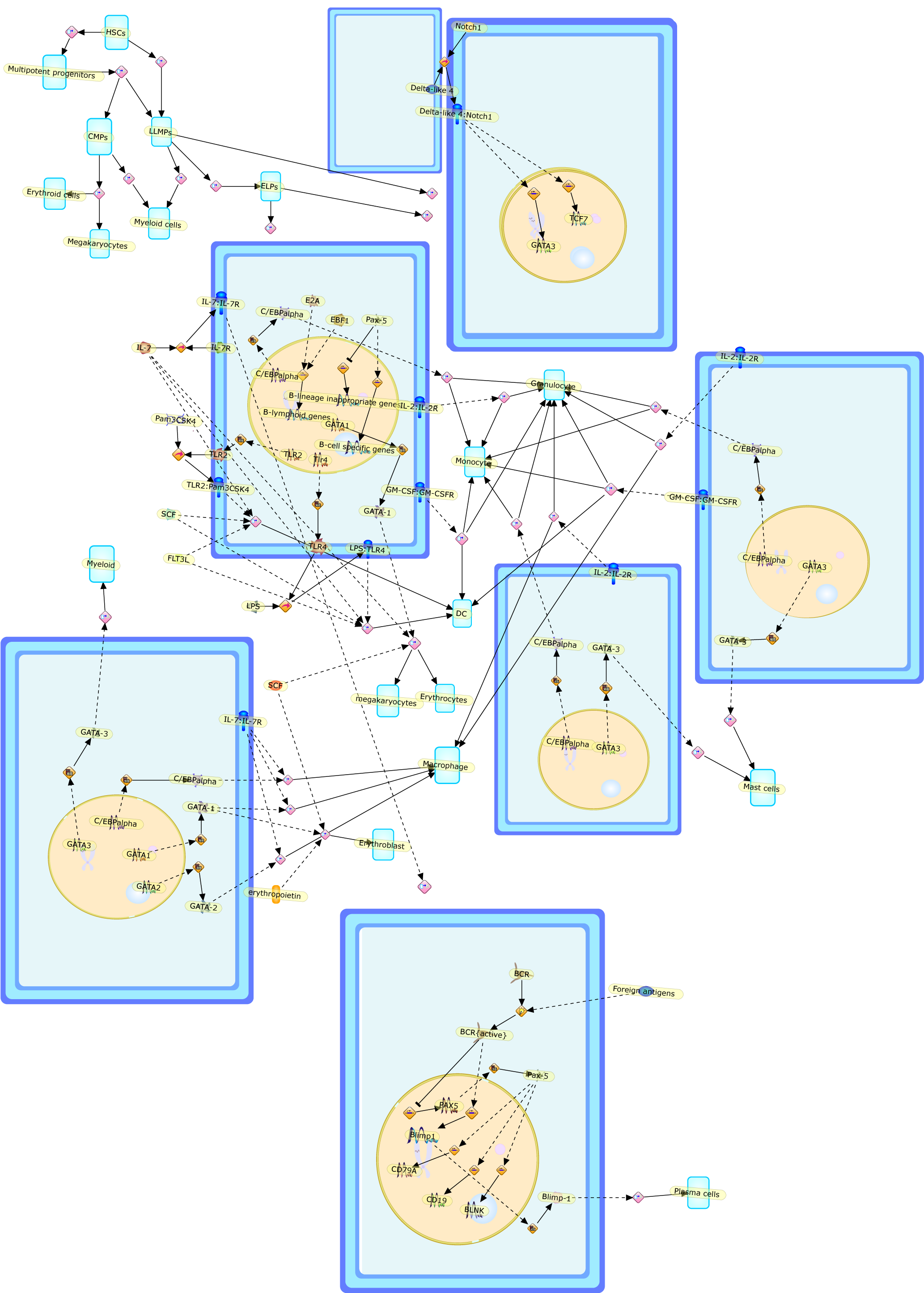{kind=link}
{kind=link}
B-cell specific genes,B-lineage inappropriate genes,B-lymphoid genes,BCR,BCR{active},BLNK,Blimp-1,Blimp1,C/EBPalpha,CD19,CD79A,CMPs,DC,Delta-like 4,Delta-like 4:Notch1,E2A,EBF1,ELPs,Erythroblast,Erythrocytes,Erythroid cells,FLT3L,Foreign antigens,GATA-1,GATA-2,GATA-3,GATA1,GATA2,GATA3,GM-CSF:GM-CSFR,Granulocyte,HSCs,IL-2:IL-2R,IL-7,IL-7:IL-7R,IL-7R,LLMPs,LPS,LPS:TLR4,Macrophage,Mast cells,Megakaryocytes,Monocyte,Multipotent progenitors,Myeloid,Myeloid cells,Notch1,PAX5,Pam3CSK4,Pax-5,Plasma cells,SCF,TCF7,TLR2,TLR2:Pam3CSK4,TLR4,Tlr4,erythropoietin,megakaryocytes
In cells of the immune system, calcium signals are essential for diversecellular functions including differentiation, effector function, and genetranscription. After the engagement of immunoreceptors such as T-cell and B-cellantigen receptors and the Fc receptors on mast cells and NK cells, theintracellular concentration of calcium ions is increased through the sequentialoperation of two interdependent processes: depletion of endoplasmic reticulumCa(2+) stores as a result of binding of inositol trisphosphate (IP(3)) to IP(3)receptors, followed by 'store-operated' Ca(2+) entry through plasma membraneCa(2+) channels. In lymphocytes, mast cells and other immune cell types,store-operated Ca(2+) entry through specialized Ca(2+) release-activated calcium(CRAC) channels constitutes the major pathway of intracellular Ca(2+) increase.A recent breakthrough in our understanding of CRAC channel function is theidentification of stromal interaction molecule (STIM) and ORAI, two essentialregulators of CRAC channel function. This review focuses on the signalingpathways upstream and downstream of Ca(2+) influx (the STIM/ORAI andcalcineurin/NFAT pathways, respectively).
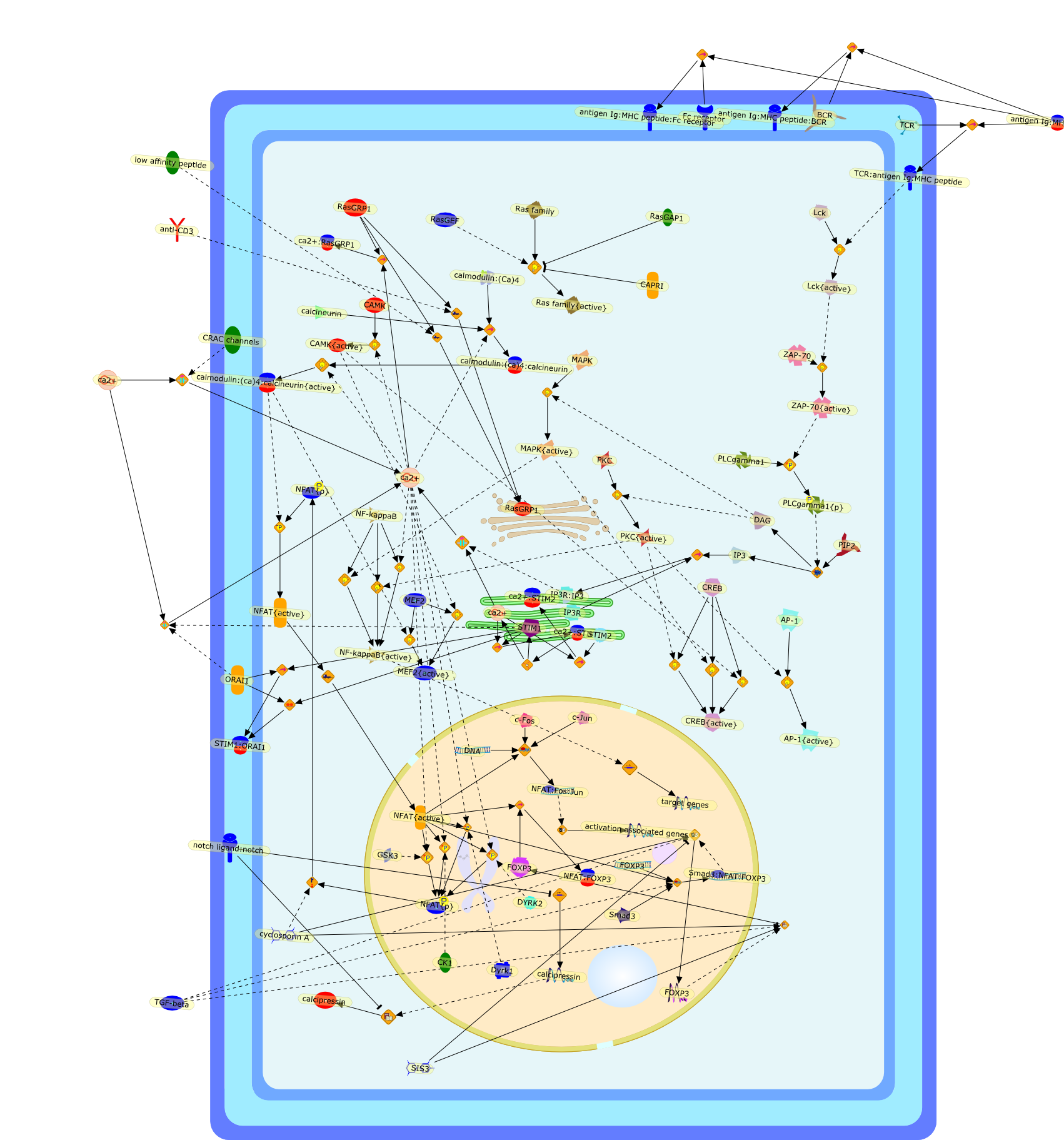{kind=link}
{kind=link}
AP-1,AP-1{active},BCR,CAMK,CAMK{active},CAPRI,CK1,CRAC channels,CREB,CREB{active},DAG,DNA,DYRK2,Dyrk1,FOXP3,Fc receptor,GSK3,IP3,IP3R,IP3R:IP3,Lck,Lck{active},MAPK,MAPK{active},MEF2,MEF2{active},NF-kappaB,NF-kappaB{active},NFAT:FOXP3,NFAT:Fos:Jun,NFAT{active},NFAT{p},ORAI1,PIP2,PKC,PKC{active},PLCgamma1,PLCgamma1{p},Ras family,Ras family{active},RasGAP1,RasGEF,RasGRP1,SIS3,STIM1,STIM1:ORAI1,STIM2,Smad3,Smad3:NFAT:FOXP3,TCR,TCR:antigen Ig:MHC peptide,TGF-beta,ZAP-70,ZAP-70{active},activation associated genes,anti-CD3,antigen Ig:MHC peptide,antigen Ig:MHC peptide:BCR,antigen Ig:MHC peptide:Fc receptor,c-Fos,c-Jun,ca2+,ca2+:RasGRP1,ca2+:STIM1,ca2+:STIM2,calcineurin,calcipressin,calmodulin:(Ca)4,calmodulin:(ca)4:calcineurin,calmodulin:(ca)4:calcineurin{active},cyclosporin A,low affinity peptide,notch ligand:notch,target genes
Dendritic cells (DCs) produce an array of cytokines after detecting variousimmune adjuvants through pattern recognition receptors (PRRs). PRR signalingleads to activation of transcription factors such as NF-kappaB or interferonregulatory factors (IRFs) but after activation must be attenuated to avoidimmunopathology and to maintain tissue homeostasis. IkappaB kinase familymembers, originally identified as classical NF-kappaB activators, are now foundto be broadly and crucially involved in PRR signaling in a member-specificmanner. Furthermore, a new mechanism for NF-kappaB downregulation is emergingthat involves the degradation of active NF-kappaB by the nuclearubiquitin-proteasome system. Here we review new aspects of NF-kappaB and IRFregulation chiefly in DCs.
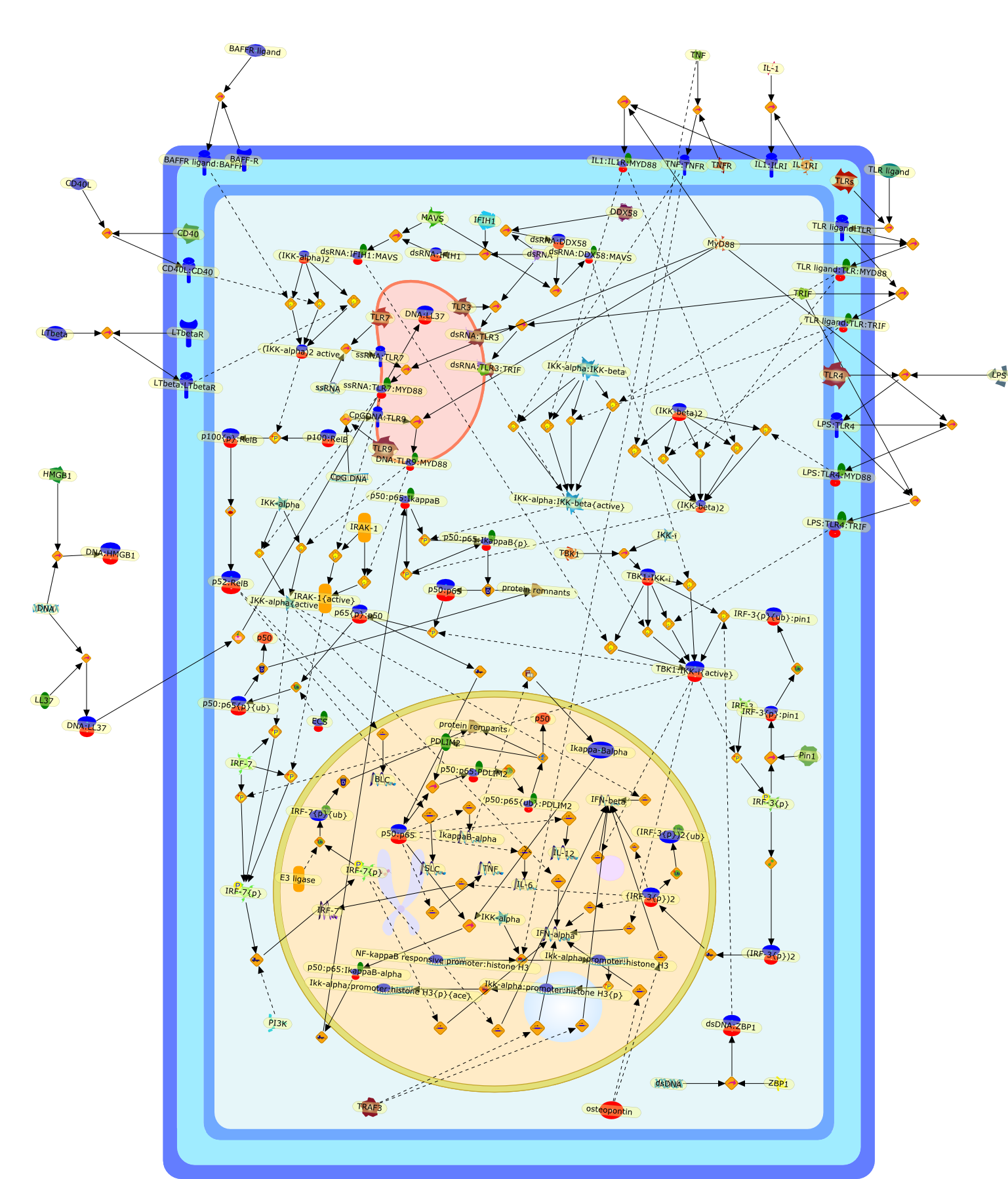{kind=link}
{kind=link}
(IKK-alpha)2,(IKK-alpha)2 active,(IKK-beta)2,(IRF-3{p})2,(IRF-3{p})2{ub},BAFF-R,BAFFR ligand,BAFFR ligand:BAFFR,BLC,CD40,CD40L,CD40L:CD40,CpG DNA,CpGDNA:TLR9,DDX58,DNA,DNA:HMGB1,DNA:LL37,DNA:TLR9:MYD88,E3 ligase,ECS,HMGB1,IFIH1,IFN-alpha,IFN-beta,IKK-alpha,IKK-alpha:IKK-beta,IKK-alpha:IKK-beta{active},IKK-alpha{active},IKK-i,IL-1,IL-12,IL-1RI,IL-6,IL1:IL1R:MYD88,IL1:ILRI,IRAK-1,IRAK-1{active},IRF-3,IRF-3{p},IRF-3{p}:pin1,IRF-3{p}{ub}:pin1,IRF-7,IRF-7{p},IRF-7{p}{ub},Ikappa-Balpha,IkappaB-alpha,Ikk-alpha:promoter:histone H3,Ikk-alpha:promoter:histone H3{p},Ikk-alpha:promoter:histone H3{p}{ace},LL37,LPS,LPS:TLR4,LPS:TLR4:MYD88,LPS:TLR4:TRIF,LTbeta,LTbeta:LTbetaR,LTbetaR,MAVS,MyD88,NF-kappaB responsive promoter:histone H3,PDLIM2,PI3K,Pin1,SLC,TBK1,TBK1:IKK-i,TBK1:IKK-i{active},TLR ligand,TLR ligand:TLR,TLR ligand:TLR:MYD88,TLR ligand:TLR:TRIF,TLR3,TLR4,TLR7,TLR9,TLRs,TNF,TNF:TNFR,TNFR,TRAF3,TRIF,ZBP1,dsDNA,dsDNA:ZBP1,dsRNA,dsRNA:DDX58,dsRNA:DDX58:MAVS,dsRNA:IFIH1,dsRNA:IFIH1:MAVS,dsRNA:TLR3,dsRNA:TLR3:TRIF,osteopontin,p100:RelB,p100{p}:RelB,p50,p50:p65,p50:p65:IkappaB,p50:p65:IkappaB-alpha,p50:p65:IkappaB{p},p50:p65:PDLIM2,p50:p65{p}{ub},p50:p65{ub}:PDLIM2,p52:RelB,p65{p}:p50,protein remnants,ssRNA,ssRNA:TLR7,ssRNA:TLR7:MYD88
Mitochondrial antiviral immunity involves the detection of viral RNA byintracellular pattern-recognition receptors (PRRs) belonging to the RIG-I-likehelicase family. The convergence of these and other signaling molecules to theouter mitochondrial membrane results in the rapid induction of antiviralcytokines including type-1 interferon. Here, we discuss recent studiesdescribing new molecules implicated in the regulation of this antiviralresponse.
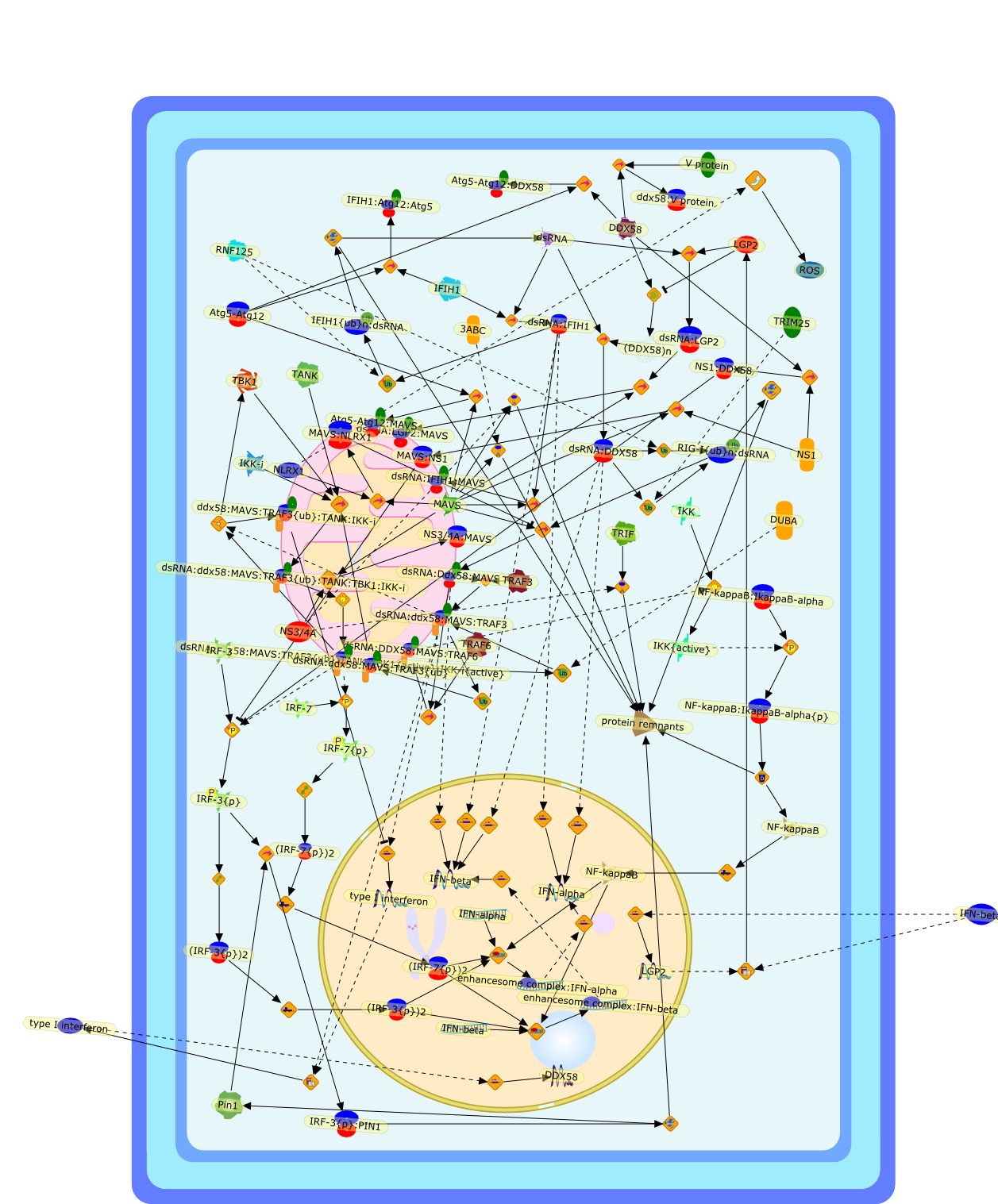{kind=link}
{kind=link}
(DDX58)n,(IRF-3{p})2,(IRF-7{p})2,3ABC,Atg5-Atg12,Atg5-Atg12:DDX58,Atg5-Atg12:MAVS,DDX58,DUBA,IFIH1,IFIH1:Atg12:Atg5,IFIH1{ub}n:dsRNA,IFN-alpha,IFN-beta,IKK,IKK-i,IKK{active},IRF-3,IRF-3{p},IRF-3{p}:PIN1,IRF-7,IRF-7{p},LGP2,MAVS,MAVS:NLRX1,MAVS:NS1,NF-kappaB,NF-kappaB:IkappaB-alpha,NF-kappaB:IkappaB-alpha{p},NLRX1,NS1,NS1:DDX58,NS3/4A,NS3/4A:MAVS,Pin1,RIG-I{ub}n:dsRNA,RNF125,ROS,TANK,TBK1,TRAF3,TRAF6,TRIF,TRIM25,V protein,ddx58:MAVS:TRAF3{ub}:TANK:IKK-i,ddx58:V protein,dsRNA,dsRNA:DDX58,dsRNA:DDX58:MAVS:TRAF6,dsRNA:Ddx58:MAVS,dsRNA:IFIH1,dsRNA:IFIH1:MAVS,dsRNA:LGP2,dsRNA:LGP2:MAVS,dsRNA:ddx58:MAVS:TRAF3,dsRNA:ddx58:MAVS:TRAF3{ub},dsRNA:ddx58:MAVS:TRAF3{ub}:TANK:TBK1:IKK-i,dsRNA:ddx58:MAVS:TRAF3{ub}:TANK:TBK1{active}:IKK-i{active},enhancesome complex:IFN-alpha,enhancesome complex:IFN-beta,protein remnants,type I interferon
Mammalian innate immunity possesses a distinct system to recognize aberrant DNAinside the cell. One class of DNA sensors is the Toll-like receptor 9, which isexpressed in the specialized immune cells, binds to single-stranded DNA in theendosome to transmit cellular signaling through myeloid differentiation primaryresponse protein 88 (MyD88). Another class of DNA sensors exists in thecytoplasm of most type of cells in the tissue, detecting double-stranded DNA tosignal through TANK-binding kinase-1 (TBK1)-mediated type-I interferonproduction and apoptosis-associated speck-like protein containing a CARD(ASC)-mediated IL-1beta secretion. Since DNA sensors have potential to recognizeaberrant DNA of both self and nonself origin, their physiological roles inmicrobial infection, tissue damage, autoimmune diseases, and DNA-basedtherapeutic applications are being intensively investigated.
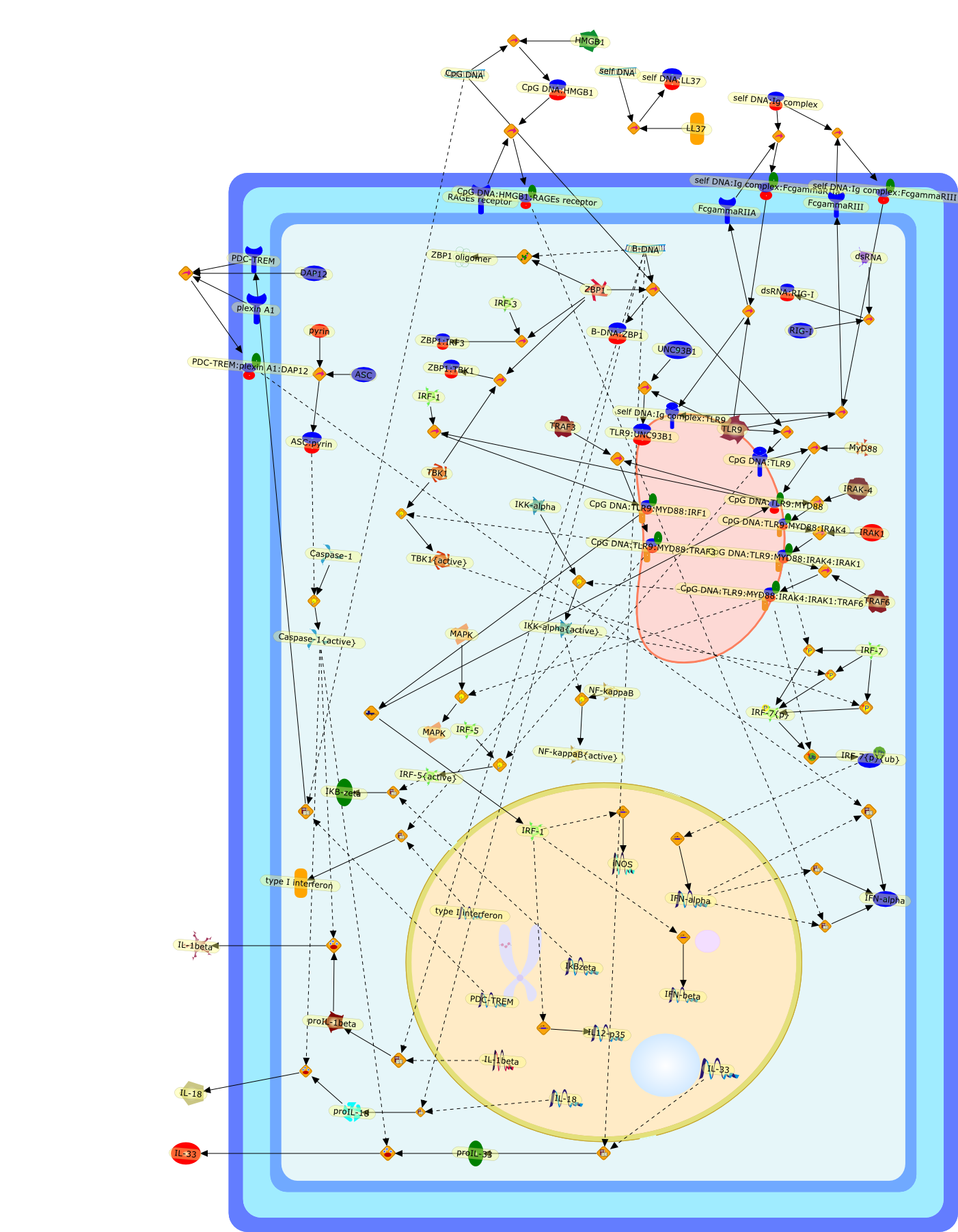{kind=link}
{kind=link}
ASC,ASC:pyrin,B-DNA,B-DNA:ZBP1,Caspase-1,Caspase-1{active},CpG DNA,CpG DNA:HMGB1,CpG DNA:HMGB1:RAGEs receptor,CpG DNA:TLR9,CpG DNA:TLR9:MYD88,CpG DNA:TLR9:MYD88:IRAK4,CpG DNA:TLR9:MYD88:IRAK4:IRAK1,CpG DNA:TLR9:MYD88:IRAK4:IRAK1:TRAF6,CpG DNA:TLR9:MYD88:IRF1,CpG DNA:TLR9:MYD88:TRAF3,DAP12,FcgammaRIIA,FcgammaRIII,HMGB1,IFN-alpha,IFN-beta,IKB-zeta,IKK-alpha,IKK-alpha{active},IL-18,IL-1beta,IL-33,IL12-p35,IRAK-4,IRAK1,IRF-1,IRF-3,IRF-5,IRF-5{active},IRF-7,IRF-7{p},IRF-7{p}{ub},IkBzeta,LL37,MAPK,MyD88,NF-kappaB,NF-kappaB{active},PDC-TREM,PDC-TREM:plexin A1:DAP12,RAGEs receptor,RIG-I,TBK1,TBK1{active},TLR9,TLR9:UNC93B1,TRAF3,TRAF6,UNC93B1,ZBP1,ZBP1 oligomer,ZBP1:IRF3,ZBP1:TBK1,dsRNA,dsRNA:RIG-I,iNOS,plexin A1,proIL-18,proIL-1beta,proIL-33,pyrin,self DNA,self DNA:Ig complex,self DNA:Ig complex:FcgammaRIII,self DNA:Ig complex:FcgammaRIIa,self DNA:Ig complex:TLR9,self DNA:LL37,type I interferon
The nucleotide-binding oligomerization domain (NOD)-like receptor (NLR)(nucleotide-binding domain leucine-rich repeat containing) family of proteinshas been demonstrated to function as regulators of innate immune responseagainst microbial pathogens. Stimulation of NOD1 and NOD2, two prototypic NLRs,results in the activation of MAPK and NF-kappaB. On the other hand, a differentset of NLRs induces caspase-1 activation through the assembly of aninflammasome. This review discusses recent findings regarding the signalingpathways utilized by NLR proteins in the control of caspase-1 and NF-kappaBactivation, as well as the nonredundant role of NLRs in pathogen clearance. Thereview also covers advances regarding the cellular localization of theseproteins and the implications this may have on pathogen sensing and signaltransduction.
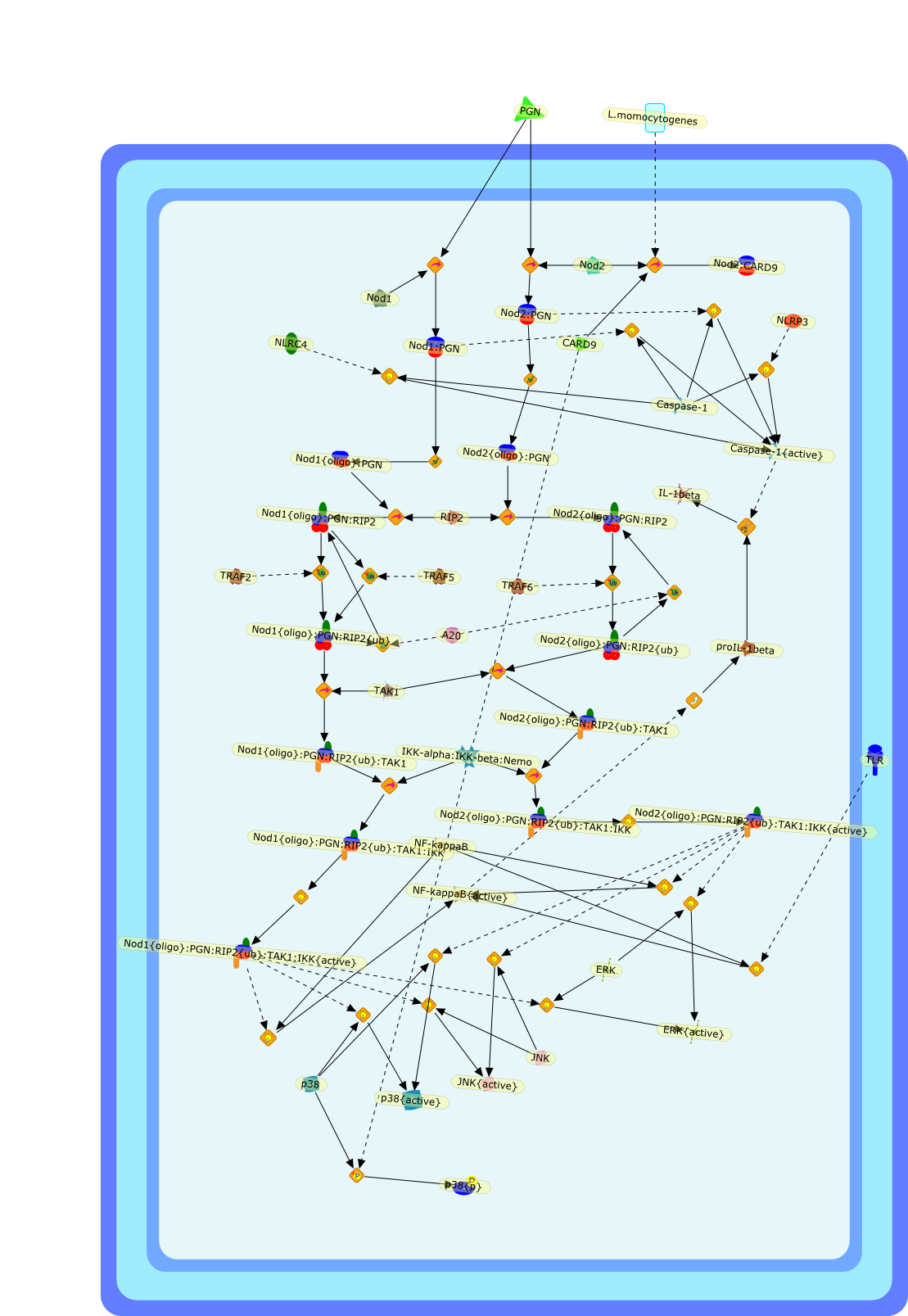{kind=link}
{kind=link}
A20,CARD9,Caspase-1,Caspase-1{active},ERK,ERK{active},IKK-alpha:IKK-beta:Nemo,IL-1beta,JNK,JNK{active},L.momocytogenes,NF-kappaB,NF-kappaB{active},NLRC4,NLRP3,Nod1,Nod1:PGN,Nod1{oligo}:PGN,Nod1{oligo}:PGN:RIP2,Nod1{oligo}:PGN:RIP2{ub},Nod1{oligo}:PGN:RIP2{ub}:TAK1,Nod1{oligo}:PGN:RIP2{ub}:TAK1:IKK,Nod1{oligo}:PGN:RIP2{ub}:TAK1:IKK{active},Nod2,Nod2:CARD9,Nod2:PGN,Nod2{oligo}:PGN,Nod2{oligo}:PGN:RIP2,Nod2{oligo}:PGN:RIP2{ub},Nod2{oligo}:PGN:RIP2{ub}:TAK1,Nod2{oligo}:PGN:RIP2{ub}:TAK1:IKK,Nod2{oligo}:PGN:RIP2{ub}:TAK1:IKK{active},PGN,RIP2,TAK1,TLR,TRAF2,TRAF5,TRAF6,p38,p38{active},p38{p},proIL-1beta
Toll-like receptor (TLR) signaling is subjected to crosstalk from other signals,with a resulting positive or negative effect. There is complex crosstalk betweenthe NLR family of immune-regulatory molecules and TLRs, and C-type lectinreceptors such as Dectin-1 synergize with TLR2 via the tyrosine kinase Syk.Bruton's tyrosine kinase plays an important positive role in TLR signaling,whereas the TAM family of receptor tyrosine kinases is inhibitory. The tyrosinephosphatase SHP1 has been shown to positively regulate induction ofinterferon-beta, whereas SHP2 inhibits the kinase TBK1, limiting this response.K63-linked polyubiquination has also been shown to be critical for theinitiation of TLR signaling. Finally, glucocorticoids affect TLR signaling byinducing the phosphatase MKP1 and inhibiting TBK1 activation. These recentfindings emphasize the importance of considering TLR signaling in the context ofother signaling pathways, as is likely to occur in vivo during infection andinflammation.
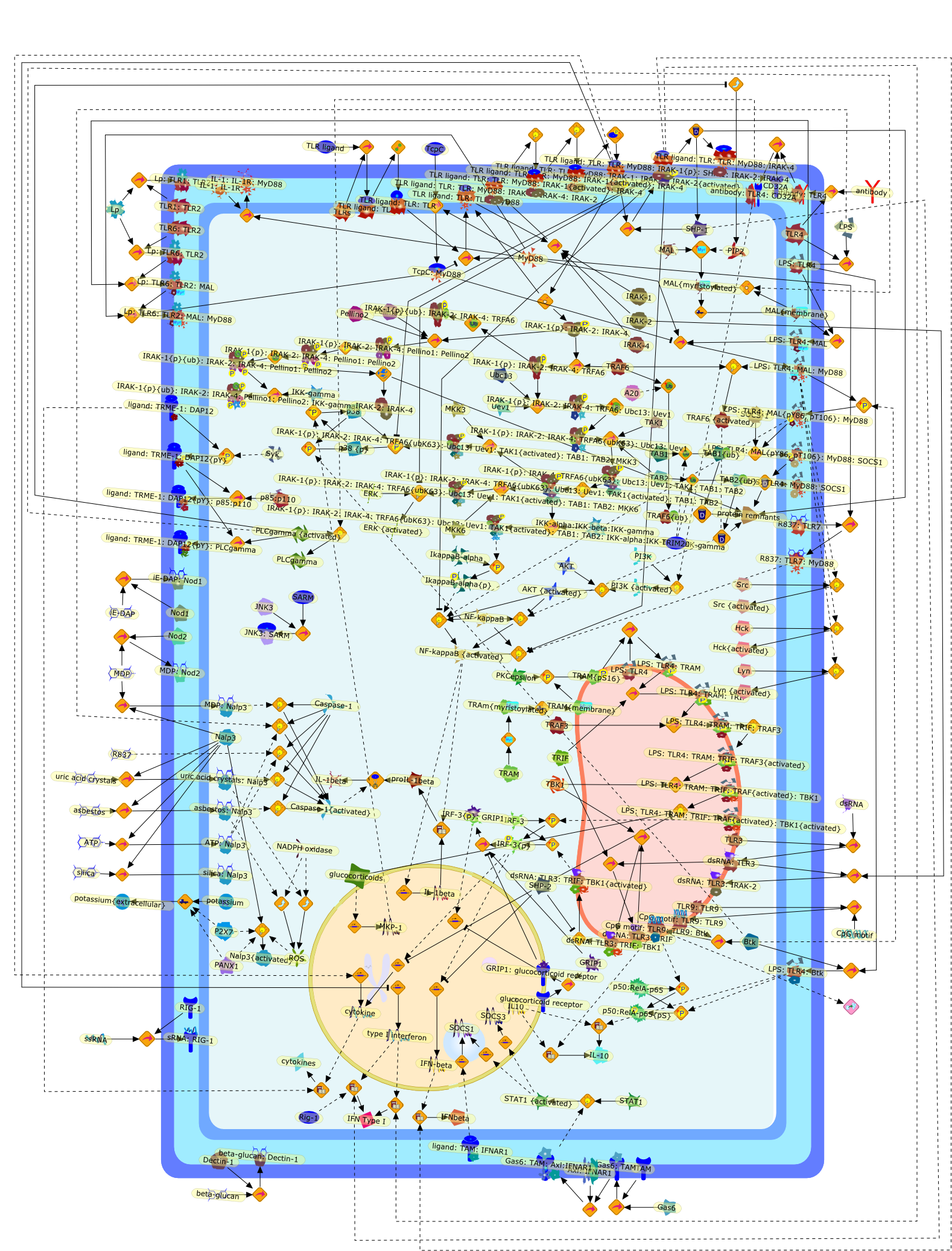{kind=link}
{kind=link}
A20,AKT,AKT {activated},ATP,ATP: Nalp3,Axl: IFNAR1,Btk,CD32A,Caspase-1,Caspase-1{activated},CpG motif,CpG motif: TLR9: TLR9,CpG motif: TLR9: TLR9: Btk,Dectin-1,ERK,ERK {activated},GRIP1,GRIP1: glucocorticoid receptor,Gas6,Gas6: TAM,Gas6: TAM: Axl:IFNAR1,Hck,Hck{activated},IFN Type I,IFN-beta,IFNbeta,IKK-alpha:IKK-beta:IKK-gamma,IKK-gamma,IL-10,IL-1: IL-1R,IL-1: IL-1R: MyD88,IL-1beta,IL10,IRAK-1,IRAK-1{p}: IRAK-2: IRAK-4,IRAK-1{p}: IRAK-2: IRAK-4: Pellino1: Pellino2,IRAK-1{p}: IRAK-2: IRAK-4: TRFA6,IRAK-1{p}: IRAK-2: IRAK-4: TRFA6: Ubc13: Uev1,IRAK-1{p}: IRAK-2: IRAK-4: TRFA6{ubK63}: Ubc13: Uev1,IRAK-1{p}: IRAK-2: IRAK-4: TRFA6{ubK63}: Ubc13: Uev1: TAK1{activated}: TAB1: TAB2,IRAK-1{p}: IRAK-2: IRAK-4: TRFA6{ubK63}: Ubc13: Uev1: TAK1{activated}: TAB1: TAB2: IKK-alpha:IKK-beta:IKK-gamma,IRAK-1{p}: IRAK-2: IRAK-4: TRFA6{ubK63}: Ubc13: Uev1: TAK1{activated}: TAB1: TAB2: MKK3,IRAK-1{p}: IRAK-2: IRAK-4: TRFA6{ubK63}: Ubc13: Uev1: TAK1{activated}: TAB1: TAB2: MKK6,IRAK-1{p}: IRAK-4: TRFA6{ubK63}: Ubc13: Uev1: TAK1: TAB1: TAB2,IRAK-1{p}{ub}: IRAK-2: IRAK-4: Pellino1: Pellino2,IRAK-1{p}{ub}: IRAK-2: IRAK-4: Pellino1: Pellino2: IKK-gamma,IRAK-1{p}{ub}: IRAK-2: IRAK-4: TRFA6,IRAK-2,IRAK-2: IRAK-4,IRAK-4,IRF-3,IRF-3{p},IRF-3{p}: GRIP1,IkappaB-alpha,IkappaB-alpha{p},JNK3,JNK3: SARM,LPS,LPS: TLR4,LPS: TLR4: Btk,LPS: TLR4: MAL,LPS: TLR4: MAL: MyD88,LPS: TLR4: MAL{pY86, pT106}: MyD88,LPS: TLR4: MAL{pY86, pT106}: MyD88: SOCS1,LPS: TLR4: MyD88: SOCS1,LPS: TLR4: TRAM,LPS: TLR4: TRAM: TRIF,LPS: TLR4: TRAM: TRIF: TRAF3,LPS: TLR4: TRAM: TRIF: TRAF3{activated},LPS: TLR4: TRAM: TRIF: TRAF{activated}: TBK1,LPS: TLR4: TRAM: TRIF: TRAF{activated}: TBK1{activated},Lp,Lp: TLR1: TLR2,Lp: TLR6: TLR2,Lp: TLR6: TLR2: MAL,Lp: TLR6: TLR2: MAL: MyD88,Lyn,Lyn {activated},MAL,MAL{membrane},MAL{myristoylated},MDP,MDP: Nalp3,MDP: Nod2,MKK3,MKK6,MKP-1,MyD88,NADPH oxidase,NF-kappaB,NF-kappaB {activated},Nalp3,Nalp3{activated},Nod1,Nod2,P2X7,PANX1,PI3K,PI3K {activated},PIP2,PKCepsilon,PLCgamma,PLCgamma {activated},Pellino1,Pellino2,R837,R837: TLR7,R837: TLR7: MyD88,RIG-1,ROS,Rig-1,SARM,SHP-1,SHP-2,SOCS-1,SOCS1,SOCS3,STAT1,STAT1 {activated},Src,Src {activated},Syk,TAB1,TAB1{ub},TAB2,TAB2{ub},TAK1,TAM,TBK1,TLR ligand,TLR ligand: TLR,TLR ligand: TLR: TLR,TLR ligand: TLR: TLR: MyD88,TLR ligand: TLR: TLR: MyD88: IRAK-1: IRAK-4: IRAK-2,TLR ligand: TLR: TLR: MyD88: IRAK-1: IRAK-4: IRAK-2{activated},TLR ligand: TLR: TLR: MyD88: IRAK-1{activated}: IRAK-4,TLR ligand: TLR: TLR: MyD88: IRAK-1{p}: SHP-1: IRAK-2: IRAK-4,TLR ligand: TLR: TLR: MyD88: IRAK-4,TLR1: TLR2,TLR3,TLR4,TLR6: TLR2,TLR9: TLR9,TLRs,TRAF3,TRAF6,TRAF6 {activated},TRAF6{ub},TRAM,TRAM{membrane},TRAM{pS16},TRAm{myristoylated},TRIF,TRIM20,TcpC,TcpC: MyD88,Ubc13,Uev1,antibody,antibody: TLR4,antibody: TLR4: CD32A,asbestos,asbestos: Nalp3,beta-glucan,beta-glucan: Dectin-1,cytokine,cytokines,dsRNA,dsRNA: TLR3,dsRNA: TLR3: IRAK-2,dsRNA: TLR3: TRIF,dsRNA: TLR3: TRIF: TBK1,dsRNA: TLR3: TRIF: TBK1{activated},glucocorticoid receptor,glucocorticoids,iE-DAP,iE-DAP: Nod1,ligand: TAM: IFNAR1,ligand: TRME-1: DAP12,ligand: TRME-1: DAP12{pY},ligand: TRME-1: DAP12{pY}: PLCgamma,ligand: TRME-1: DAP12{pY}: p85:p110,p38,p38 {p},p50:RelA-p65,p50:RelA-p65{pS},p85:p110,potassium,potassium{extracellular},proIL-1beta,protein remnants,sRNA: RIG-1,silica,silica: Nalp3,ssRNA,type I interferon,uric acid crystals,uric acid crystals: Nalp3
Plasmacytoid dendritic cells (pDCs) are important mediators of antiviralimmunity through their ability to produce large amounts of type I interferons(IFNs) on viral infection. This function of pDCs is linked to their expressionof Toll-like receptor 7 (TLR7) and TLR9, which sense viral nucleic acids withinthe early endosomes. Exclusion of self nucleic acids from TLR-containing earlyendosomes normally prevents pDC responses to them. However, in some autoimmunediseases, self nucleic acids can be modified by host factors and gain entranceto pDC endosomes, where they activate TLR signalling. Several pDC receptorsnegatively regulate type I IFN responses by pDCs during viral infection and fornormal homeostasis.
{kind=link}
{kind=link}
2', 5'--linked oligoadenylate,5'-triphosphate,A type CpG ODN: TLR9,A type CpG ODN: TLR9: MyD88,ADP,BDCA2ligand: BDCA2,BDCA2ligand: BDCA2: IgE: FCepsilonRIgamma,BLNK: BCAP,BLNK: BCAP{activated},Btk,CD40,CD80,CD86,CpG DNA: TLR9,CpG DNA: TLR9: MyD88,CpG DNA: TLR9: MyD88: IRAK-1: IRAK-4: TRAF6: BTK,CpG ODN,CpG ODN: TLR9,CpG ss-DNA ,DAI,DNA-specific IgG antibody,ERK1,ERK1{activated},ERK2,ERK2{activated},FADD: caspase-8,FADD: caspase-8{activated},FCgammaRIIA,FcgammaRIIA,HMGB1,IFN Type I,IFN beta: IFNAR,IFN-alpha1,IFN-beta,IFN-gamma,IFNAR,IFNL,IFNW1,IFNalpha1,IFNbeta,IKK-alpha,IL-12,IL-6,ILT7 ligand: ILT7,ILT7 ligand: ILT7: IgE: FCepsilonRIgamma,IPS1,IRAK,IRAK-1: IRAK4:TRAF6: BTK: IRF-7,IRAK-1: IRAK4:TRAF6: BTK: IRF-7{ub},IRAK-1: IRAK4:TRAF6: BTK{activated},IRAK-1: TRAF3: OPN: IRF-7,IRAK-1: TRAF6: BTK,IRAK-4,IRAK-4: TRAF6,IRAK-4: TRAF6{activated},IRAK1: IRF-7{ub},IRAK1: IRF-7{ub}: TRAF3: TABK1: IKK-alpha: OPN: PI3K,IRAK4:TRAF6: BTK,IRF-3,IRF-3{p},IRF-4,IRF-5,IRF-5{activated},IRF-7,IRF-7{p},IRF-7{ub}{p},IRF-8,IgE: FCepsilonRIgamma,LGP2,LL37,LL37: self DNA,LL37: self DNA: HMGB1,LL37: self DNA: HMGB1: DNA-specific IgG antibody,LL37: self DNA: HMGB1: DNA-specific IgG antibody: FCgammaRIIA,LL37: self DNA: HMGB1: DNA-specific IgG antibody: TLR9,LL37: self DNA: HMGB1: RAGE,LL37: self DNA: HMGB1: TLR9,LL37: self DNA: HMGB1: TLR9: MyD88,LPS,LPS: TLR4,LPS: TLR4: MAL,LPS: TLR4: MAL: MyD88,LPS: TLR4: TRAM,LPS: TLR4: TRAM: TRIF,Lp,Lp: TLR1: TLR2,Lp: TLR1: TLR2: MAL,Lp: TLR1: TLR2: MAL: MyD88,MAL,MAPKs,MAPKs{activated},MDA-5,MHC,MyD88,MyD88: IRF-4,MyD88: IRF-5,OPN,P2Y,PDC-TREM,PDC-TREM: plexin A1,PGN,PGN: TLR6: TLR2,PI3K,PI3K{activated},PKR,PKR{activated},PTKs,PTKs{activated},Poly I:C: TLR3,R-848,R-848: TLR7,R-848: TLR8,RAGE,RIG-1,RNASEL,RNASEL{activated},Syk,Syk{activated},TAK1,TAK1{activated},TBK1,TBK1{activated},TLR1: TLR2,TLR3,TLR3: UNC93B: gp96,TLR4,TLR5,TLR6: TLR2,TLR7,TLR7: UNC93B: gp96,TLR8,TLR9,TLR9: UNC93B: gp96,TNF-alpha,TRAF3,TRAF3: TBK1: IKK-i,TRAF3: TBK1: IKK-i{activated},TRAF6,TRAM,TRIF,Type B CpG ODN: TLR9,Type B CpG ODN: TLR9: MyD88,UNC93B,antibody,auto-antibody,dsRNA,dsRNA: DAI,dsRNA: LGP2,dsRNA: LGP2: IPS1,dsRNA: MDA-5,dsRNA: MDA-5: IPS1,dsRNA: RIG-1,dsRNA: RIG-1: IPS1,dsRNA: RIG1: IPS1: TRAF3: TBK1: IKK-i,dsRNA:TLR3,dsRNA:TLR3:TRIF,flagellin,flagellin: TLR5,gp96,guanosine analogues,guanosine analogues: TLR7,guanosine analogues: TLR8,imidazoquinoline: TLR7,imidazoquinoline: TLR8,imiquimod,ligand: BCR,ligand: BCR{activated},ligand: DAP12,ligand: NKp44,ligand: NKp44: ligand: DAP12,ligand: Siglec-H,ligand: Siglec-H: ligand: DAP12,nucleortide: P2Y,p50:RelA-p65:IkappaB-alpha,p50:RelA-p65:IkappaB-alpha{p},p50:RelA-p65{activated},plexin A1,poly I:C: TLR3,self DNA,self RNA,self RNA: TLR7,semaphorin 6D,semaphorin 6D: PDC-TREM: plexin A1,small RNA cleavage products,snRNA,snRNA: TLR7,ssRNA,ssRNA: TLR7,ssRNA: TLR7: MyD88,ssRNA: TLR7: MyD88: IRAK-1: IRAK-4: TRAF6: BTK,ssRNA: TLR8,ssRNA: TLR8: MyD88,ssRNA: TLR8: MyD88: IRAK-1: IRAK-4: TRAF6: BTK
Inflammation underlies a wide variety of physiological and pathologicalprocesses. Although the pathological aspects of many types of inflammation arewell appreciated, their physiological functions are mostly unknown. The classicinstigators of inflammation - infection and tissue injury - are at one end of alarge range of adverse conditions that induce inflammation, and they trigger therecruitment of leukocytes and plasma proteins to the affected tissue site.Tissue stress or malfunction similarly induces an adaptive response, which isreferred to here as para-inflammation. This response relies mainly ontissue-resident macrophages and is intermediate between the basal homeostaticstate and a classic inflammatory response. Para-inflammation is probablyresponsible for the chronic inflammatory conditions that are associated withmodern human diseases.
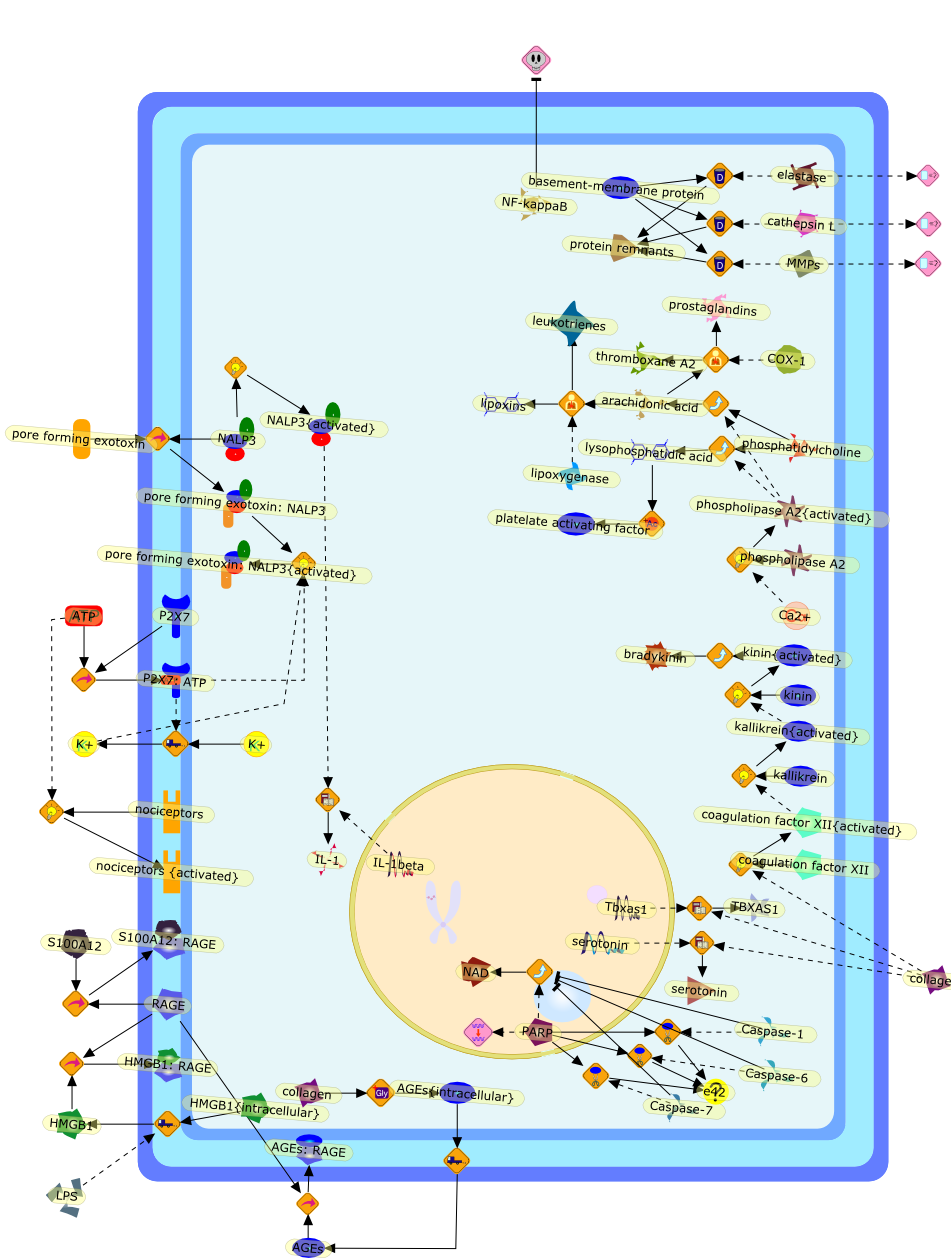{kind=link}
{kind=link}
AGEs,AGEs: RAGE,AGEs{intracellular},ATP,COX-1,Ca2+,Caspase-1,Caspase-6,Caspase-7,HMGB1,HMGB1: RAGE,HMGB1{intracellular},IL-1,IL-1beta,K+,LPS,MMPs,NAD,NALP3,NALP3{activated},NF-kappaB,P2X7,P2X7: ATP,PARP,RAGE,S100A12,S100A12: RAGE,TBXAS1,Tbxas1,arachidonic acid,basement-membrane protein,bradykinin,cathepsin L,coagulation factor XII,coagulation factor XII{activated},collagen,e42,elastase,kallikrein,kallikrein{activated},kinin,kinin{activated},leukotrienes,lipoxins,lipoxygenase,lysophosphatidic acid,nociceptors,nociceptors {activated},phosphatidylcholine,phospholipase A2,phospholipase A2{activated},platelate activating factor,pore forming exotoxin,pore forming exotoxin: NALP3,pore forming exotoxin: NALP3{activated},prostaglandins,protein remnants,serotonin,thromboxane A2
In viral infections the host innate immune system is meant to act as a firstline defense to prevent viral invasion or replication before more specificprotection by the adaptive immune system is generated. In the innate immuneresponse, pattern recognition receptors (PRRs) are engaged to detect specificviral components such as viral RNA or DNA or viral intermediate products and toinduce type I interferons (IFNs) and other pro-inflammatory cytokines in theinfected cells and other immune cells. Recently these innate immune receptorsand their unique downstream pathways have been identified. Here, we summarizetheir roles in the innate immune response to virus infection, discriminationbetween self and viral nucleic acids and inhibition by virulent factors andprovide some recent advances in the coordination between innate and adaptiveimmune activation.
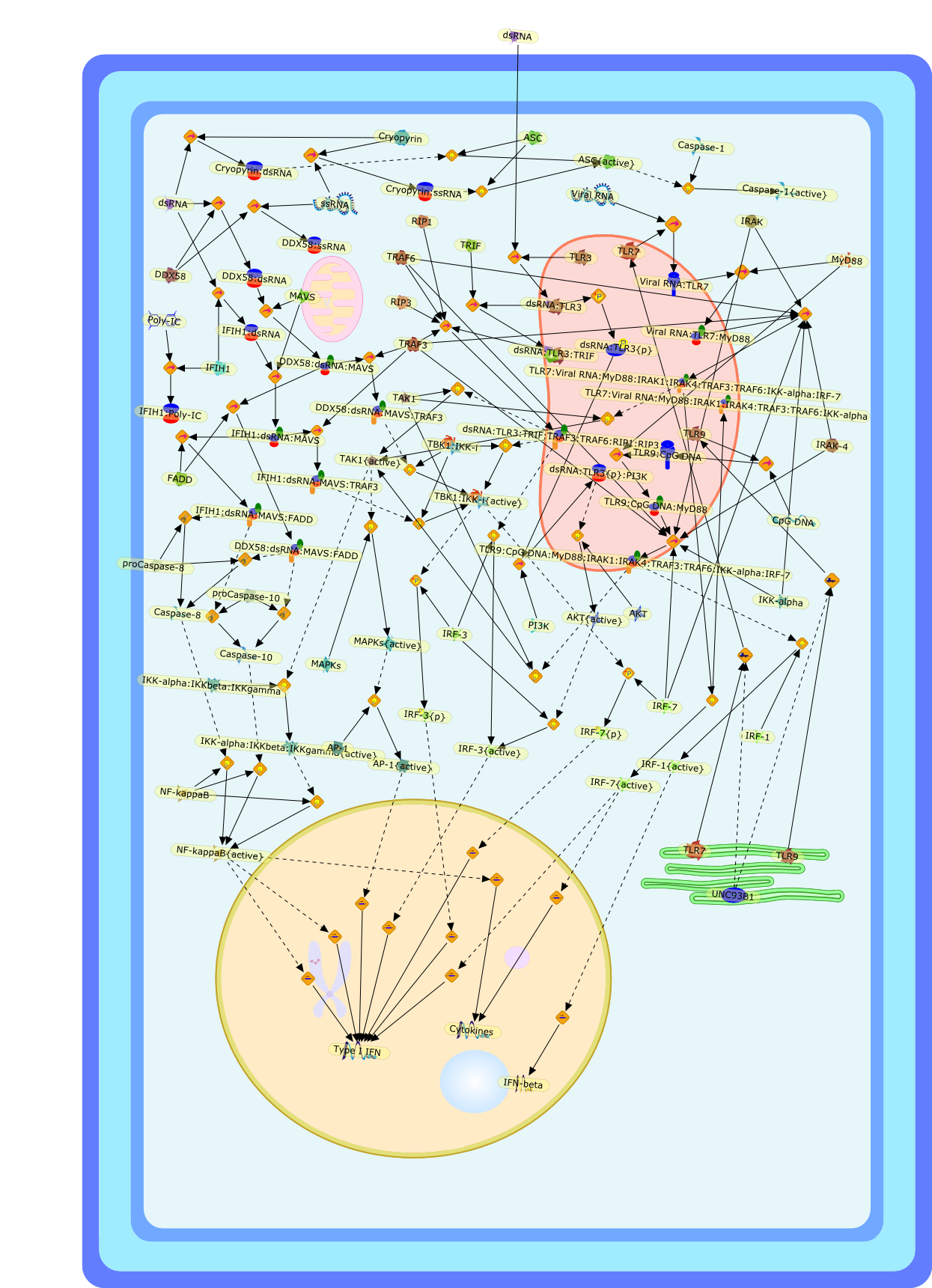{kind=link}
{kind=link}
AKT,AKT{active},AP-1,AP-1{active},ASC,ASC{active},Caspase-1,Caspase-10,Caspase-1{active},Caspase-8,CpG DNA,Cryopyrin,Cryopyrin:dsRNA,Cryopyrin:ssRNA,Cytokines,DDX58,DDX58:dsRNA,DDX58:dsRNA:MAVS,DDX58:dsRNA:MAVS:FADD,DDX58:dsRNA:MAVS:TRAF3,DDX58:ssRNA,FADD,IFIH1,IFIH1:Poly-IC,IFIH1:dsRNA,IFIH1:dsRNA:MAVS,IFIH1:dsRNA:MAVS:FADD,IFIH1:dsRNA:MAVS:TRAF3,IFN-beta,IKK-alpha,IKK-alpha:IKKbeta:IKKgamma,IKK-alpha:IKKbeta:IKKgamma{active},IRAK,IRAK-4,IRF-1,IRF-1{active},IRF-3,IRF-3{active},IRF-3{p},IRF-7,IRF-7{active},IRF-7{p},MAPKs,MAPKs{active},MAVS,MyD88,NF-kappaB,NF-kappaB{active},PI3K,Poly-IC,RIP1,RIP3,TAK1,TAK1{active},TBK1:IKK-i,TBK1:IKK-i{active},TLR3,TLR7,TLR7:Viral RNA:MyD88:IRAK1:IRAK4:TRAF3:TRAF6:IKK-alpha,TLR7:Viral RNA:MyD88:IRAK1:IRAK4:TRAF3:TRAF6:IKK-alpha:IRF-7,TLR9,TLR9:CpG DNA,TLR9:CpG DNA:MyD88,TLR9:CpG DNA:MyD88:IRAK1:IRAK4:TRAF3:TRAF6:IKK-alpha:IRF-7,TRAF3,TRAF6,TRIF,Type I IFN ,UNC93B1,Viral RNA,Viral RNA:TLR7,Viral RNA:TLR7:MyD88,dsRNA,dsRNA:TLR3,dsRNA:TLR3:TRIF,dsRNA:TLR3:TRIF:TRAF3:TRAF6:RIP1:RIP3,dsRNA:TLR3{p},dsRNA:TLR3{p}:PI3K,proCaspase-10,proCaspase-8,ssRNA
The recent, rapid progress in our understanding of cytoplasmic RNA-mediatedantiviral innate immune signaling was initiated by the discovery of retinoicacid-inducible gene I (RIG-I) as a sensor of viral RNA. It is now widelyrecognized that RIG-I and related RNA helicases, melanomadifferentiation-associated gene-5 (MDA5) and laboratory of genetics andphysiology-2 (LGP2), can initiate and/or regulate RNA and virus-mediated type IIFN production and antiviral responses. As with other cytokine systems,production of type I IFN is a transient process, and can be hazardous to thehost if unregulated, resulting in chronic cellular toxicity or inflammatory andautoimmune diseases. In addition, the RIG-I-like receptor (RLR) system is afundamental target for virus-encoded immune suppression, with many indirect anddirect examples of interference described. In this article, we review thecurrent understanding of endogenous negative regulation in RLR signaling andexplore direct inhibition of RLR signaling by viruses as a host immune evasionstrategy.
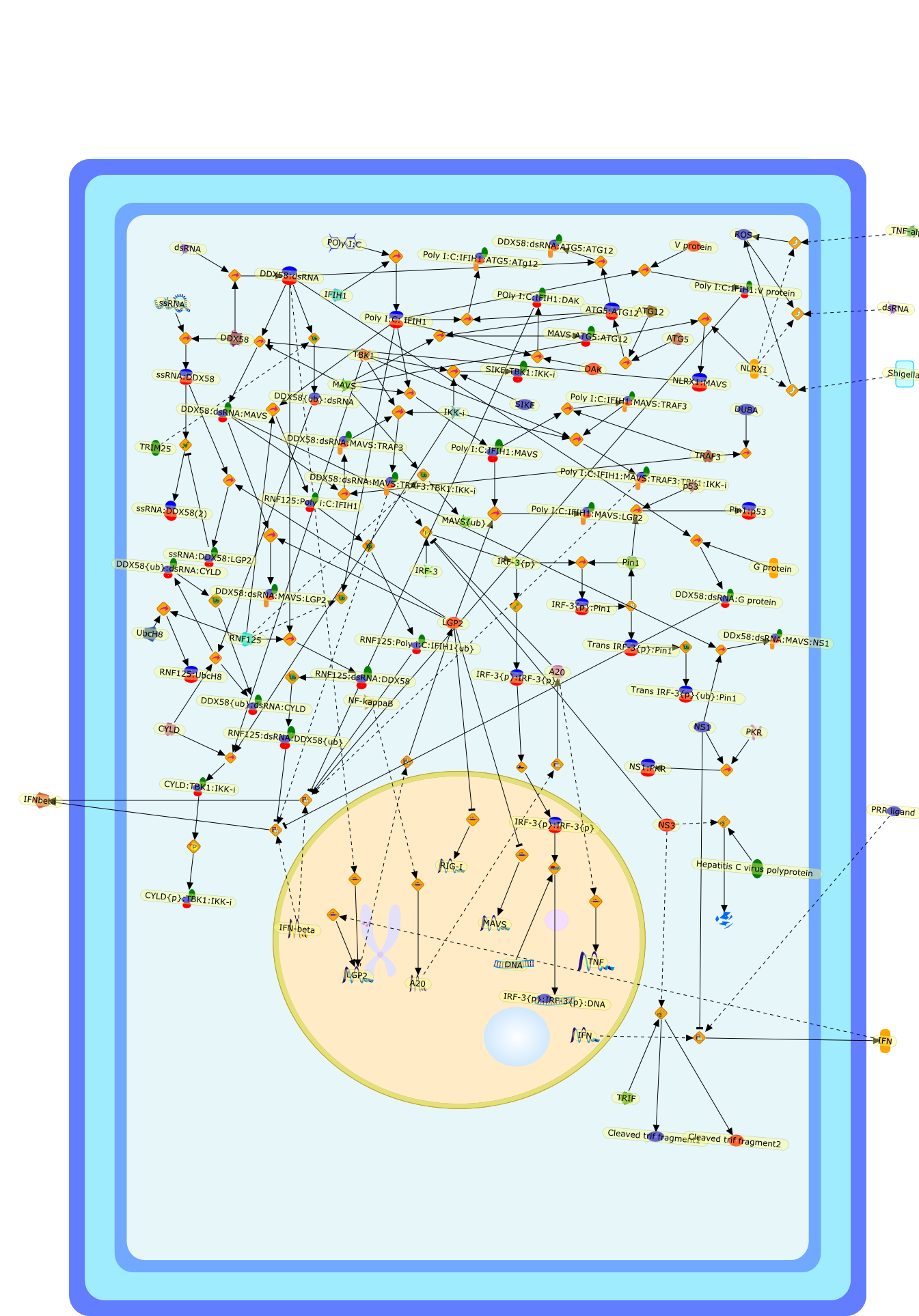{kind=link}
{kind=link}
A20,ATG12,ATG5,ATG5:ATG12,CYLD,CYLD:TBK1:IKK-i,CYLD{p}:TBK1:IKK-i,Cleaved fragments,Cleaved trif fragment1,Cleaved trif fragment2,DAK,DDX58,DDX58:dsRNA,DDX58:dsRNA:ATG5:ATG12,DDX58:dsRNA:G protein,DDX58:dsRNA:MAVS,DDX58:dsRNA:MAVS:LGP2,DDX58:dsRNA:MAVS:TRAF3,DDX58:dsRNA:MAVS:TRAF3:TBK1:IKK-i,DDX58{ub}:dsRNA,DDX58{ub}:dsRNA:CYLD,DDx58:dsRNA:MAVS:NS1,DNA,DUBA,G protein,Hepatitis C virus polyprotein,IFIH1,IFN,IFN-beta,IFNbeta,IKK-i,IRF-3,IRF-3{p},IRF-3{p}:IRF-3{p},IRF-3{p}:IRF-3{p}:DNA,IRF-3{p}:Pin1,LGP2,MAVS,MAVS:ATG5:ATG12,MAVS{ub},NF-kappaB,NLRX1,NLRX1:MAVS,NS1,NS1:PKR,NS3,PKR,POly I:C,POly I:C:IFIH1:DAK,PRR ligand,Pin1,Pin1:p53,Poly I:C: IFIH1,Poly I:C:IFIH1:ATG5:ATg12,Poly I:C:IFIH1:MAVS,Poly I:C:IFIH1:MAVS:LGP2,Poly I:C:IFIH1:MAVS:TRAF3,Poly I:C:IFIH1:MAVS:TRAF3:TBK1:IKK-i,Poly I:C:IFIH1:V protein,RIG-I,RNF125,RNF125:Poly i:C:IFIH1,RNF125:Poly i:C:IFIH1{ub},RNF125:UbcH8,RNF125:dsRNA:DDX58,RNF125:dsRNA:DDX58{ub},ROS,SIKE,SIKE:TBK1:IKK-i,Shigella,TBK1,TNF,TNF-alpha,TRAF3,TRIF,TRIM25,Trans IRF-3{p}:Pin1,Trans IRF-3{p}{ub}:Pin1,UbcH8,V protein,dsRNA,p53,ssRNA,ssRNA:DDX58,ssRNA:DDX58(2),ssRNA:DDX58:LGP2
Research into the five Toll/IL1 receptor (TIR) adaptor proteins involved ininnate immunity continues to advance. Here we outline some of the more recentfindings. MyD88 has a key role in signalling by the IL1 receptor complex andTLRs. However, a MyD88-independent pathway of IL1beta signalling in neurons hasbeen described which involves the protein kinase Akt, and which has ananti-apoptotic effect. This pathway may also be important for the mechanismwhereby Alum exerts its adjuvant effect since this depends on IL1beta but isMyD88-independent. MyD88 is also involved in tumourigenesis in models ofhepatocarcinoma and familial associated polyposis (FAP); negative regulation ofTLR3 signalling and in PKCepsilon activation. The adaptor Mal is regulated byphosphorylation and caspase-1 cleavage. A variant form of Mal in humans termedS180L confers protection in multiple infectious diseases. TRAM is controlled bymyristoylation and phosphorylation and the localisation of TRAM with TLR4 toendosomes is required for activation of IRF3 and induction of IFNbeta. FinallySARM has been shown to regulate TRIF and also appears to be involved in neuronalinjury mediated by oxidative stress in mouse neurons. These advances confirm theimportance for the TIR domain-containing adapters in host defence andinflammation.
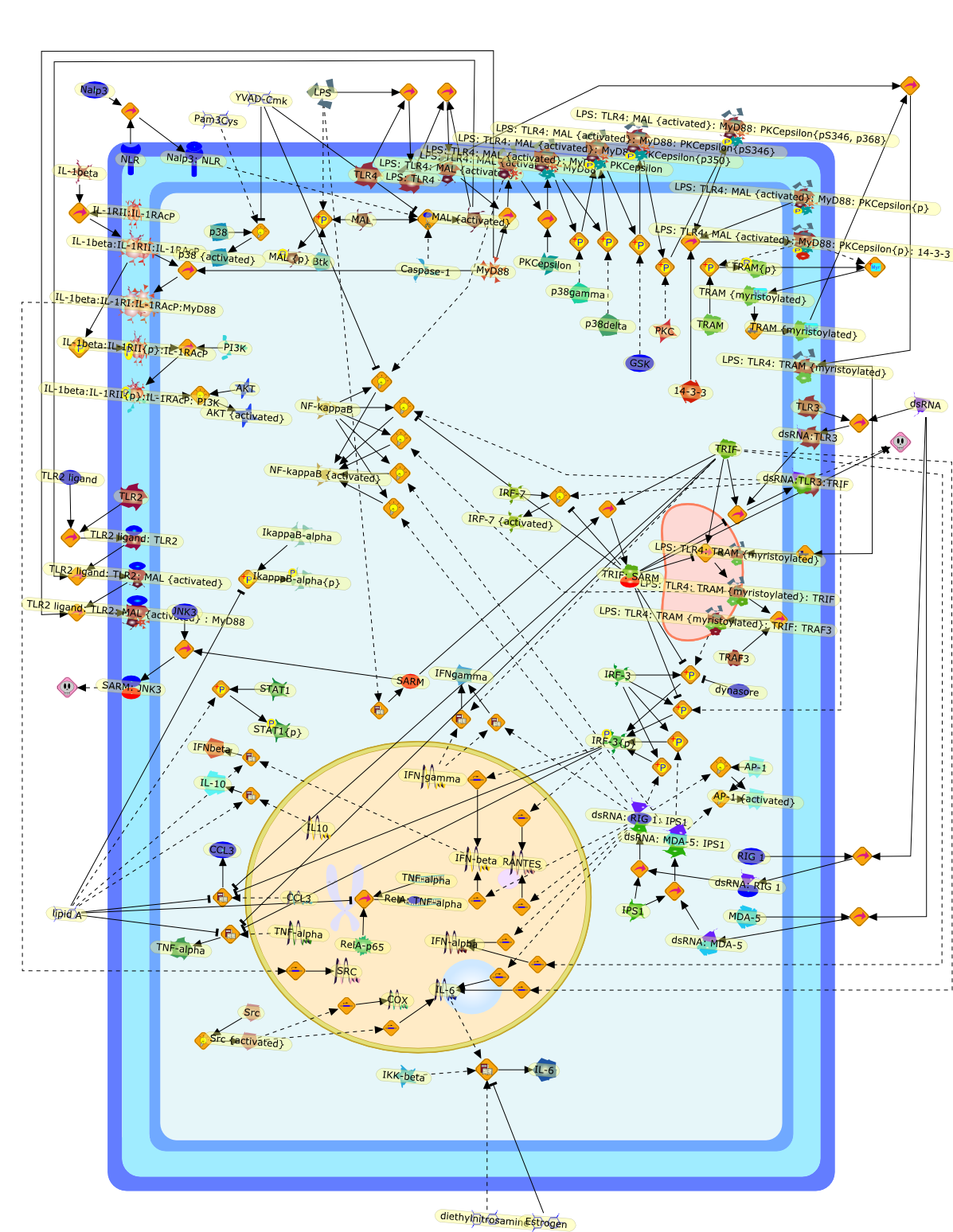{kind=link}
{kind=link}
14-3-3,AKT,AKT {activated},AP-1,AP-1 {activated},Btk,CCL3,COX,Caspase-1,Estrogen,GSK,IFN-alpha,IFN-beta,IFN-gamma,IFNbeta,IFNgamma,IKK-beta,IL-10,IL-1RII:IL-1RAcP,IL-1beta,IL-1beta:IL-1RI:IL-1RAcP:MyD88,IL-1beta:IL-1RII:IL-1RAcP,IL-1beta:IL-1RII{p}:IL-1RAcP,IL-1beta:IL-1RII{p}:IL-1RAcP: PI3K,IL-6,IL10,IPS1,IRF-3,IRF-3{p},IRF-7,IRF-7 {activated},IkappaB-alpha,IkappaB-alpha{p},JNK3,LPS,LPS: TLR4,LPS: TLR4: MAL {activated},LPS: TLR4: MAL {activated}: MyD88,LPS: TLR4: MAL {activated}: MyD88: PKCepsilon,LPS: TLR4: MAL {activated}: MyD88: PKCepsilon{p350},LPS: TLR4: MAL {activated}: MyD88: PKCepsilon{pS346, p368},LPS: TLR4: MAL {activated}: MyD88: PKCepsilon{pS346},LPS: TLR4: MAL {activated}: MyD88: PKCepsilon{p},LPS: TLR4: MAL {activated}: MyD88: PKCepsilon{p}: 14-3-3,LPS: TLR4: TRAM {myristoylated},LPS: TLR4: TRAM {myristoylated}: TRIF,LPS: TLR4: TRAM {myristoylated}: TRIF: TRAF3,MAL,MAL {activated},MAL {p},MDA-5,MyD88,NF-kappaB,NF-kappaB {activated},NLR,Nalp3,Nalp3: NLR,PI3K,PKC,PKCepsilon,Pam3Cys,RANTES,RIG 1,RelA-p65,RelA: TNF-alpha,SARM,SARM: JNK3,SRC,STAT1,STAT1{p},Src,Src {activated},TLR2,TLR2 ligand,TLR2 ligand: TLR2,TLR2 ligand: TLR2: MAL {activated},TLR2 ligand: TLR2: MAL {activated} : MyD88,TLR3,TLR4,TNF-alpha,TRAF3,TRAM,TRAM {myristoylated},TRAM{p},TRIF,TRIF: SARM,YVAD-Cmk,diethylnitrosamine,dsRNA,dsRNA: MDA-5,dsRNA: MDA-5: IPS1,dsRNA: RIG 1,dsRNA: RIG 1: IPS1,dsRNA:TLR3,dsRNA:TLR3:TRIF,dynasore,lipid A,p38,p38 {activated},p38delta,p38gamma
NF-kappaB (nuclear factor-kappaB) transcription factors have multiple criticalroles in the regulation of immune responses. In unstimulated cells, NF-kappaBproteins are sequestered in the cytoplasm by IkappaB inhibitory proteins.Various immune stimuli induce the IkappaB kinase (IKK) to phosphorylateIkappaBs, triggering their ubiquitination and proteasomal degradation, whichpermits nuclear translocation of associated NF-kappaB subunits and activation ofNF-kappaB target genes. Recent studies have highlighted the importance ofdynamic ubiquitination-deubiquitination events in regulating this canonicalNF-kappaB signaling pathway. Ubiquitination additionally plays critical roles inactivation of the noncanonical pathway that regulates NF-kappaB viasignal-induced processing of NF-kappaB2 p100. New research has also identifiedseveral novel regulatory proteins that control the transcriptional activity ofnuclear NF-kappaB.
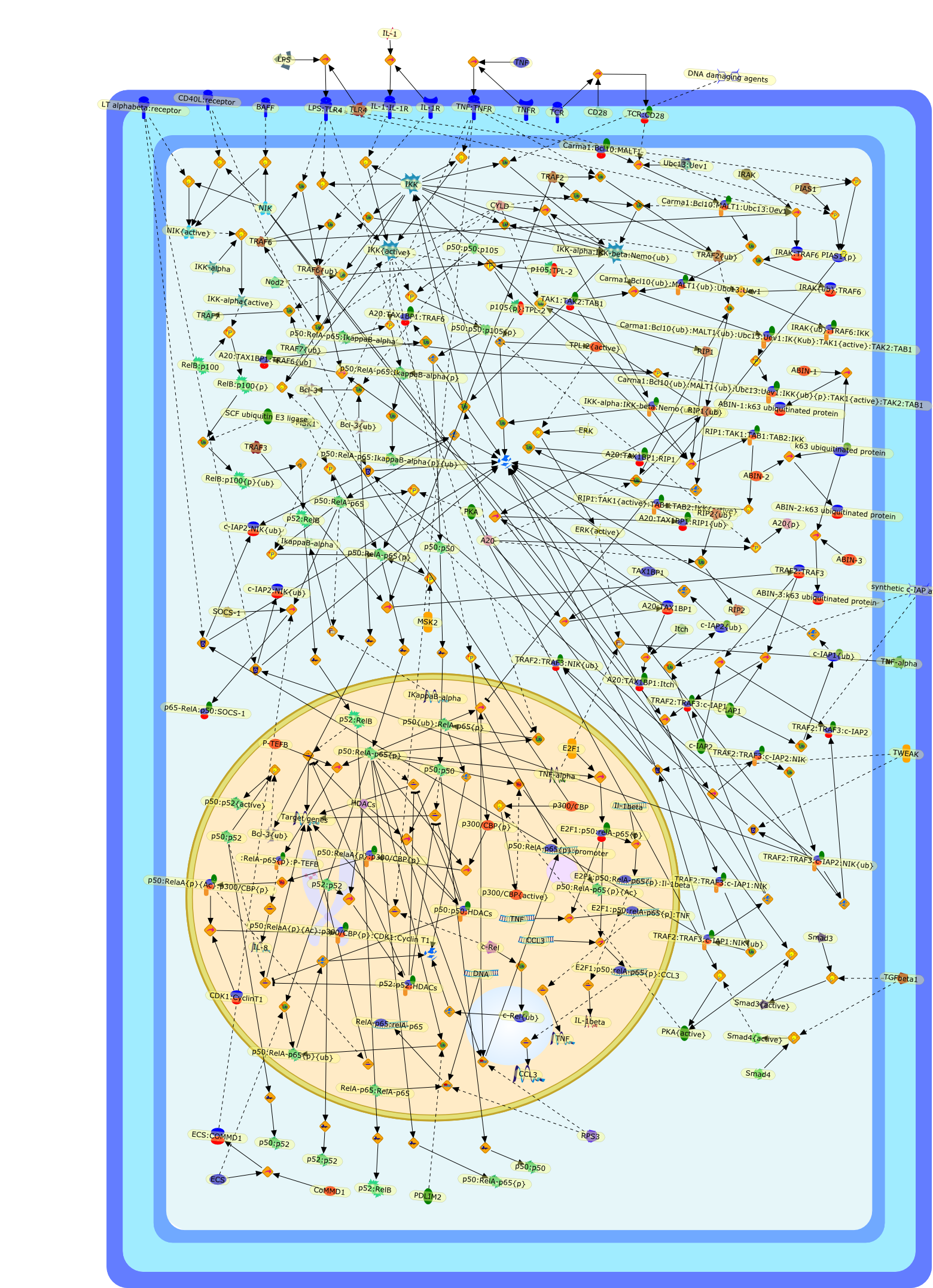{kind=link}
{kind=link}
:RelA-p65{p}:P-TEFB,A20,A20:TAX1BP1,A20:TAX1BP1:Itch,A20:TAX1BP1:RIP1,A20:TAX1BP1:RIP1{ub},A20:TAX1BP1:TRAF6,A20:TAX1BP1:TRAF6{ub],A20{p},ABIN-1,ABIN-1:k63 ubiquitinated protein,ABIN-2,ABIN-2:k63 ubiquitinated protein,ABIN-3,ABIN-3:k63 ubiquitinated protein,BAFF,Bcl-3,Bcl-3{ub},CCL3,CD28,CD40L:receptor,CDK1:CyclinT1,CYLD,Carma1:Bcl10:MALT1,Carma1:Bcl10:MALT1:Ubc13:Uev1,Carma1:Bcl10{ub}:MALT1{ub}:Ubc13:Uev1,Carma1:Bcl10{ub}:MALT1{ub}:Ubc13:Uev1:IKK{ub}{p}:TAK1{active}:TAK2:TAB1,Carma1:Bcl10{ub}:MALT1{ub}:Ubc13:Uev1:IK{Kub}:TAK1{active}:TAK2:TAB1,CoMMD1,DNA,DNA damaging agents,E2F1,E2F1:p50:RelA-p65{p}:Il-1beta,E2F1:p50:relA-p65{p]:TNF,E2F1:p50:relA-p65{p},E2F1:p50:relA-p65{p}:CCL3,ECS,ECS:COMMD1,ERK,ERK{active},HDACs,IKK,IKK-alpha,IKK-alpha:IKK-beta:Nemo{ub},IKK-alpha:IKK-beta:Nemo{ub}:CYLD,IKK-alpha{active},IKK{active},IKappaB-alpha,IL-1,IL-1:IL-1R,IL-1R,IL-1beta,IL-8,IRAK,IRAK:TRAF6,IRAK{ub}:TRAF6,IRAK{ub}:TRAF6:IKK,IkappaB-alpha,Il-1beta,Itch,LPS,LPS:TLR4,LT alphabeta:receptor,MSK1,MSK2,NIK,NIK{active},Nod2,P-TEFB,PDLIM2,PIAS1,PIAS1{p},PKA,PKA{active},Proteosome degradants,RIP1,RIP1:TAK1:TAB1:TAB2:IKK,RIP1:TAK1{active}:TAB1:TAB2:IKK{active},RIP1{ub},RIP2,RIP2{ub},RPS3,RelA-p65:RelA-p65,RelA-p65:relA-p65,RelB:p100,RelB:p100{p},RelB:p100{p}{ub},SCF ubiquitin E3 ligase,SOCS-1,Smad3,Smad3{active},Smad4,Smad4{active},TAK1:TAK2:TAB1,TAX1BP1,TCR,TCR:CD28,TGFbeta1,TLR4,TNF,TNF-alpha,TNF:TNFR,TNFR,TPL-2{active},TRAF2,TRAF2:TRAF3,TRAF2:TRAF3:NIK{ub},TRAF2:TRAF3:c-IAP1,TRAF2:TRAF3:c-IAP1:NIK,TRAF2:TRAF3:c-IAP1:NIK{ub},TRAF2:TRAF3:c-IAP2,TRAF2:TRAF3:c-IAP2:NIK,TRAF2:TRAF3:c-IAP2:NIK{ub},TRAF2{ub},TRAF3,TRAF6,TRAF6{ub},TRAF7,TRAF7{ub},TWEAK,Target genes,Ubc13:Uev1,c-IAP1,c-IAP1{ub},c-IAP2,c-IAP2:NIK{ub},c-IAP2{ub},c-Rel,c-Rel{ub},k63 ubiquitinated protein,p105:TPL-2,p105{p}:TPL-2,p300/CBP,p300/CBP{active},p300/CBP{p},p50:RelA-p65,p50:RelA-p65:IkappaB-alpha,p50:RelA-p65:IkappaB-alpha{p},p50:RelA-p65:IkappaB-alpha{p}{ub},p50:RelA-p65{p},p50:RelA-p65{p}:promoter,p50:RelA-p65{p}{Ac},p50:RelA-p65{p}{ub},p50:RelaA{p}:p300/CBP{p},p50:RelaA{p}{Ac}:p300/CBP{p},p50:RelaA{p}{Ac}:p300/CBP{p}:CDK1:Cyclin T1,p50:p50,p50:p50:HDACs,p50:p50:p105,p50:p50:p105{p},p50:p52,p50:p52{active},p50{ub}:RelA-p65{p},p52:RelB,p52:p52,p52:p52:HDACs,p65-RelA:p50:SOCS-1,proteosome degradants,synthetic c-IAP antagonist
The production of type I interferon -- the first line of defense against virusinfection and critical for innate immunity -- in plasmacytoid dendritic cellsrelies on the mammalian target of rapamycin.
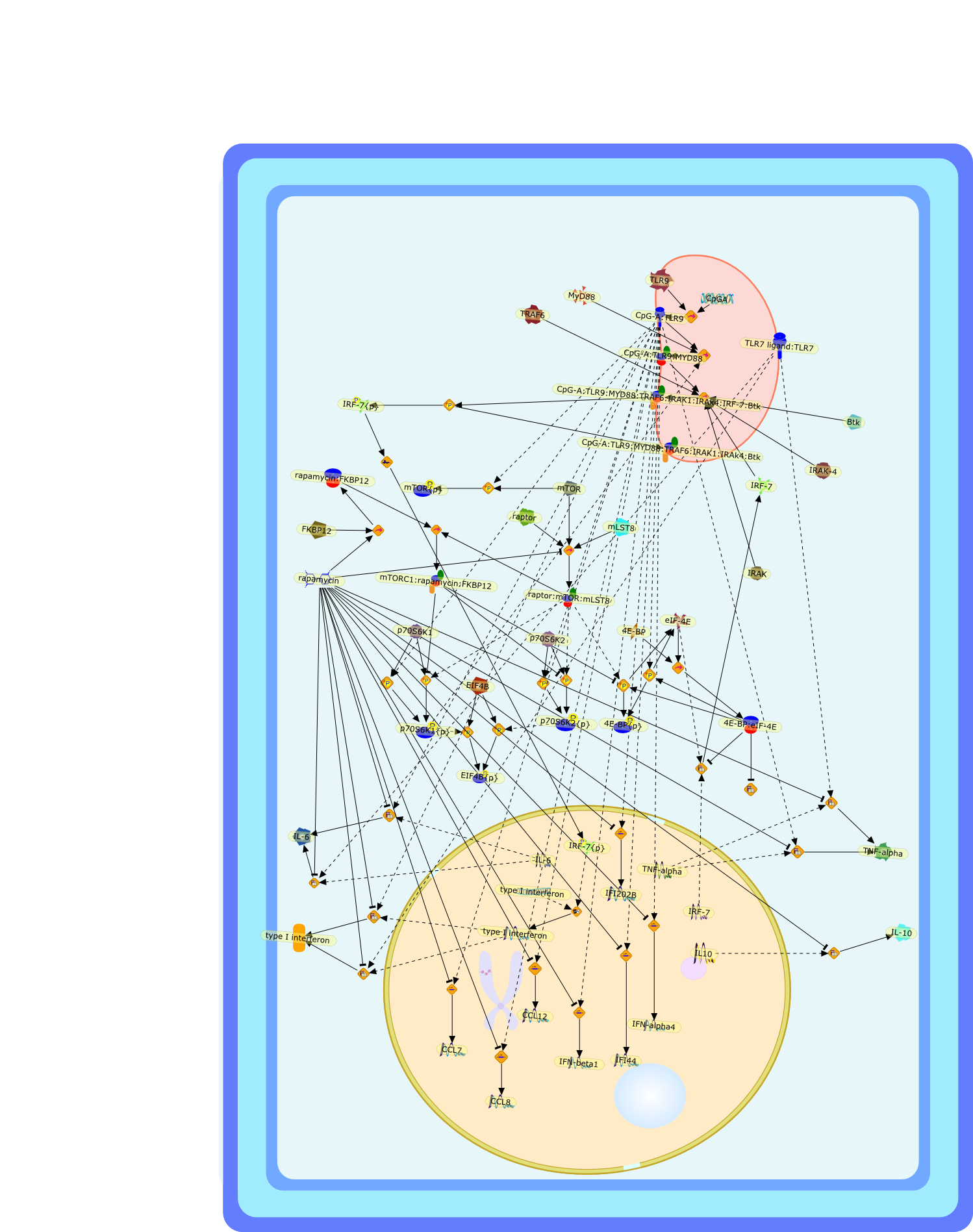{kind=link}
{kind=link}
4E-BP,4E-BP:eIF-4E,4E-BP{p},Btk,CCL12,CCL7,CCL8,CpG-A:TLR9,CpG-A:TLR9:MYD88,CpG-A:TLR9:MYD88:TRAF6:IRAK1:IRAk4:Btk,CpG-A:TLR9:MYD88:TRAF6:IRAK1:IRAk4:IRF-7:Btk,CpGA,EIF4B,EIF4B{p},FKBP12,IFI202B,IFI44,IFN-alpha4,IFN-beta1,IL-10,IL-6,IL10,IRAK,IRAK-4,IRF-7,IRF-7{p},MyD88,TLR7 ligand:TLR7,TLR9,TNF-alpha,TRAF6,eIF-4E,mLST8,mTOR,mTORC1:rapamycin:FKBP12,mTOR{p},p70S6K1,p70S6K1{p},p70S6K2,p70S6K2{p},rapamycin,rapamycin:FKBP12,raptor,raptor:mTOR:mLST8,type I interferon
Innate immune responses mediated by Toll-like receptors (TLRs), a class ofpattern-recognition receptors, play a critical role in the defense againstmicrobial pathogens. However, excessive TLR-mediated responses result in sepsis,autoimmunity, and chronic inflammation. To prevent deleterious activation ofTLRs, cells have evolved multiple mechanisms that inhibit innate immunereactions. Stimulation of TLRs induces the expression of the gene encoding themitogen-activated protein kinase (MAPK) phosphatase-1 (MKP-1), anuclear-localized dual-specificity phosphatase that preferentiallydephosphorylates p38 MAPK and c-Jun N-terminal kinase (JNK), resulting in theattenuation of TLR-triggered production of proinflammatory cytokines. MKP-1 isposttranslationally modified by multiple mechanisms, including phosphorylation.A study now demonstrates that MKP-1 is also acetylated on a key lysine residuefollowing stimulation of TLRs. Acetylation of MKP-1 promotes the interaction ofMKP-1 with its substrate p38 MAPK, which results in dephosphorylation of p38MAPK and the inhibition of innate immunity.
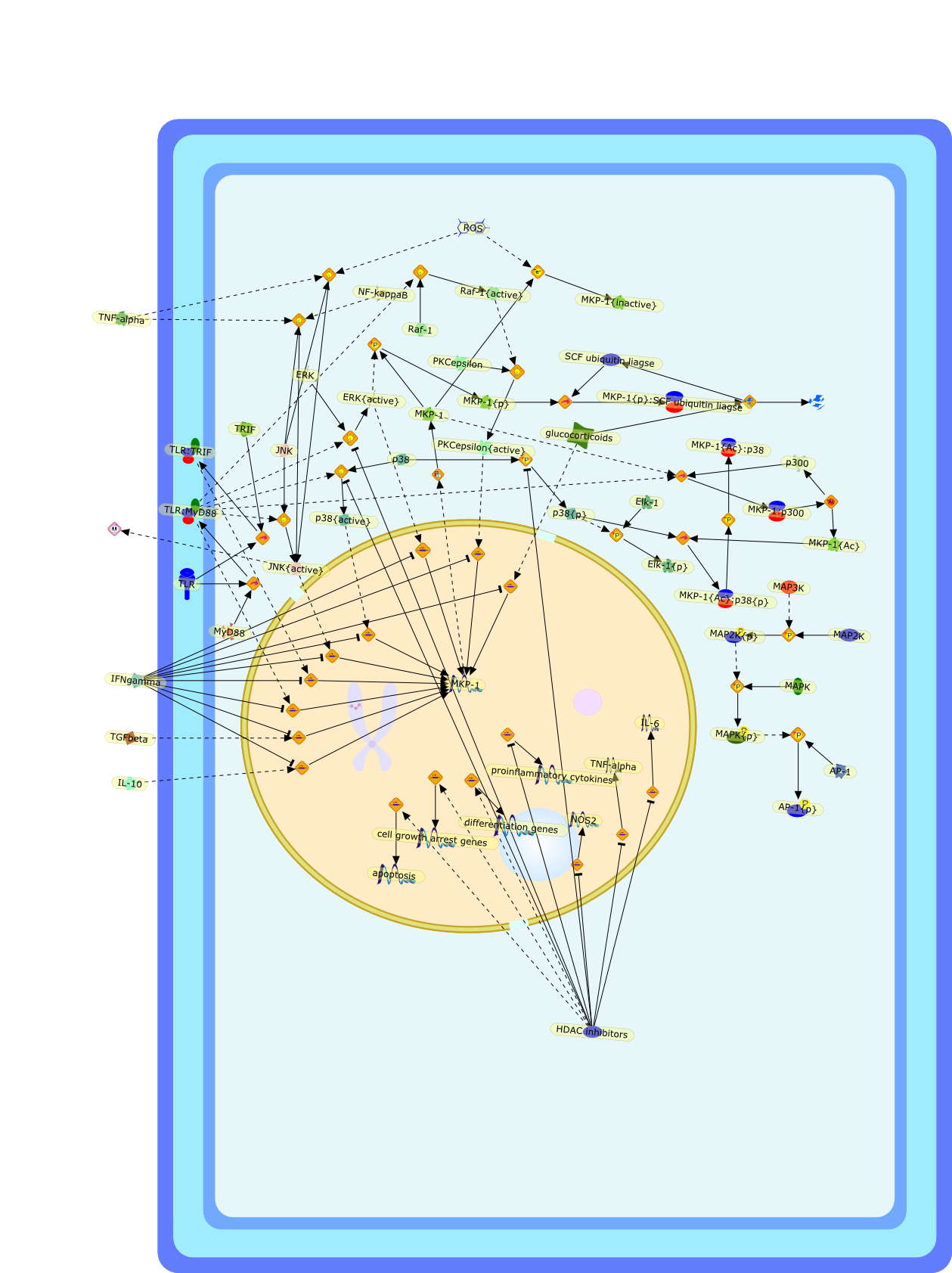{kind=link}
{kind=link}
AP-1,AP-1{p},ERK,ERK{active},Elk-1,Elk-1{p},HDAC inhibitors,IFNgamma,IL-10,IL-6,JNK,JNK{active},MAP2K,MAP2K{p},MAP3K,MAPK,MAPK{p},MKP-1,MKP-1:p300,MKP-1{Ac},MKP-1{Ac}:p38,MKP-1{Ac}:p38{p},MKP-1{inactive},MKP-1{p},MKP-1{p}:SCF ubiquitin liagse,MyD88,NF-kappaB,NOS2,PKCepsilon,PKCepsilon{active},ROS,Raf-1,Raf-1{active},SCF ubiquitin liagse,TGFbeta,TLR,TLR:MyD88,TLR:TRIF,TNF-alpha,TRIF,apoptosis,cell growth arrest genes,degradants,differentiation genes,glucocorticoids,p300,p38,p38{active},p38{p},proinflammatory cytokines